The Quality and Bacterial Community Changes in Freshwater Crawfish Stored at 4 °C in Vacuum Packaging


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Determination of the Total Volatile Base Nitrogen (TVB-N) and Total Aerobic Microorganisms
2.3. Texture Profile Analysis (TPA)
2.4. GC-MS Measurement of Volatile Flavor Compounds
2.5. DNA Extraction
2.6. 16S rDNA Amplification and SMRT Sequencing
2.7. Sequence Analyses
2.8. Statistical Analysis
3. Results and Discussion
3.1. Total Volatile Base Nitrogen (TVB-N) and Total Aerobic Microorganisms
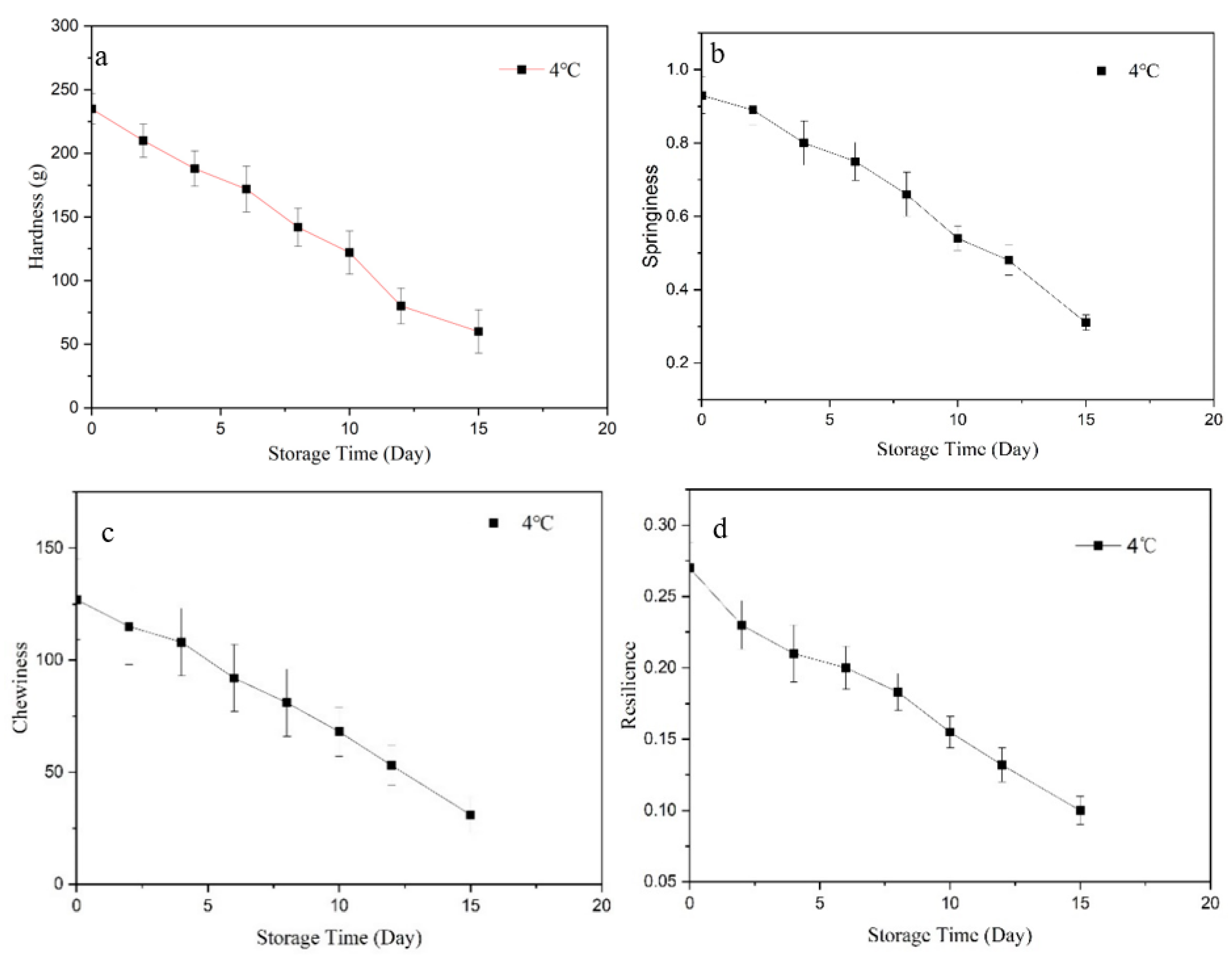
3.2. Texture Profile Analysis (TPA)
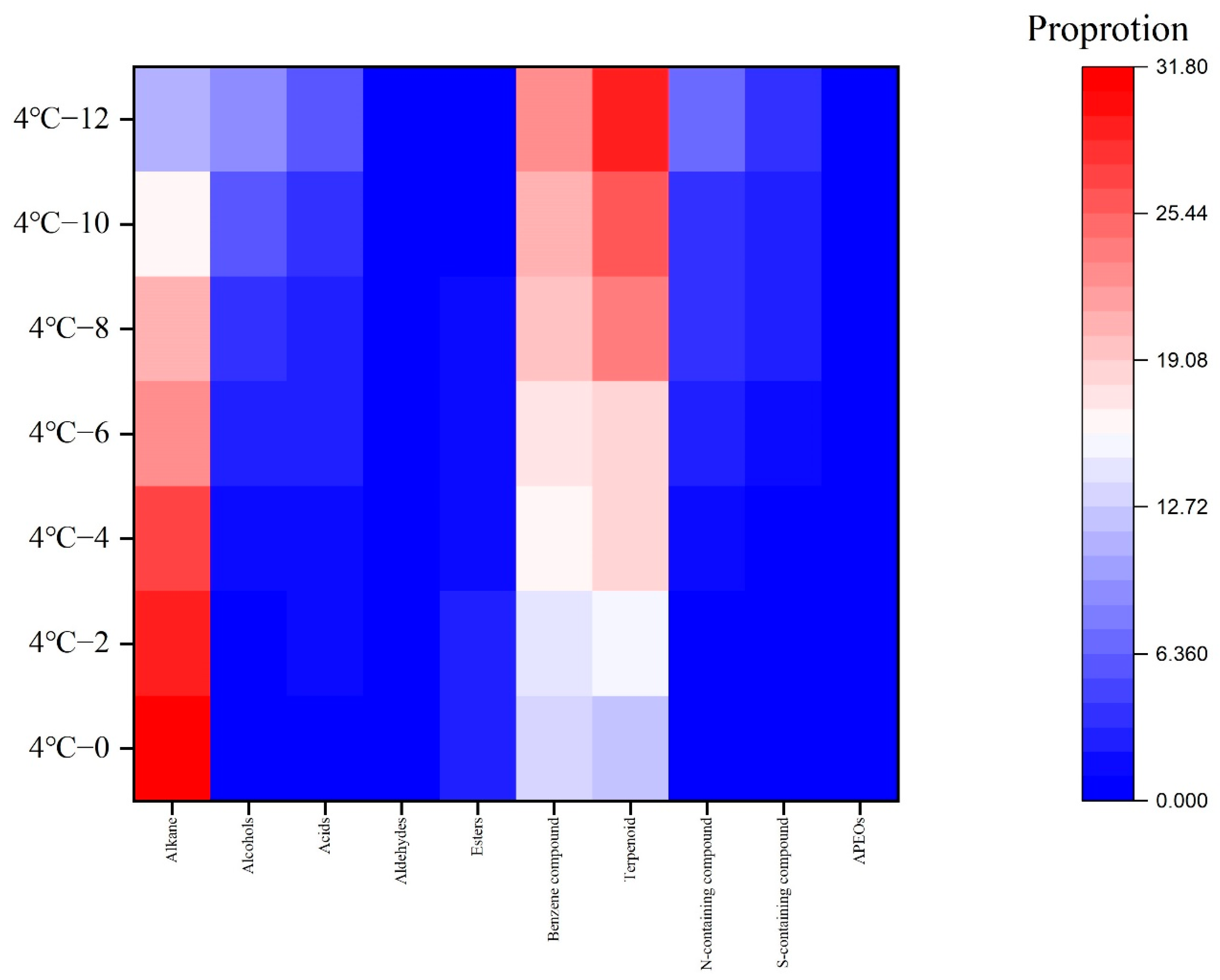
3.3. Volatile Flavor Compounds
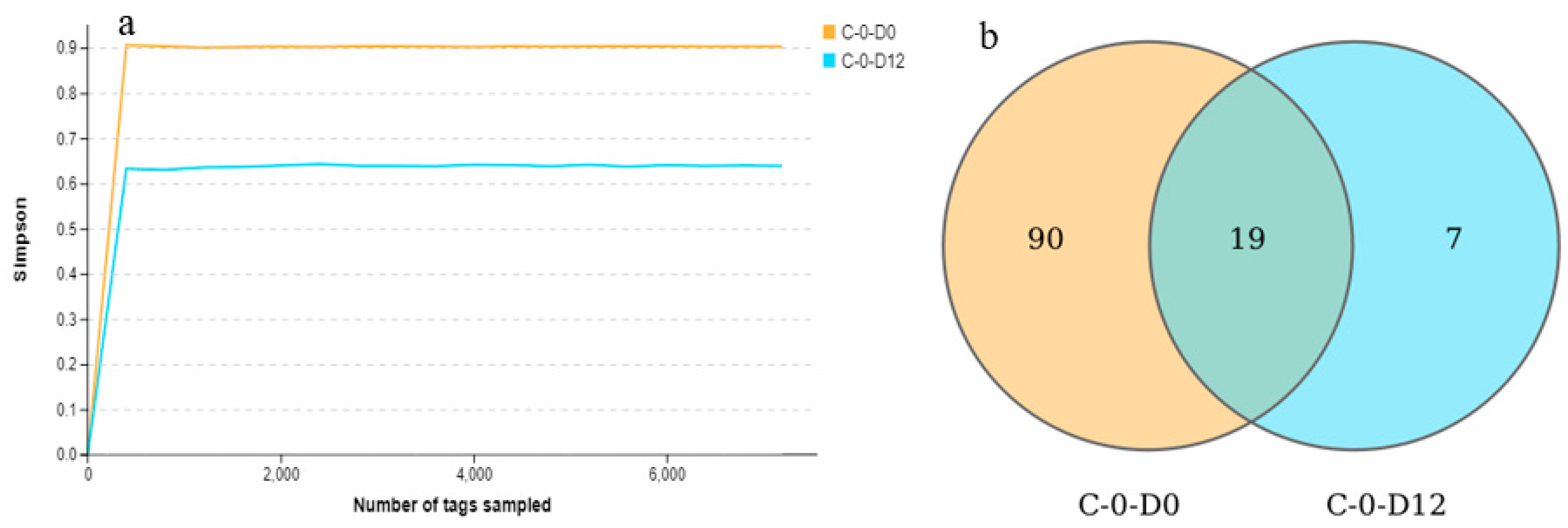
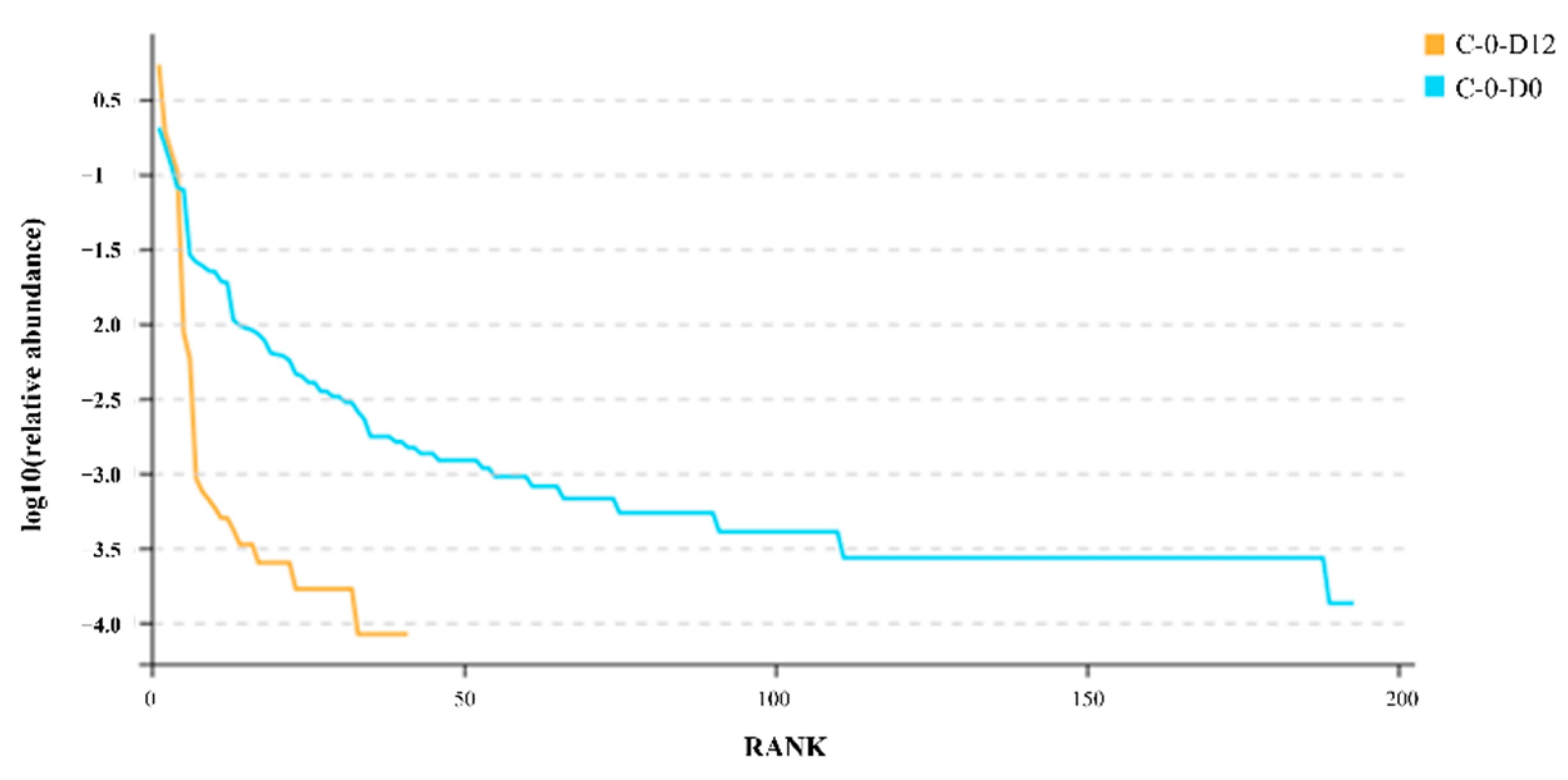
3.4. Dataset Features and Species Richness
3.5. Alpha Diversity of the Microbiota of the Crawfish
3.6. Correlation Analysis between Dominant Bacteria and Quality
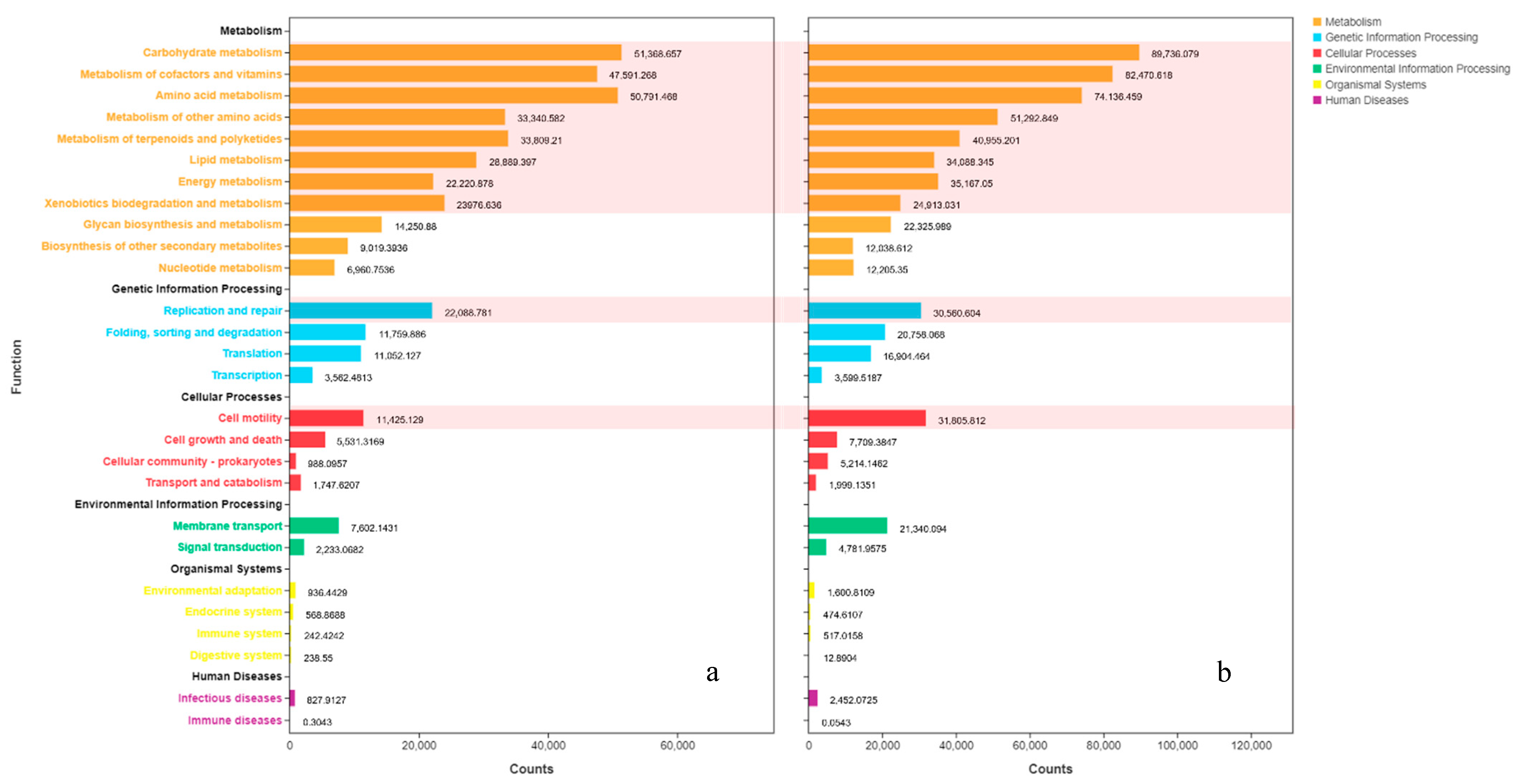
3.7. Function Prediction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Costa, M.J.; Duarte, G.; Segurado, P.; Branco, P. Major threats to European freshwater fish species. Sci. Total Environ. 2021, 797, 149105–149110. [Google Scholar] [CrossRef] [PubMed]
- Jaikumar, M. Food and Agriculture Organization of the United Nations for a World without Hunger Fisheries and Aquaculture Department Cultured Aquatic Species Information Programme Siganus spp.; Food and Agriculture Organization of the United Nations: Roma, Italy, 2019. [Google Scholar]
- Donsì, F.; Marchese, E.; Maresca, P.; Pataro, G.; Vu, K.D.; Salmieri, S.; Lacroix, M.; Ferrari, G. Green beans preservation by combination of a modified chitosan based-coating containing nanoemulsion of mandarin essential oil with high pressure or pulsed light processing. Postharvest. Biol. Technol. 2015, 106, 21–32. [Google Scholar] [CrossRef]
- Powrie, W.D. Low-Temperature Preservation of Foods and Living Matter; M. Dekker: New York, NY, USA, 1973. [Google Scholar]
- Yin, Y.M.; Xu, Y.X.; Zhang, C.M.; Li, X.P.; Li, Q.Y.; Xie, J.; Li, J.R. The progress on flavor deterioration mechanism of aquatic products during storage. Food Ferment. Ind. 2020, 46, 269–274. [Google Scholar]
