
Chemical Composition and Antifungal Activity of Zanthoxylum armatum Fruit Essential Oil against Phytophthora capsici

 ,
,
Abstract
:1. Introduction
2. Results
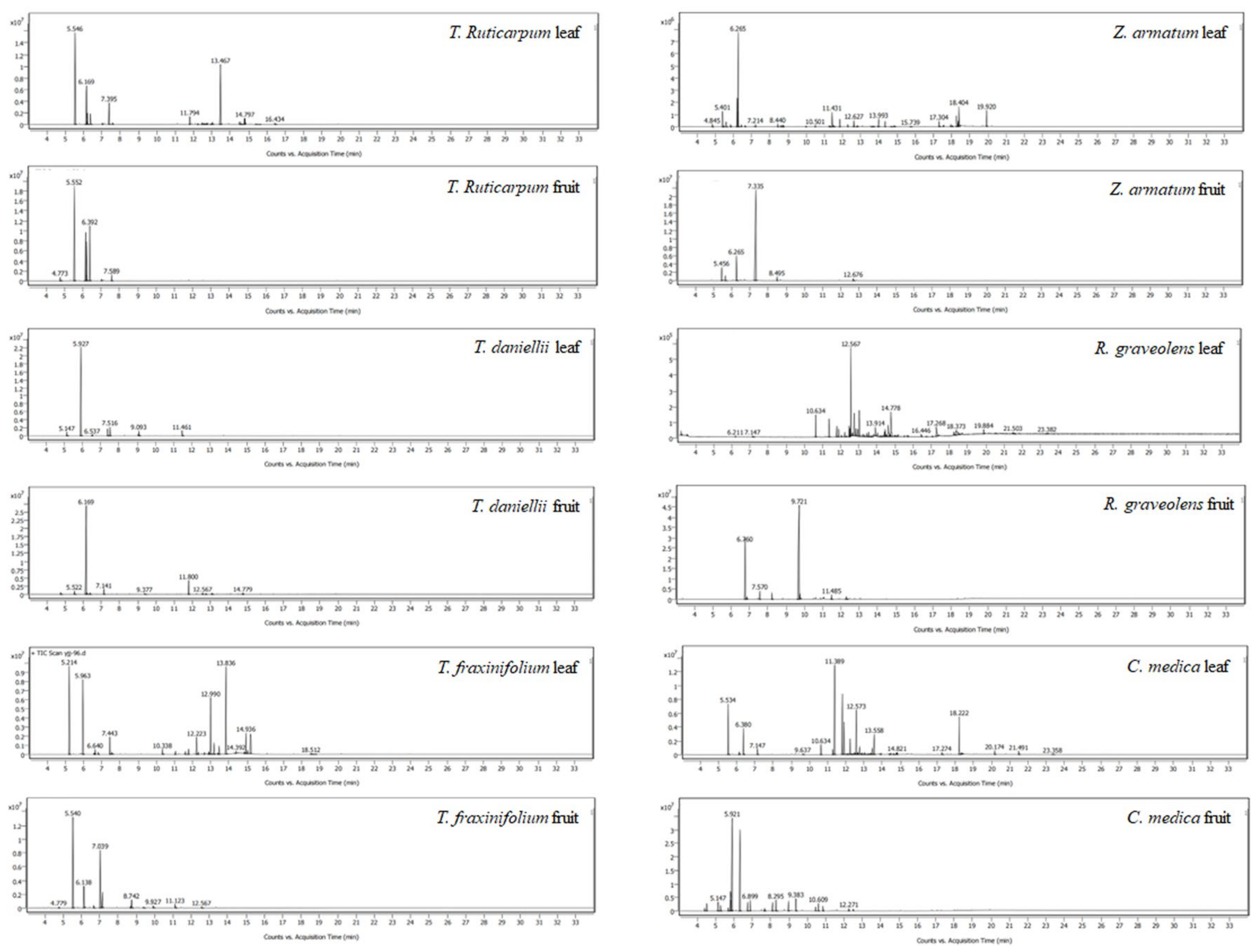
2.1. Chemical Composition of Twelve Plant EOs
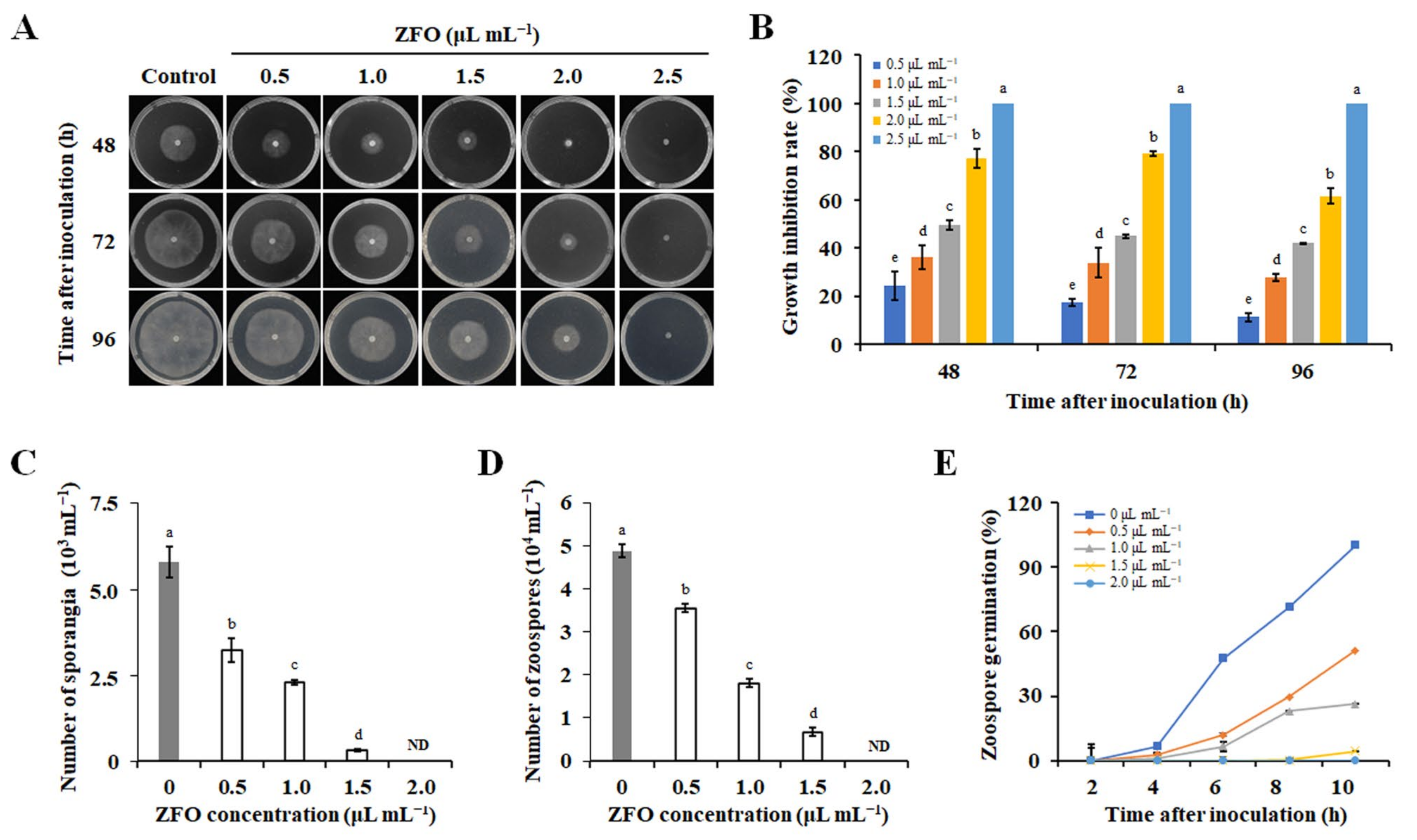
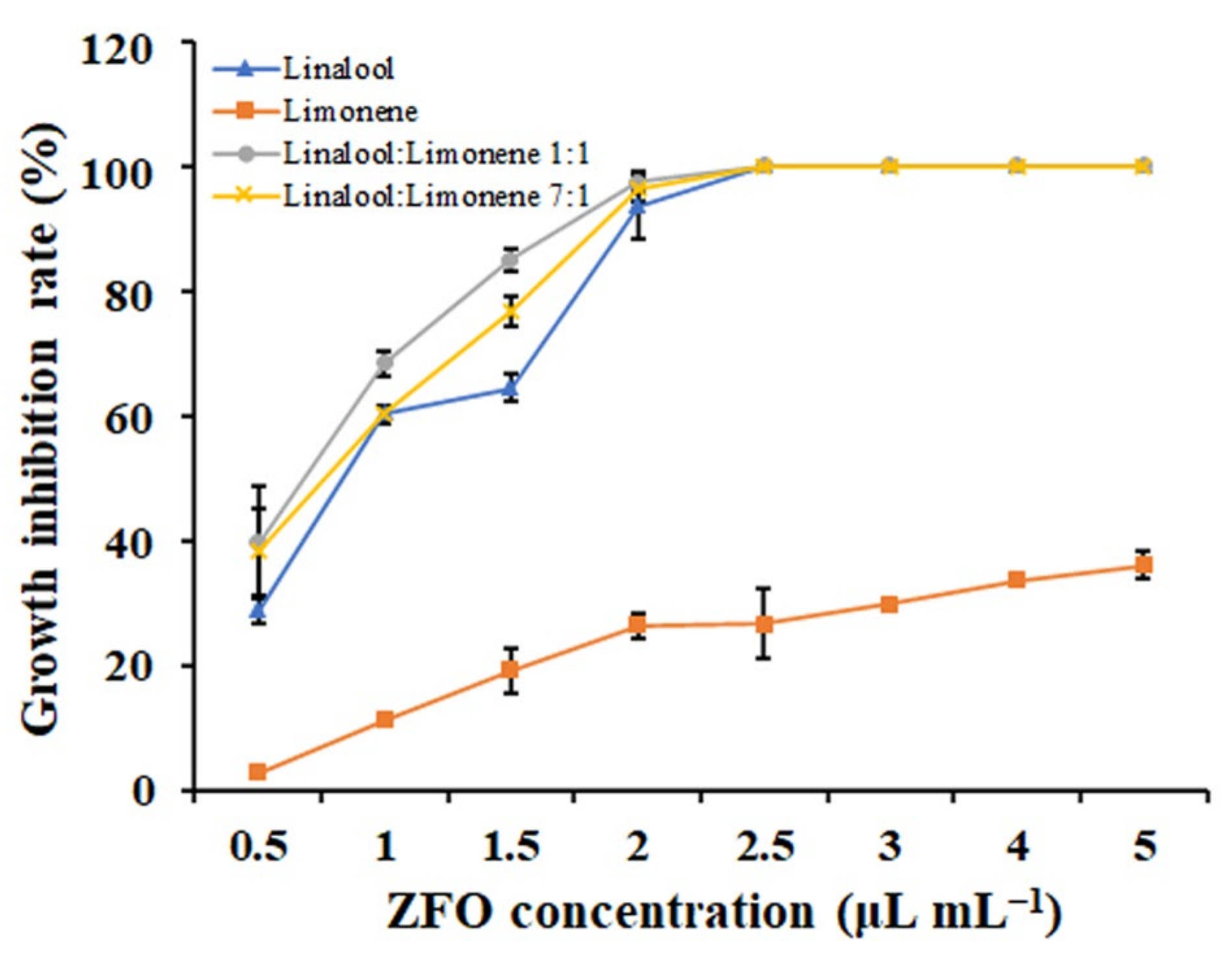
2.2. Antifungal Activities of the Twelve Plant EOs
2.3. ZFO Inhibited Secondary Infection of P. capsici
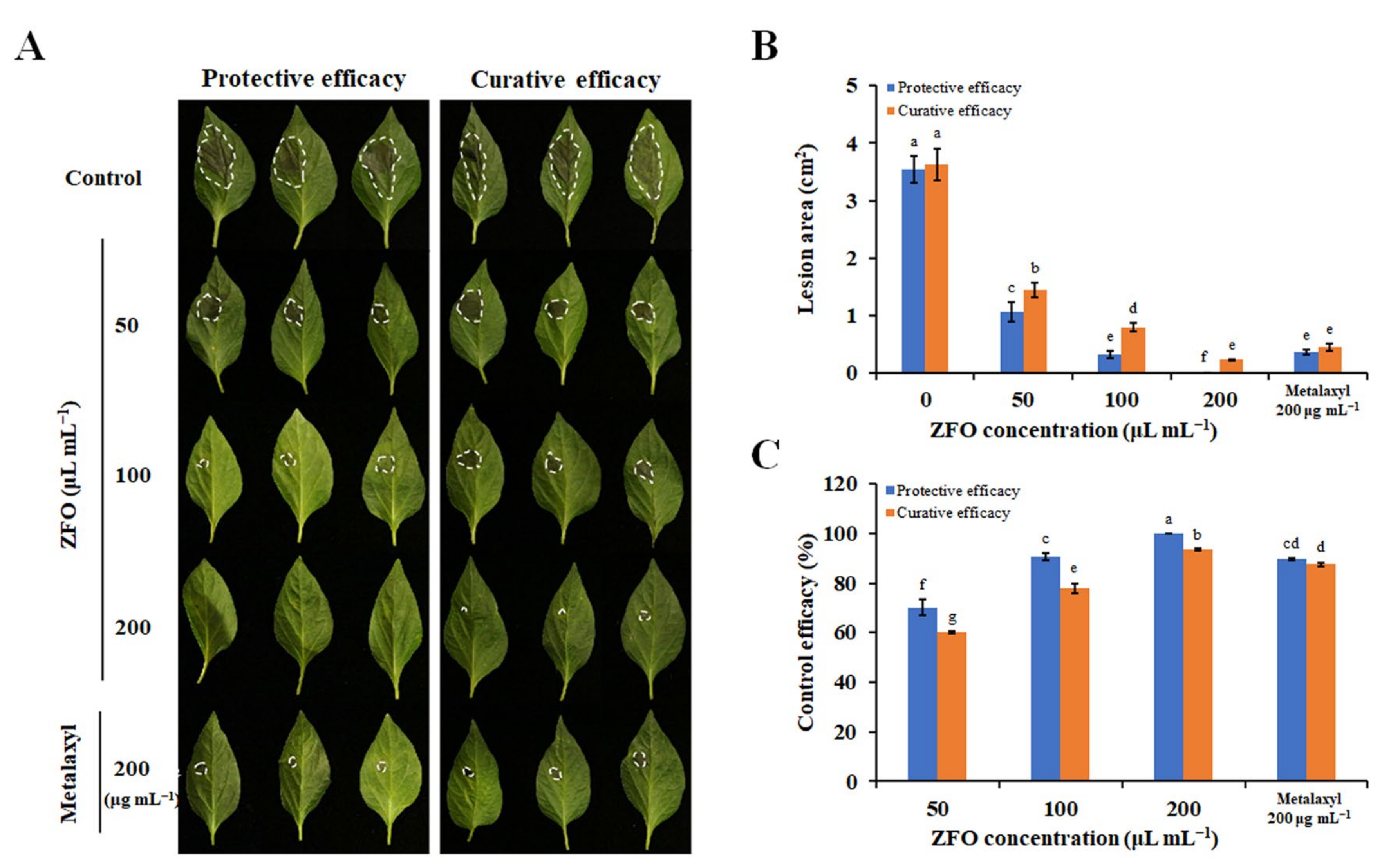
2.4. ZFO Inhibited P. capsici Infection in Pepper LEAVES and fruits
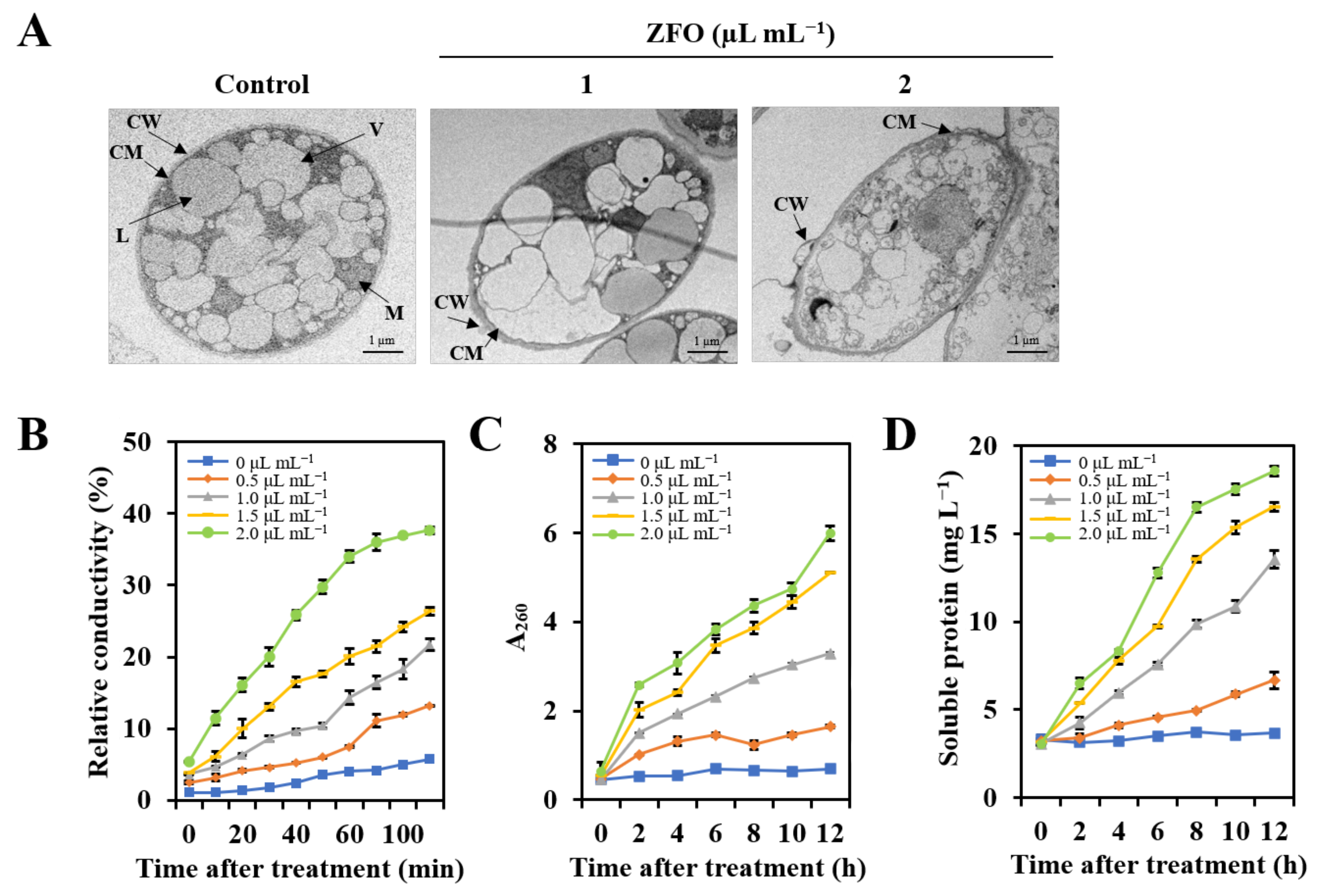
2.5. ZFO Destroyed the Ultrastructure of P. capsici
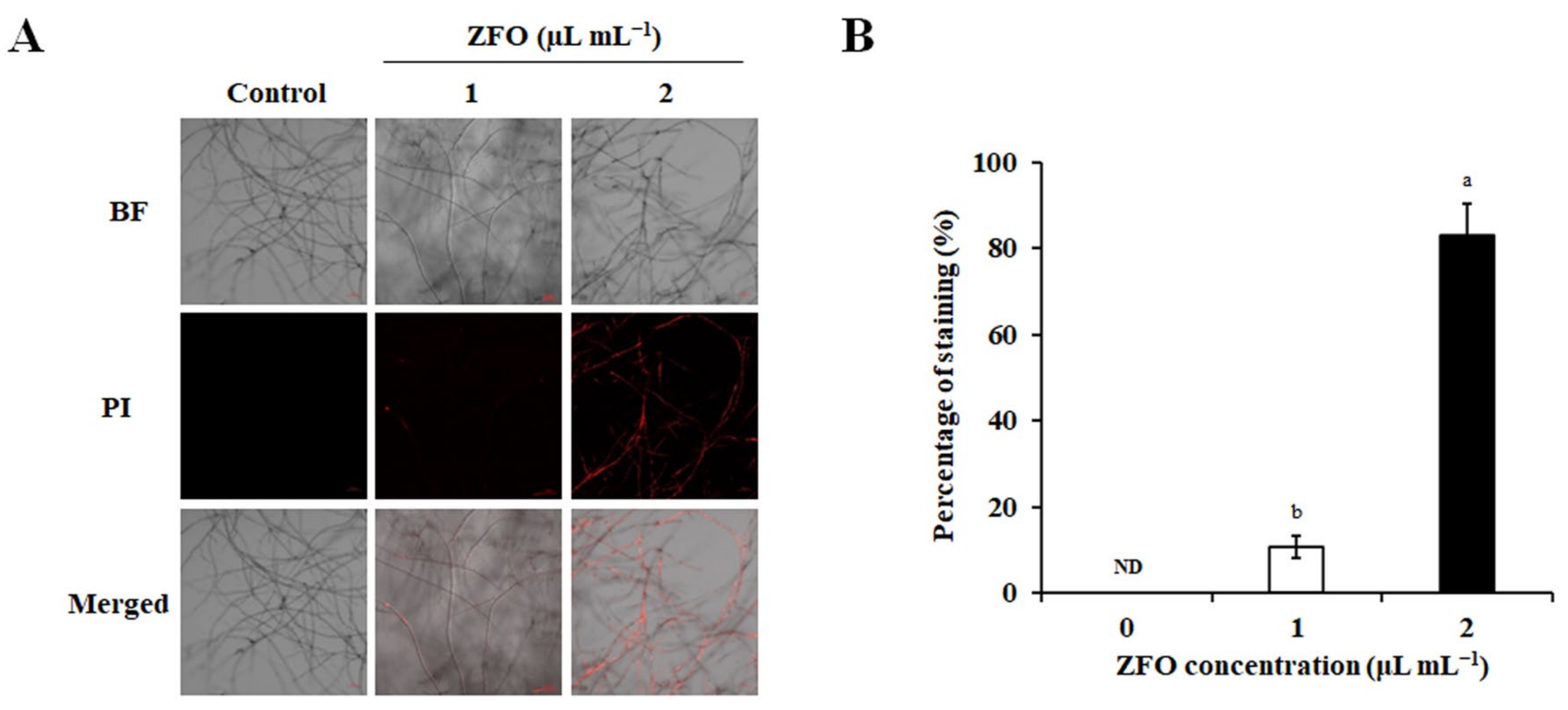
2.6. ZFO Damaged the Plasma Membrane of P. capsici
2.7. ZFO Induced Cellular Leakage of P. capsici
2.8. ZFO Increased the Defense-Related Enzyme Activities in Pepper Fruits
2.9. The Main Bioactive Components of ZFO
3. Discussion
4. Materials and Methods
4.1. Oomycete Pathogens and Chemicals
4.2. Plant Materials
4.3. EO Extraction
4.4. EO Identification
4.5. Effects of EOs on Mycelial Growth
4.6. Effects of ZFO on Secondary Infection of P. capsici
4.7. In Vivo Antifungal Bioassay of ZFO
4.8. Transmission Electron Microscopy (TEM) Analysis
4.9. Effects of ZFO on Membrane Damage of P. capsici
4.10. Effects of ZFO on Intracellular Leakage of P. capsici
4.11. Effects of ZFO on Defense-Related Enzyme Activities in Pepper Fruits
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lamour, K.H.; Stam, R.; Jupe, J.; Huitema, E. The oomycete broad-host-range pathogen Phytophthora capsici. Mol. Plant Pathol. 2012, 13, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Matson, M.E.H.; Small, I.M.; Fry, W.E.; Judelson, H.S. Metalaxyl resistance in Phytophthora infestans: Assessing role of rpa190 gene and diversity within clonal lineages. Phytopathology 2015, 105, 1594–1600. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Ai, G.; Shen, D.; Zou, F.; Wang, J.; Bai, T.; Chen, Y.; Li, S.; Zhang, M.; Jing, M.; et al. A Phytophthora capsici effector targets ACD11 binding partners that regulate ROS-mediated defense response in Arabidopsis. Mol. Plant 2019, 12, 565–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Ortuño, D.; Torés, J.A.; Chamorro, M.; Pérez-García, A.; de Vicente, A. Characterization of resistance to six chemical classes of site-specific fungicides registered for gray mold control on strawberry in Spain. Plant Dis. 2016, 100, 2234–2239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arif, T.; Bhosale, J.D.; Kumar, N.; Mandal, T.K.; Bendre, R.S.; Lavekar, G.S.; Dabur, R. Natural products-antifungal agents derived from plants. J. Asian Nat. Prod. Res. 2009, 11, 621–638. [Google Scholar] [CrossRef]
- Hu, L.; Wang, D.; Liu, L.; Chen, J.; Xue, Y.; Shi, Z. Ca2+ efflux is involved in cinnamaldehyde-induced growth inhibition of Phytophthora capsici. PLoS ONE 2013, 8, e76264. [Google Scholar] [CrossRef] [Green Version]
- Nikkhah, M.; Hashemi, M. Boosting antifungal effect of essential oils using combination approach as an efficient strategy to control postharvest spoilage and preserving the jujube fruit quality. Postharvest Biol. Technol. 2020, 164, 111159. [Google Scholar] [CrossRef]
- An, P.; Yang, X.; Yu, J.; Qi, J.; Ren, X.; Kong, Q. α-Terpineol and terpene-4-ol, the critical components of tea tree oil, exert antifungal activities in vitro and in vivo against Aspergillus niger in grapes by inducing morphous damage and metabolic changes of fungus. Food Control 2019, 98, 42–53. [Google Scholar] [CrossRef]
- Bi, Y.; Jiang, H.; Hausbeck, M.K.; Hao, J.J. Inhibitory effects of essential oils for controlling Phytophthora capsici. Plant Dis. 2012, 96, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, T.; Singh, S.; Nishad, I.; Kumar, A.; Tiwari, N.; Tandon, S.; Saikia, D.; Verma, R.S. Chemical composition and antimicrobial activity of the essential oil of senescent leaves of guava (Psidium guajavav L.). Nat. Prod. Res. 2021, 35, 1393–1397. [Google Scholar] [CrossRef]
- Akkari, H.; Ezzine, O.; Dhahri, S.; B’chir, F.; Rekik, M.; Hajaji, S.; Darghouth, M.A.; Jamâa, M.L.B.; Gharbi, M. Chemical composition, insecticidal and in vitro anthelmintic activities of Ruta chalepensis (Rutaceae) essential oil. Ind. Crop. Prod. 2015, 74, 745–751. [Google Scholar] [CrossRef]
- Phuyal, N.; Jha, P.K.; Raturi, P.P.; Rajbhandary, S. Zanthoxylum armatum DC.: Current knowledge, gaps and opportunities in Nepal. J. Ethnopharmacol. 2019, 229, 326–341. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Q.; Yang, W.; Sun, B.; Zhou, Y.; Zheng, Y.; Huang, M.; Yang, W. Characterization of the potent odorants in Zanthoxylum armatum DC Prodr. pericarp oil by application of gas chromatography-mass spectrometry-olfactometry and odor activity value. Food Chem. 2020, 319, 126564. [Google Scholar] [CrossRef] [PubMed]
- de Souza, E.L.; de Barros, J.C.; de Oliveira, C.E.V.; da Conceição, M.L. Influence of Origanum vulgare L. essential oil on enterotoxin production, membrane permeability and surface characteristics of Staphylococcus aureus. Int. J. Food Microbiol. 2010, 137, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, M.; Ren, G.; Hua, G.; Mi, J.; Jiang, D.; Liu, C. Antifungal Activity of essential oil from Zanthoxylum armatum DC. on Aspergillus flavus and Aflatoxins in stored platycladi semen. Front. Microbiol. 2021, 12, 633714. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Zhang, Z.; Li, B.; Xu, Y.; Tian, S. Effect of natamycin on Botrytis cinerea and Penicillium expansum-Postharvest pathogens of grape berries and jujube fruit. Postharvest Biol. Technol. 2019, 151, 134–141. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Z.; Huang, Q.; Zhang, D. Antifungal activity of several essential oils and major components against wood-rot fungi. Ind. Crop. Prod. 2017, 108, 278–285. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, Y.H.; Ye, M.; Wang, K.B.; Fan, L.M.; Su, F.W. Chemical composition and antifungal activity of essential oil from Origanum vulgare against Botrytis cinerea. Food Chem. 2021, 365, 130506. [Google Scholar] [CrossRef]
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Control 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Cao, J.Q.; Guo, S.S.; Wang, Y.; Pang, X.; Geng, Z.F.; Du, S.S. Toxicity and repellency of essential oil from Evodia lenticellata Huang fruits and its major monoterpenes against three stored-product insects. Ecotox Environ. Saf. 2018, 160, 342–348. [Google Scholar] [CrossRef]
- Li, X.; Ge, J.; Zheng, Q.; Zhang, J.; Sun, R.; Liu, R. Evodiamine and Rutaecarpine from Tetradium ruticarpum in the treatment of liver diseases. Phytomedicine 2020, 68, 153180. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhong, L.; Zeng, J.; Jiang, L.; Zhao, J. Study on the antibacterial activity of essential oils from seven species in the Rutaceae family. China Food Addit. 2020, 31, 37–41. [Google Scholar]
- Orlanda, J.F.F.; Nascimento, A.R. Chemical composition and antibacterial activity of Ruta graveolens L. (Rutaceae) volatile oils, from São Luís, Maranhão, Brazil. South Afr. J. Bot. 2015, 99, 103–106. [Google Scholar] [CrossRef]
- Alsaadi, J.H.H. Isolation, purification and identification of active chemical compound lup-20(29)-ene-3, 28-diol (Betulin) from Tetradium daniellii leaves and study the hypoglycemic effect on rabbits. Univ. Thi-Qar J. Sci. 2016, 6, 54–61. [Google Scholar]
- Liu, S.S.; Liu, Z.X.; Wei, H.; Yin, Y.Y.; Zhang, Q.W.; Yan, L.H.; Wang, Z.M.; Yang, L.X. Chemical compositions, yield variations and antimicrobial activities of essential oils from three species of Euodiae Fructus in China. Ind. Crop. Prod. 2019, 138, 111481. [Google Scholar] [CrossRef]
- Wu, K.; Jin, R.; Bao, X.; Yu, G.; Yi, F. Potential roles of essential oils from the flower, fruit and leaf of Citrus medica L. var. sarcodactylis in preventing spoilage of Chinese steamed bread. Food Biosci. 2021, 43, 101271. [Google Scholar] [CrossRef]
- Hosni, K.; Msaâda, K.; Taârit, M.B.; Ouchikh, O.; Kallel, M.; Marzouk, B. Essential oil composition of Hypericum perfoliatum L. and Hypericum tomentosum L. growing wild in Tunisia. Ind. Crop. Prod. 2008, 27, 308–314. [Google Scholar] [CrossRef]
- Abedini, S.; Sahebkar, A.; Hassanzadeh, M.K. Chemical composition of the essential oil of Thymus vulgaris L. grown in Iran. J. Essent. Oil Bear. Plants 2014, 17, 538–543. [Google Scholar] [CrossRef]
- Xu, D.; Deng, Y.; Han, T.; Jiang, L.; Xi, P.; Wang, Q.; Jiang, Z.; Gao, L. In vitro and in vivo effectiveness of phenolic compounds for the control of postharvest gray mold of Table grapes. Postharvest Biol. Technol. 2018, 139, 106–114. [Google Scholar] [CrossRef]
- Wang, B.; Li, P.; Xu, S.; Liu, L.; Xu, Y.; Feng, X.; Zhao, X.; Chen, Y. Inhibitory effects of the natural product esculetin on Phytophthora capsici and its possible mechanism. Plant Dis. 2021, 105, 1814–1822. [Google Scholar] [CrossRef]
- Shao, X.; Cheng, S.; Wang, H.; Yu, D.; Mungai, C. The possible mechanism of antifungal action of tea tree oil on Botrytis cinerea. J. Appl. Microbiol. 2013, 114, 1642–1649. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yuan, K.; Zhou, Q.; Lu, C.; Du, L.; Liu, F. Mechanism of antifungal activity of Perilla frutescens essential oil against Aspergillus flavus by transcriptomic analysis. Food Control 2021, 123, 107703. [Google Scholar] [CrossRef]
- Li, Z.; Wang, N.; Wei, Y.; Zou, X.; Jiang, S.; Xu, F.; Wang, H.; Shao, X. Terpinen-4-ol enhances disease resistance of postharvest strawberry fruit more effectively than tea tree oil by activating the phenylpropanoid metabolism pathway. J. Agric. Food Chem. 2020, 68, 6739–6747. [Google Scholar] [CrossRef]
- Li, Q.; Wang, J.; Bai, T.; Zhang, M.; Jia, Y.; Shen, D.; Zhang, M.; Dou, D. A Phytophthora capsici effector suppresses plant immunity via interaction with EDS1. Mol. Plant Pathol. 2020, 21, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Fridovich, I. Biological effects of the superoxide radical. Arch. Biochem. Biophys. 1986, 247, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Gu, Q.; Yun, Y.; Yin, Y.; Xu, J.R.; Shim, W.B.; Ma, Z. The TOR signaling pathway regulates vegetative development and virulence in Fusarium graminearum. New Phytol. 2014, 203, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Bi, Y.; Jiang, Q.; Mao, R.; Liu, Z.; Huang, Y.; Zhang, M.; Prusky, D.B. Induction of defense response against Alternaria rot in Zaosu pear fruit by exogenous L-lysine through regulating ROS metabolism and activating defense-related proteins. Postharvest Biol. Technol. 2021, 179, 111567. [Google Scholar] [CrossRef]
- Wang, B.; Li, P.; Yang, J.; Yong, X.; Yin, M.; Chen, Y.; Feng, X.; Wang, Q. Inhibition efficacy of Tetradium glabrifolium fruit essential oil against Phytophthora capsici and potential mechanism. Ind. Crop. Prod. 2022, 176, 114310. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Q.; Guo, Y.; Kang, L.; Yu, Y. Carbon monoxide enhances the resistance of jujube fruit against postharvest Alternaria rot. Postharvest Biol. Technol. 2020, 168, 111268. [Google Scholar] [CrossRef]
- Sato, K.; Krist, S.; Buchbauer, G. Antimicrobial effect of vapors of geraniol, (R)-(-)-linalool, terpineol, γ-terpinene and 1,8-cineole on airborne microbes using an airwasher. Flavour Fragr. J. 2007, 22, 435–437. [Google Scholar] [CrossRef]
- Marei, G.I.K.; Abdel Rasoul, M.A.; Abdelgaleil, S.A.M. Comparative antifungal activities and biochemical effects of monoterpenes on plant pathogenic fungi. Pestic. Biochem. Physiol. 2012, 103, 56–61. [Google Scholar] [CrossRef]
- Karimi, A.; Meiners, T. Antifungal activity of Zataria multiflora Boiss. essential oils and changes in volatile compound composition under abiotic stress conditions. Ind. Crop. Prod. 2021, 171, 113888. [Google Scholar] [CrossRef]
- Han, Y.; Sun, Z.C.; Chen, W.X. Antimicrobial susceptibility and antibacterial mechanism of limonene against Listeria monocytogenes. Molecules 2019, 25, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Lin, Z.X.; Xiang, W.L.; Huang, M.; Tang, J.; Lu, Y.; Zhao, Q.H.; Zhang, Q.; Rao, Y.; Liu, L. Antifungal activity and mechanism of D-limonene against foodborne opportunistic pathogen Candida tropicalis. LWT-Food Sci. Technol. 2022, 159, 113144. [Google Scholar] [CrossRef]
- Lu, X.H.; Zhu, S.S.; Bi, Y.; Liu, X.L.; Hao, J.J. Baseline sensitivity and resistance-risk assessment of Phytophthora capsici to iprovalicarb. Phytopathology 2010, 100, 1162–1168. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yang, Z.; