

Preparation of Dendrobium officinale Flower Anthocyanin and Extended Lifespan in Caenorhabditis elegans

Abstract
:1. Introduction
2. Results
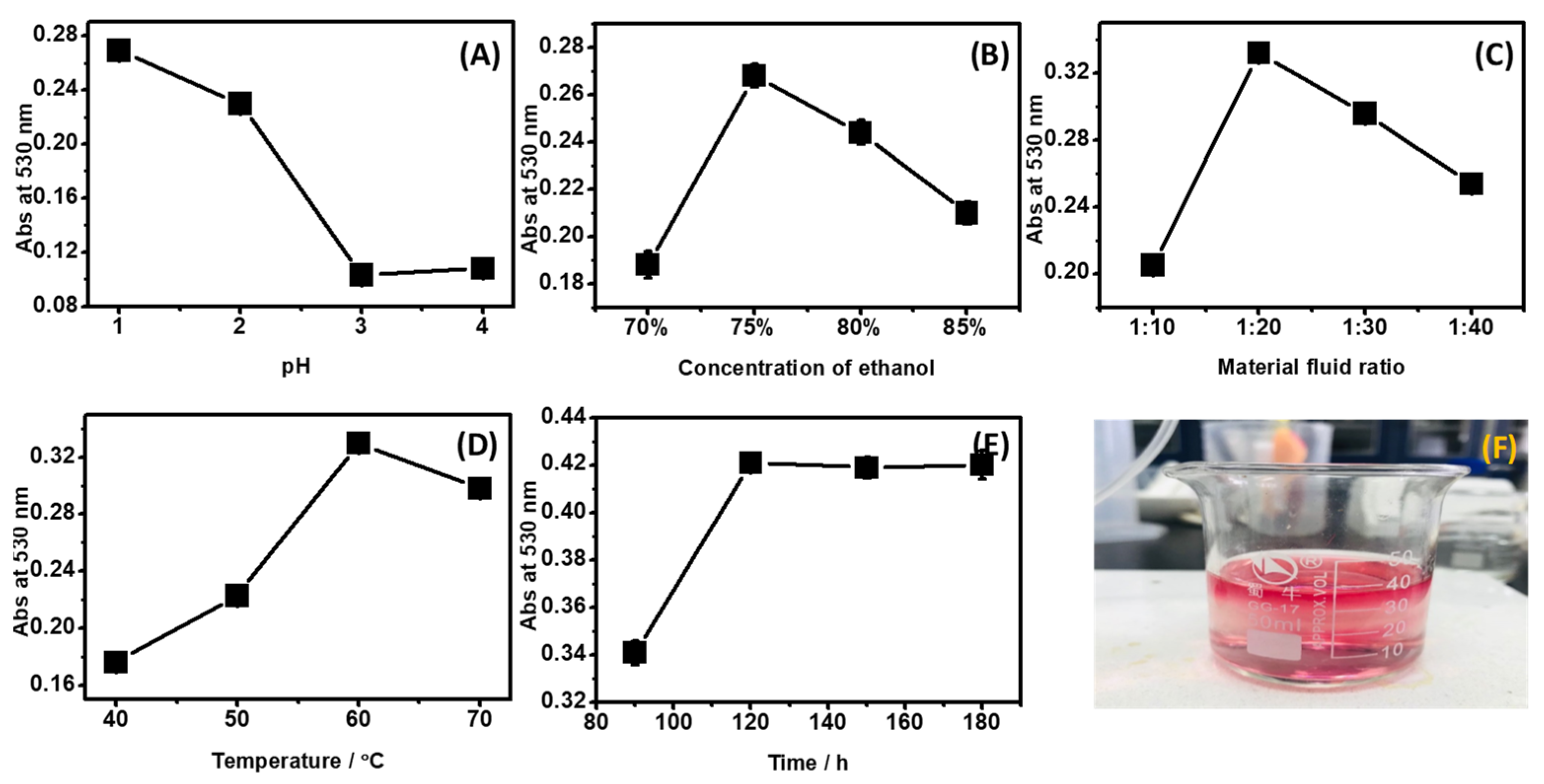
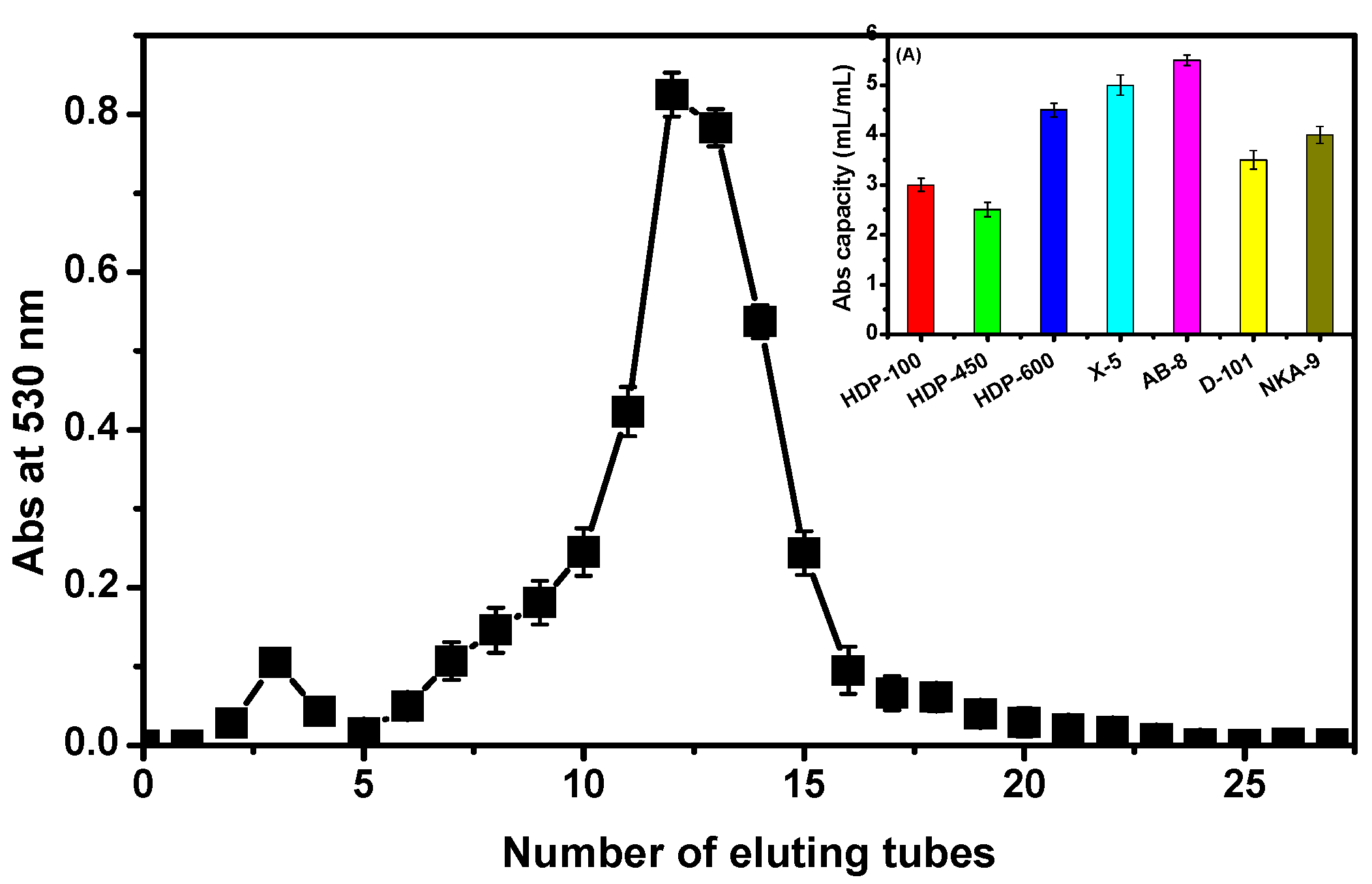
2.1. ACNs Extraction and Purification
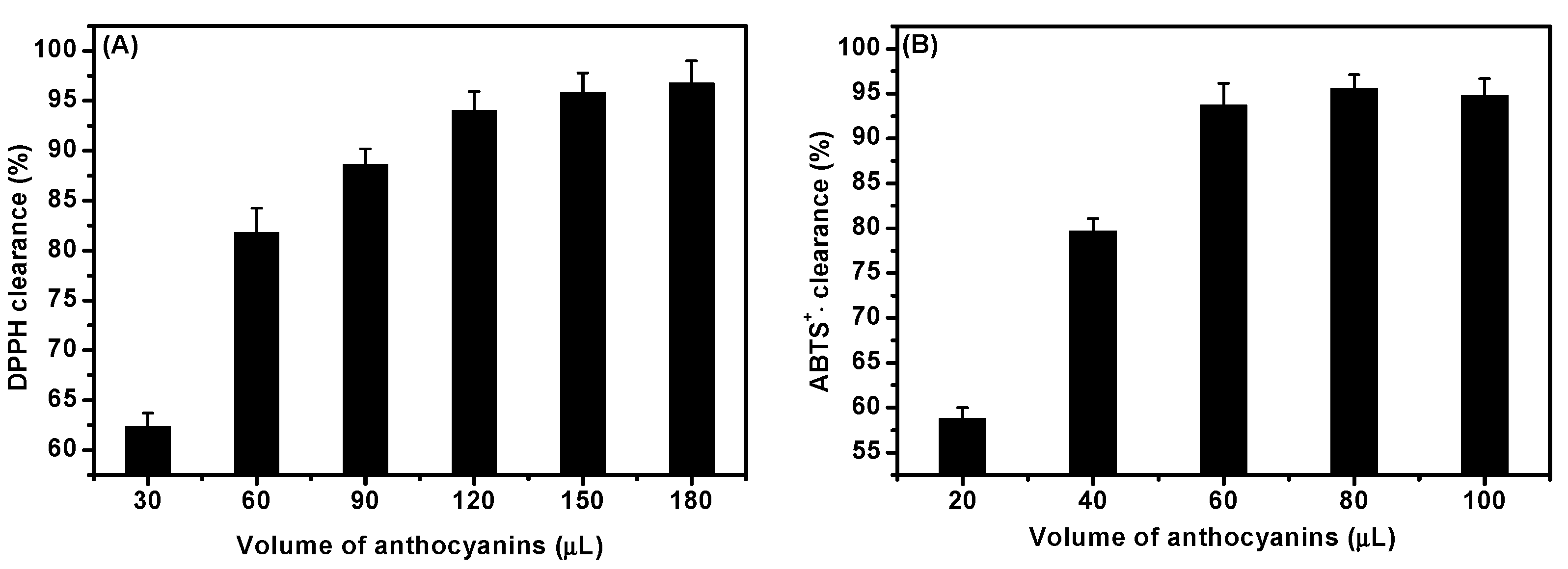
2.2. ACNs Antioxidant Activity In Vitro
2.3. Antioxidant Properties In Vivo
2.4. Effect of ACNs on Longevity and Spawning
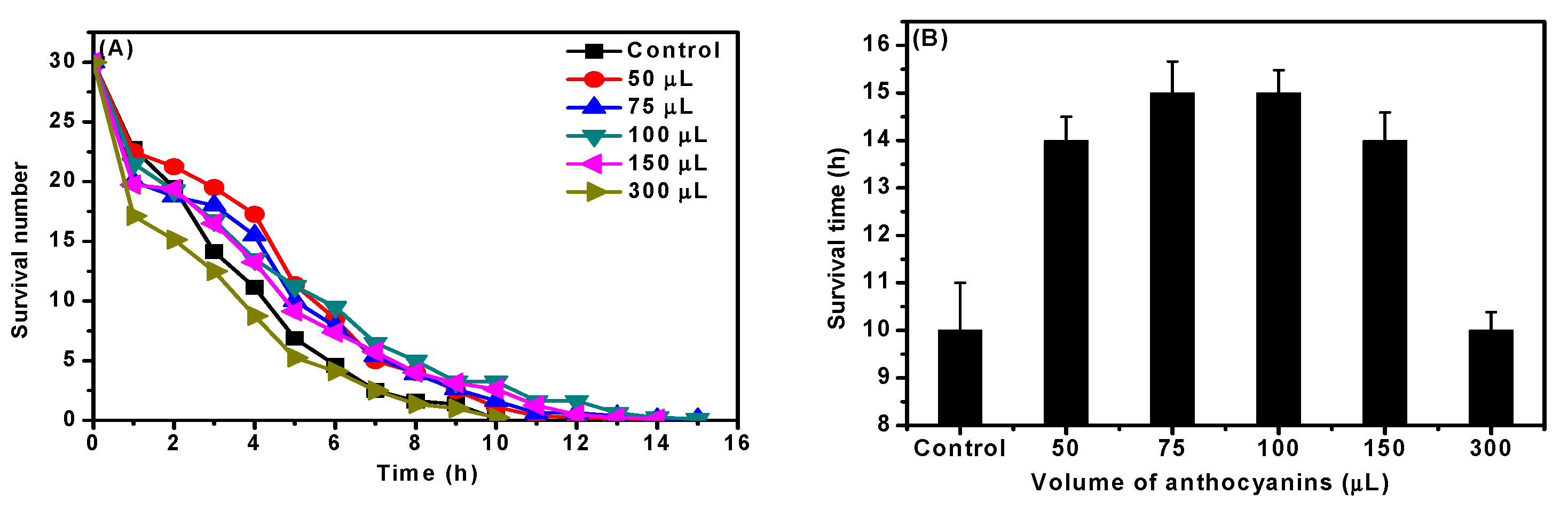
2.5. Effect of ACNs on Stress Resistance
2.5.1. Heat Stress Resistance
2.5.2. Oxidative Stress Resistance
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Extraction of the ACNs
4.3. Purification of the ACNs
4.4. In Vitro Antioxidation Assay
4.5. Effect of ACNs on Antioxidant Enzyme Activity in C. elegans
4.6. Lifespan Assay
4.7. Fecundity Assay
4.8. Assessment of Stress Resistance
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Song, B.B.; Wang, H.; Xie, W.; Zheng, B.S.; Li, T.; Liu, R.H. Combination of apple peel and blueberry extracts synergistically induced lifespan extension via DAF-16 in Caenorhabditis elegans. Food Funct. 2020, 11, 6170–6185. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, S.; Zhai, L.; Sun, L.W.; Zhao, D.Q.; Wang, Z.Y.; Li, X.Y. Ginsenoside extract from ginseng extends lifespan and health span in Caenorhabditis elegans. Food Funct. 2021, 12, 6793–6808. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Cargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Herrera, P.; Maieves, H.A.; Vega, E.N.; Perez-Rodriguez, M.L.; Fernandez-Ruiz, V.; Iriondo-DeHond, A.; Castillo, M.D.d.; Sanchez-Mata, M.C. Dwarf Kiwi (Actinidia arguta Miq.), a source of antioxidants for a healthy and sustainable diet. Molecules 2022, 27, 5495. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.F.; Liu, X.H.; Hang, S.Q.; Cao, C.X.; He, Y.C.; Sun, X.M.; Zheng, R.Q.; Yuan, J.F. Onion Vinegar Quality Evaluation and its Alleviate Oxidative Stress Mechanism in Caenorhabditis elegans Via SKN-1. Plant Food Hum. Nutr. 2022, 77, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Yu, Y.; Lin, H.; Liao, D.; Cui, X.; Wang, H. Lonicera japonica extends lifespan and health span in Caenorhabditis elegans. Free Radic. Biol. Med. 2018, 129, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Havermann, S.; Rohrig, R.; Chovolou, Y.; Humpf, H.U.; Wätjen, W. Molecular effects of Baicalein in Hct116 cells and Caenorhabditis elegans: Activation of the Nrf2 signaling pathway and prolongation of lifespan. J. Agric. Food Chem. 2013, 61, 2158–2164. [Google Scholar] [CrossRef] [PubMed]
- Havermann, S.; Humpf, H.U.; Wätjen, W. Baicalein modulates stress-resistance and life span in C. elegans via SKN-1. Fitoterapia 2016, 113, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Rubio, M.A.; Hernández-García, S.; Escribano, J.; Jiménez-Atiénzar, M.; Cabanes, J.; García-Carmona, F.; Gandía-Herrero, F. Betalain health-promoting effects after ingestion in Caenorhabditis elegans are mediated by DAF-16/FOXO and SKN-1/Nrf2 transcription factors. Food Chem. 2020, 330, 127228. [Google Scholar] [CrossRef]
- Sobeh, M.; ElHawary, E.; Peixoto, H.; Labib, R.M.; Handoussa, H.; Swilam, N.; El-Khatib, A.H.; Sharapov, F.; Mohamed, T.; Krstin, S.; et al. Identification of phenolic secondary metabolites from Schotia brachypetala Sond. (Fabaceae) and demonstration of their antioxidant activities in Caenorhabditis elegans. PeerJ 2016, 4, e2404. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.X.; Chen, Y.; Lin, Y.Z.; Wang, X.B.; Hu, L.Y.; Cao, Y.; Chen, Y.J. Antistress and anti-aging activities of Caenorhabditis elegans were enhanced by Momordica saponin extract. Eur. J. Nutr. 2021, 60, 1819–1832. [Google Scholar] [CrossRef]
- Martel, J.; Wu, C.Y.; Peng, H.H.; Ko, Y.F.; Yang, H.C.; Yong, J.D.; Ojcius, D.M. Plant and fungal products that extend lifespan in Caenorhabditis elegans. Microb. Cell 2020, 7, 255–269. [Google Scholar] [CrossRef]
- Yuan, Y.Y.; Kang, N.X.; Li, Q.X.; Zhang, Y.L.; Liu, Y.G.; Tan, P. Study of the effect of neutral polysaccharides from Rehmannia glutinosa on lifespan of Caenorhabditis elegans. Molecules 2019, 24, 4592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.H.; Chen, X.D.; Wang, D.; Wu, Z.; Chen, Y.; Wu, Q.X. Antioxidant and anti-aging effects of polysaccharide LDP-1 from wild Lactarius deliciosus on Caenorhabditis elegans. Food. Nutr. Res. 2022, 66, 8110. [Google Scholar] [CrossRef]
- Xu, D.L.; Lin, Q.L.; Wu, W.; Wu, Y.X.; Liang, Y. Revealing the antiaging effects of cereal- and food oil-derived active substances by a Caenorhabditis elegans model. Food Funct. 2021, 12, 3296. [Google Scholar] [CrossRef]
- Lv, X.F.; Zhou, X.H.; Wang, Y.; Wu, Y.N. Component analysis of Dendrobium phalaenopsis anthocyanin extract and its antioxidant activity and irritation in vitro. J. Trop. Subtrop. Bot. 2021, 29, 374–381. [Google Scholar]
- Chen, Z.Q.; Wang, C.; Pan, Y.X.; Gao, X.D.; Chen, H.X. Hypoglycemic and hypolipidemic effects of anthocyanins extract from black soybean seed coat in high fat diet and streptozotocin-induced diabetic mice. Food Funct. 2018, 9, 426–439. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.H.; Mo, J.L.; Ni, J.D.; Xu, Y.; Su, H.M.; Xie, J.H.; Chen, W. Structure-based design of human pancreatic amylase inhibitors from the natural anthocyanin database for type 2 diabetes. Food Funct. 2020, 11, 2910–2923. [Google Scholar] [CrossRef]
- Tan, J.Q.; Han, Y.M.; Han, B.; Qi, X.M.; Cai, X.; Ge, S.Q.; Xue, H.K. Extraction and purification of anthocyanins: A review. J. Agri. Food Res. 2022, 8, 100306. [Google Scholar] [CrossRef]
- Zhang, X.F.; Zhang, S.J.; Gao, B.B.; Qian, Z.; Liu, J.J.; Wu, S.H.; Si, J.P. Identification and quantitative analysis of phenolic glycosides with antioxidant activity in methanolic extraction of Dendrobium catenatum flowers and selection control herb-markers. Food Res. Int. 2019, 123, 732–745. [Google Scholar] [CrossRef]
- Wu, Y.L.; Huang, S.H.; He, C.M.; Qiu, B.; Liu, J.J.; Li, J.; Lin, Y.; Yu, S.L.; Wang, H.F.; Zhang, G.F. Dendrobium officinale flower extraction mitigates alcohol induced liver injury in mice: Role of antisteatosis, antioxidative, and antiinflammatory. Evid.-Based Complement. Altern. Med. 2020, 2020, 1421853. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.H.; Li, X.F.; Wang, M.N.; Zha, X.Q.; Yang, X.F.; Liu, Z.J.; Luo, Y.B.; Luo, J.P. Comparison of hypoglycemic and antioxidative effects of polysaccharides from four different Dendrobium species. Int. J. Biol. Macromol. 2014, 64, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.G.; Yang, L.L.; Zhang, Y.; Liu, X.C.; Wu, Z.J.; Gilbert, R.G.; Deng, B.; Wang, K.P. Dendrobium officinale polysaccharide ameliorates diabetic hepatic glucose metabolism via glucagon-mediated signaling pathways and modifying liver-glycogen structure. J. Ethnopharmacol. 2020, 248, 112308. [Google Scholar] [CrossRef]
- Wang, S.S.; Liu, J.M.; Sun, J.; Sun, Y.F.; Jia, N.; Liu, J.N.; Fan, B.; Wang, F.Z. Research progress on dendrobine of medicinal Dendrobium. Food Nutr. China 2020, 26, 9–15. [Google Scholar]
- Wang, D.H.; Fan, B.; Wang, Y.; Zhang, L.J.; Wang, F.Z. Optimum extraction, characterization, and antioxidant activities of polysaccharides from flowers of Dendrobium devonianum. Int. J. Anal. Chem. 2018, 2018, 3013497. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekhar, J.; Aduja, N.; Raghavarao, K.S.M.S. Purification of anthocyanins from jamun (Syzygium cumini L.) employing adsorption. Sep. Purif. Technol. 2014, 125, 170–178. [Google Scholar]
- Chandrasekhar, J.; Raghavarao, K.S.M.S. Process integration for purification and concentration of red cabbage (Brassica oleracea L.) anthocyanins. Sep. Purif. Technol. 2015, 141, 10–16. [Google Scholar]
- Hang, S.Q.; Sun, X.M.; He, Y.C.; Yuan, J.F. Microcapsules preparation of alcohol extracts from Nymphaea Hybrid and determination of its antioxidant capability in vitro and in vivo. Food Sci. Technol. 2022, 42, e91021. [Google Scholar] [CrossRef]
- Kudron, M.M.; Victorsen, A.; Gevirtzman, L.; Hillier, L.W.; Fisher, W.W.; Vafeados, D.; Kirkey, M.; Hammonds, A.S.; Gersch, J.; Ammouri, H.; et al. The ModERN resource: Genome-wide binding profiles for hundreds of Drosophila and Caenorhabditis elegans transcription factors. Genetics 2018, 208, 937–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, K.; Zhou, T.; Shao, J.F.; Ren, X.L.; Zhao, Z.Y.; Liu, D.H. Dynamic regulation of genetic pathways and targets during aging in Caenorhabditis elegans. Aging 2014, 6, 215–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Xia, C.; Zhong, X.; Lin, Q.; Li, X.; Zhang, Z.; Shi, W.; Xu, N.; Wu, Q.; Hu, Y.; et al. Food nutritional evaluation: Caenorhabditis elegans as a model organism. Shipin Kexue/Food Sci. 2019, 40, 268–276. [Google Scholar]
- Liang, C.Y.; Liang, Y.M.; Liu, H.Z.; Zhu, D.M.; Hou, S.Z.; Wu, Y.Y.; Huang, S.; Lai, X.P. Effect of Dendrobium officinale on D-galactose-induced aging mice. Chin. J. Integr. Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.H.; Wang, P.; Ma, Z.; Peng, L.; Wang, Z.M.; Chen, Z.L. Ultrasonic treatment of Dendrobium officinale polysaccharide enhances antioxidant and anti-inflammatory activity in a mouse D-galactose-induced aging model. Food Sci. Nutr. 2022, 10, 2620–2630. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Cai, H.; Li, W.D.; Cai, B.C. Ultrafiltration coupled with high-performance liquid chromatography and quadrupole time of flight mass spectrometry for screening lipase binders from different extracts of Dendrobium officinale. Anal. Bioanal. Chem. 2015, 407, 6081–6093. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.L.; Fang, P.; Shi, Q.Q.; Jie, S.U.; Bo, L.I.; Chen, S.H.; Lv, G.Y. Antihypertensive effect and mechanism of Dendrobium officinale flos on high blood pressure rats induced by high glucose and high fat compound alcohol. Zhongguo Zhongyao Zazhi/Chin. J. Chin. Mater. Med. 2018, 43, 147–153. [Google Scholar]
- Cacace, J.E.; Mazza, G. Extraction of anthocyanins and other phenolics from black currants with sulfured water. J. Agric. Food Chem. 2002, 50, 5939–5946. [Google Scholar] [CrossRef]
- Bridgers, E.N.; Chinn, M.S.; Truong, V.-D. Extraction of anthocyanins from industrial purple-fleshed sweet potatoes and enzymatic hydrolysis of residues for fermentable sugars. Ind. Crops. Prod. 2010, 32, 613–620. [Google Scholar] [CrossRef]
- Nayak, C.A.; Srinivas, P.; Rastogi, N.K. Characterization of anthocyanins from Garcinia indica Choisy. Food Chem. 2010, 118, 719–724. [Google Scholar] [CrossRef]
- Kirakosyan, A.; Seymour, E.M.; Llanes, D.E.U.; Kaufman, P.B.; Bolling, S.F. Chemical profile and antioxidant capacities of tart cherry products. Food Chem. 2009, 115, 20–25. [Google Scholar] [CrossRef]
- Lauren, F.F.; Ferreira, N.M.M.; Renata, F.R.; Roberson, P.; Eliseu, R.; Tatiana, E.; Vivian, C.B. Citric acid water-based solution for blueberry bagasse anthocyanins recovery: Optimization and comparisons with microwave-assisted extraction (MAE). LWT-Food Sci. Technol. 2020, 133, 110064. [Google Scholar]
- Li, X.J.; Liu, Y.M.; Wang, J.J.; Cao, S.R.; He, C.Y.; Song, Z.J. Effect of three extraction methods on the stability and antioxidant activities of anthocyanins from Dendrobium officinale. Nat. Prod. Res. Dev. 2018, 30, 26–35. [Google Scholar]
- Cao, S.Q.; Pan, S.Y.; Yao, X.L.; Fu, H.F. Isolation and purification of anthocyanins from blood organges by column chromatography. Agric. Sci. China 2010, 9, 207–215. [Google Scholar] [CrossRef]
- Grigoras, C.G.; Destandau, E.; Zubrzycki, S.; Elfakir, C. Sweet cherries anthocyanins: An environmental friendly extraction and purification method. Sep. Purif. Technol. 2012, 100, 51–58. [Google Scholar] [CrossRef]
- Li, X.C. Comparative study of 1,1-diphenyl-2-picryl-hydrazyl radical (DPPH·) scavenging capacity of the antioxidant xanthones family. ChemistrySelect 2018, 3, 13081–13086. [Google Scholar] [CrossRef]
- Ali, H.M.; Almagribi, W.; Al-Rashidi, M.N. Antiradical and reductant activities of anthocyanidins and anthocyanins, structure activity relationship and synthesis. Food Chem. 2016, 194, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C.; Ouyang, X.J.; Cai, R.X.; Chen, D.F. 3’,8’-Dimerization enhances the antioxidant capacity of flavonoids: Evidence from acacetin and isoginkgetin. Molecules 2019, 24, 2039. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Shao, S.; Ji, X.H.; Zhang, R.Y.; Zhang, S.W. Purification process of anthocyanin from opuntia fificus-indica. China Food Addit. 2022, 4, 108–115. [Google Scholar]
- Dehghan, E.; Zhang, Y.Q.; Saremi, B.; Yadavali, S.; Hakimi, A.; Dehghani, M.; Goodarzi, M.; Tu, X.Q.; Robertson, S.; Lin, R.; et al. Hydralazine induces stress resistance and extends C. elegans lifespan by activating the NRF2/SKN-1 signaling pathway. Nat. Commun. 2017, 8, 2223. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SOD Activity (U/mg Prot) | CAT Activity (U/mg Prot) | GSH-Px Activity (U/mg Prot) | |
|---|---|---|---|
| D. officinale ACNs | 1.3087 ± 0.4451 a | 0.0578 ± 0.0091 b | 0.5641 ± 0.0415 b |
| Control | 0.6129 ± 0.0828 | 0.0551 ± 0.0052 | 0.5385 ± 0.0381 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wang, J.; Zhang, L.; Zheng, Y.; Ma, G.; Sun, X.; Yuan, J. Preparation of Dendrobium officinale Flower Anthocyanin and Extended Lifespan in Caenorhabditis elegans. Molecules 2022, 27, 8608. https://doi.org/10.3390/molecules27238608
Li S, Wang J, Zhang L, Zheng Y, Ma G, Sun X, Yuan J. Preparation of Dendrobium officinale Flower Anthocyanin and Extended Lifespan in Caenorhabditis elegans. Molecules. 2022; 27(23):8608. https://doi.org/10.3390/molecules27238608
Chicago/Turabian StyleLi, Shuangxi, Jianfeng Wang, Liangliang Zhang, Yang Zheng, Guorong Ma, Xiaoming Sun, and Jianfeng Yuan. 2022. "Preparation of Dendrobium officinale Flower Anthocyanin and Extended Lifespan in Caenorhabditis elegans" Molecules 27, no. 23: 8608. https://doi.org/10.3390/molecules27238608
APA StyleLi, S., Wang, J., Zhang, L., Zheng, Y., Ma, G., Sun, X., & Yuan, J. (2022). Preparation of Dendrobium officinale Flower Anthocyanin and Extended Lifespan in Caenorhabditis elegans. Molecules, 27(23), 8608. https://doi.org/10.3390/molecules27238608