- Kerry, J.F. Advances in meat, poultry and seafood packaging. Adv. Meat Poult. Seaf. Packag. 2012, 4, 661–681. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, H.; Liu, X.; Zhang, Y.; Lu, H.; Xu, Q.; Shi, C.; Luo, Y. Spoilage potential of three different bacteria isolated from spoiled grass carp (Ctenopharyngodon idellus) fillets during storage at 4 °C. LWT—Food Sci. Technol. 2016, 81, 10–17. [Google Scholar] [CrossRef]
- Levin, R.E. Detection and Incidence of Specific Species of Spoilage Bacteria on Fish. Appl. Microbiol. 1968, 16, 1734–1737. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Yu, D.; Tang, C.; Shen, J.; Xu, Y.; Xia, W.; Jiang, Q.; Yang, F. Physicochemical and Microbiological changes in Postmortem Crayfish (Procambarus clarkii) stored at 4 °C and 25 °C. Int. J. Food Sci. Technol. 2022, 13, 54–61. [Google Scholar] [CrossRef]
- Goldberg, I.; Rokem, J.S. Organic and Fatty Acid Production, Microbial. Encycl. Microbiol. 2009, 41, 421–442. [Google Scholar]
- Wang, H.; Li, Q. Quality and Bacterial phase changes of High-moisture Roast shrimp during storage at different temperatures. Food Mach. 2012, 21, 121–125. [Google Scholar]
- Yu, M.J.; Tan, H.; Shuang, H.E.; Hui, Y. Characterization of Specific Spoilage Bacteria and Volatile Flavor Compounds of Flavored Crayfish. Agric. Sci. Technol. 2021, 22, 42–50. [Google Scholar]
- Wang, R.R.; Lu, G.X.; Yu, Y.X. Effects of different quick-frozen treatments on the quality and safety of purple cabbage slices. Food Sci. Technol. 2017, 44, 156–168. [Google Scholar]
- Zhang, Y.; Magro, A.; Puolanne, E.; Zotte, A.D.; Ertbjerg, P. Myofibrillar protein characteristics of fast or slow frozen pork during subsequent storage at 3 °C. Meat Sci. 2021, 176, 108468. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.T.; Huang, J.Q.; Jiang, Y.Y.; Hu, Y.Q.; Liu, D.; Chen, J. Formation of hydrogels based on chitosan/alginate for the delivery of lysozyme and their and their antibacterial activity. Food Chem. 2017, 240, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.-Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, C.; Tsementzi, D.; Kyrpides, N.; Read, T.; Konstantinidis, K.T. Direct comparisons of illumina vs. Roche 454 sequencing technologies on the same microbial community DNA sample. PLoS ONE 2012, 7, e30087. [Google Scholar]
- He, Q.; Kwok, L.Y.; Xi, X.; Liu, Y.; Yang, Y.F. The meconium microbiota shares more features with the amniotic fluid microbiota than the maternal fecal and vaginal microbiota. Gut Microbes 2020, 12, 1794266. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcationof prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Yamamuro, A.; Kouzuma, A.; Abe, T.; Watanabe, K. Relative abundances of genus-level phylogenetic groups based on pyrosequenced 16S rRNA-gene amplicons showing structures of bacterial communities in the original sludge, anode biofilm, cathode biofilm, and electrolyte. PLoS One 2014, 22, 413–428. [Google Scholar]
- Bekhit, E.; Holman, B.; Giteru, S.G.; Hopkins, D.L. Total volatile basic nitrogen (TVB-N) and its role in meat spoilage: A review. Trends Food Sci. Technol. 2021, 109, 280–302. [Google Scholar] [CrossRef]
- Valizadeh, S.; Naseri, M.; Babaei, S.; Hosseini, S.M.H. Shelf life extension of fish patty using biopolymer-coated active paper sheets. Food Packag. Shelf Life 2020, 47, 133–140. [Google Scholar] [CrossRef]
- Kuswandi, B.; Larasati, T.S.; Abdullah, A.; Heng, Y.H. Real-Time Monitoring of Shrimp Spoilage Using On-Package Sticker Sensor Based on Natural Dye of Curcumin. Food Anal. Methods 2012, 5, 881–887. [Google Scholar] [CrossRef]
- Benjakul, S.; Visessanguan, W.; Thongkaew, C.; Tananka, M. Comparative study on physicochemical changes of muscle proteins from some tropical fish during frozen storage. Food Res. Int. 2003, 36, 787–795. [Google Scholar] [CrossRef]
- Benjakul, S.; Sutthipan, N. Muscle changes in hard and soft shell crabs during frozen storage. LWT—Food Sci. Technol. 2009, 42, 723–729. [Google Scholar] [CrossRef]
- Leduc, F.; Tournayre, P.; Kondjoyan, N.; Mercier, M.; Malle, P.; Kol, O.; Berdagué, O.; Duflos, G. Evolution of volatile odorous compounds during the storage of European seabass (Dicentrarchus labrax). Food Chem. 2012, 51, 641–650. [Google Scholar] [CrossRef]
- Xu, E.; Long, J.; Wu, Z.; Li, H.; Jiao, A. Characterization of Volatile Flavor Compounds in Chinese Rice Wine Fermented from Enzymatic Extruded Rice. J. Food Sci. 2015, 80, 111–120. [Google Scholar] [CrossRef]
- Hou, X.R.; Hong-Bo, M.I.; Mao, L.C. Influence of Thawing Methods on Physico-chemical Changes of Chinese Shrimp (Fenneropenaeus chinensis). Food Sci. 2014, 35, 243–247. [Google Scholar]
- Yang, F.; Wan, J.Q.; Zhang, N.; Xing, N. Research Progress on Corruption Mechanism and Control Methods of Aquatic Products. Sci. Technol. Food Ind. 2019, 41, 146–149. [Google Scholar]
- Majolini, D.; Trocino, A.; Tazzoli, M.; Xiccato, G. Evolution of European sea bass (Dicentrarchus labrax) freshness during storage. Ital. J. Anim. Sci. 2016, 8, 282–284. [Google Scholar] [CrossRef]
- Noseda, B.; Islamm, T. Microbiological spoilage of vacuum and modified atmosphere packaged Vietnamese Pangasius hypophthalmus fillets. Food Microbiol. 2012, 30, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Nychas, G.J.E.; Drosinos, E.H.; Board, R.G. Chemical changes in stored meat. Microbiol. Meat Poult. 1998, 6, 86–94. [Google Scholar]
- Gram, L.; Huss, H.H. Microbiological spoilage of fish and fish products. Int. J. Food Microbiol. 1996, 33, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Azzam-Sayuti, M.; Ina-Salwany, M.Y.; Zamri-Saad, M.; Annas, S.; Yusof, M.T.; Monir, M.S.; Mohamad, A.; Muhamad-Sofie, M.H.N.; Lee, J.Y.; Chin, Y.K.; et al. Comparative Pathogenicity of Aeromonas spp. in Cultured Red Hybrid Tilapia (Oreochromis niloticus × O. mossambicus). Biology 2021, 10, 1192. [Google Scholar] [CrossRef]
- Zhu, S.; Wu, H.; Zeng, M.; Liu, Z. The involvement of bacterial quorum sensing in the spoilage of refrigerated Litopenaeus vannamei. Int. J. Food Microbiol. 2015, 192, 26–33. [Google Scholar] [CrossRef]
- Yumoto, I. Oceanobacillus oncorhynchi sp. nov. a halotolerant obligate alkaliphile isolated from the skin of a rainbow trout (Oncorhynchus mykiss), and emended description of the genus Oceanobacillus. Int. J. Syst. Evol. Microbiol. 2005, 55, 1521. [Google Scholar] [CrossRef] [Green Version]
- Sangkaew, O.; Phaonakrop, N.; Roytrakul, S.; Yompakdee, C. Metaproteomic investigation of functional insight into special defined microbial starter on production of fermented rice with melanogenesis inhibition activity. PLoS ONE 2020, 15, e0241819. [Google Scholar] [CrossRef]
- Qian, Y.F.; Lin, T. Advances in microbial community interactions in aquatic products. Chin. J. Bioprocess Eng. 2020, 18, 150–157. [Google Scholar]
- Jia, S.; Liu, Y.; Zhuang, S.; Sun, S.; Li, Y.; Hong, H.; Lv, Y.; Luo, Y. Effect of ε-polylysine and ice storage on microbiota composition and quality of Pacific white shrimp (Litopenaeus vannamei) stored at 0 °C. Food Microbiol. 2019, 83, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lei, Y.; Tan, Y.; Zhang, J.; Hong, H.; Luo, Y. Efficacy of freeze-chilled storage combined with tea polyphenol for controlling melanosis, quality deterioration, and spoilage bacterial growth of Pacific white shrimp (Litopenaeus vannamei). Food Chem. 2022, 370, 130924. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xie, J.; Qian, Y. Determination of Spoilage Microbiota of Pacific White Shrimp During Ambient and Cold Storage Using Next-Generation Sequencing and Culture-Dependent Method. J. Food Sci. 2017, 82, 1178–1183. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, K.; Heyndrickx, M.; Herman, L.; Vlaemuclk, G. Molecular identification of the microbiota of peeled and unpeeled brown shrimp (Crangon crangon) during storage on ice and at 7.5 °C. Food Microbiol. 2013, 2, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Du, L.H.; Fan, X.R.; Liu, F.; Zhou, Q.; Yuan, J.A.; Ju, X.R. Changes of Dominant Spoilage Bacteria and Biogenic Amines of Taihu White Prawn (Exopalaemon modestus) during Ice Storage. J. Food Prot. 2017, 80, 2099–2104. [Google Scholar] [CrossRef]
- Lund, B.M.; Parker, T.C.; Gould, G.W. The Microbiological Safety and Quality of Food Toxigenic Fungi and Mycotoxin; Aspen Publishers: Boston, MA, USA, 2000. [Google Scholar]
- Cox, N.A.; Lovell, R.T. Identification and characterization of the microflora and spoilage bacteria in freshwater crayfish Procambarus clarkii (Girard). J. Food Sci. 2010, 38, 679–681. [Google Scholar] [CrossRef]
- Qian, Y.; Yang, S.; Xie, J.; Xiong, Q.; Li, Z.; Gao, S. Impact of the O2 Concentrations on Bacterial Communities and Quality of Modified Atmosphere Packaged Pacific White Shrimp (Litopenaeus Vannamei). J. Food Sci. 2013, 78, M1878–M1884. [Google Scholar] [CrossRef]
- Zhao, F.; Liu, H.; Zhang, Z.; Zhang, L.; Sun, X.H.; Xie, J.; Pan, Y.J.; Zhao, Y. Reducing bias in complex microbial community analysis in shrimp based on propidium monoazide combined with PCR-DGGE. Food Control 2016, 68, 139–144. [Google Scholar] [CrossRef]
- Macé, S.; Cardinal, M.; Jaffrès, E.; Cornet, J.; Lalanne, V.; Chevalier, F.; Serot, T.; Pilet, M.; Dousset, X.; Joffraud, J.J. Evaluation of the spoilage potential of bacteria isolated from spoiled cooked whole tropical shrimp (Penaeus vannamei) stored under modified atmosphere packaging. Food Microbiol. 2014, 40, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Wierda, R.L.; Fletcher, G.; Xu, L.; Jean-Pierre, D. Analysis of Volatile Compounds as Spoilage Indicators in Fresh King Salmon (Oncorhynchus tshawytscha) during Storage Using SPME−GC−MS. J. Agric. Food Chem. 2006, 54, 8480–8490. [Google Scholar] [CrossRef]
- Galli, A.; Franzetti, L.; Carelli, S. Microbiological quality and shelf-life of chilled cod fillets in vacuum-skin and modified atmosphere packaging. Packag. Technol. Sci. 2010, 6, 147–157. [Google Scholar] [CrossRef]
- Baltic, T.; Borovic, B.; Spiric, D. The influence of vacuum packaging on microbiological status and sensory properties of fresh beef. Tehnol. Mesa 2012, 53, 103–111. [Google Scholar] [CrossRef]
- Jia, S.; Li, Y.; Zhuang, S.; Sun, X.; Luo, Y. Biochemical changes induced by dominant bacteria in chill-stored silver carp (Hypophthalmichthys molitrix) and GC-IMS identification of volatile organic compounds. Food Microbiol. 2019, 84, 103248. [Google Scholar] [CrossRef] [PubMed]
- Ayala, M.D.; Abdel, I.; Santaella, M.; Martínez, C.; Periago, M.J.; Gil, F.; Blanco, A.; Albors, O.L. Muscle tissue structural changes and texture development in sea bream, Sparus aurata L. during post-mortem storage. Food Sci. Technol. 2010, 43, 465–475. [Google Scholar] [CrossRef]
- Zhao, Q.L.; Shen, Q.; Guo, R.; Wu, J.; Dai, Z. Characterization of Flavor Properties from Fish through Enzymatic Hydrolysis and Maillard Reaction. J. Aquat. Food Prod. Technol. 2013, 25, 182–195. [Google Scholar]
- Clark, K.A.; Mcelhinny, A.S.; Beckerle, M.C.; Gregorio, C.C. Striated muscle cytoarchitecture. Annu. Rev. Cell Dev. Biol. 2002, 18, 637–706. [Google Scholar] [CrossRef] [Green Version]
- Nirmal, N.P.; Benjakul, S. Inhibition of melanosis formation in Pacific white shrimp by the extract of lead (Leucaena leucocephala) seed. Food Chem. 2011, 128, 427–432. [Google Scholar] [CrossRef]
- Qiao, X.; Yang, L.; Gao, Q.; Shu, Y.; Li, S.; Zhao, J.; Jie, X.; Chang, H. Oxidation evaluation of free astaxanthin and astaxanthin esters in Pacific white shrimp during iced storage and frozen storage. J. Sci. Food Agric. 2018, 41, 245–256. [Google Scholar] [CrossRef]
- Annamalai, J.; Sasikala, R.; Debbarma, J.; Ravishankar, C.N.; Aliyamveetil, Z.A.; Ninan, G.; Ronda, V.; Velayudhanelayadom, L.K. Effect of delayed icing on the quality of white shrimp (litopenaeus vannamei) during chilled storage. J. Food Process. Preserv. 2015, 39, 2878–2885. [Google Scholar] [CrossRef]
- Bai, S.J.; Zheng, P.J.; Jiang, D.; Xin, Y.D.; Cheng, W.Z.; Liu, J.; Pei, X.Y.; Song, H.L. Comparative Study of the Gut Microbiota Among Four Different Marine Mammals in an Aquarium. Front. Microbiol. 2021, 11, 106–119. [Google Scholar] [CrossRef]
- Bartolo, I.; Birk, E.O. Some factors affecting Norway lobster (Nephrops norvegicus) cuticle polyphenol oxidase activity and blackspot development. Int. J. Food Sci. Technol. 2010, 33, 329–336. [Google Scholar] [CrossRef]
- Abramova, N.A.; Russell, J.; Botchan, M. Interaction between replication protein A and p53 is disrupted after UV damage in a DNA repair-dependentmanner. Proc. Natl. Acad. Sci. USA 1997, 94, 7186–7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeters, C.; Canck, E.D.; Cnockaert, M.; Brandt, E.D.; Vandamme, P. Comparative Genomics of Pandoraea, a Genus Enriched in Xenobiotic Biodegradation and Metabolism. Front. Microbiol. 2019, 10, 2556–2564. [Google Scholar] [CrossRef]
- Zhang, W. Involvement of protein degradation, calpain autolysis and protein nitrosylation in fresh meat quality during early postmortem refrigerated storage. Diss. Gradworks 2009, 12, 556–581. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | C-0-D0 (%) | C-0-D12 (%) |
|---|---|---|
| Aeromonas sobria | 0.03 | 55.09 |
| Paucibacter oligotrophus | 20.59 | 0 |
| Shewanella putrefaciens | 0.57 | 19.29 |
| Cloacibacterium rupense | 15.57 | 0 |
| Trichococcus pasteurii | 0.03 | 13.99 |
| Enterococcus aquimarinus | 0 | 10.39 |
| Flavobacterium sangjuense | 8.19 | 0 |
| Bacteroides nordii | 7.85 | 0.02 |
| Nitrospira moscoviensis | 5.19 | 0 |
| Brachymonas chironomi | 2.58 | 0 |
| Samples | OTUs | Shannon | Simpson |
|---|---|---|---|
| C-0-D0 | 193 | 4.46 | 0.91 |
| C-0-D12 | 41 | 1.88 | 0.64 |
| Level 1 | Level 2 | C-0-D0 (Abundance) | C-0-D12 (Abundance) |
|---|---|---|---|
| Metabolism | Carbohydrate metabolism | 51,368.66 | 89,736.08 |
| Metabolism | Metabolism of cofactors and Vitamins | 47,591.27 | 82,470.62 |
| Metabolism | Amino acid metabolism | 50,791.47 | 74,136.46 |
| Metabolism | Metabolism of other amino acids | 33,340.58 | 51,292.85 |
| Metabolism | Metabolism of terpenoids and polyketides | 33,809.21 | 40,955.21 |
| Metabolism | Lipid metabolism | 28,889.41 | 34,088.35 |
| Metabolism | Energy metabolism | 22,220.88 | 35,167.05 |
| Genetic information | Replication and repair | 22,088.78 | 30,560.60 |
| Metabolism | Xenobiotics biodegradation and metabolism | 23,976.63 | 24,913.03 |
| Cellular processes | Cell motility | 11,425.13 | 31,805.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, L.; Zhao, Y.; Ma, H.; Tian, X.; Bai, C.; Liao, T. The Quality and Bacterial Community Changes in Freshwater Crawfish Stored at 4 °C in Vacuum Packaging. Molecules 2022, 27, 8618. https://doi.org/10.3390/molecules27238618
Qiu L, Zhao Y, Ma H, Tian X, Bai C, Liao T. The Quality and Bacterial Community Changes in Freshwater Crawfish Stored at 4 °C in Vacuum Packaging. Molecules. 2022; 27(23):8618. https://doi.org/10.3390/molecules27238618
Chicago/Turabian StyleQiu, Liang, Yunchun Zhao, Hui Ma, Xiaofei Tian, Chan Bai, and Tao Liao. 2022. "The Quality and Bacterial Community Changes in Freshwater Crawfish Stored at 4 °C in Vacuum Packaging" Molecules 27, no. 23: 8618. https://doi.org/10.3390/molecules27238618
APA StyleQiu, L., Zhao, Y., Ma, H., Tian, X., Bai, C., & Liao, T. (2022). The Quality and Bacterial Community Changes in Freshwater Crawfish Stored at 4 °C in Vacuum Packaging. Molecules, 27(23), 8618. https://doi.org/10.3390/molecules27238618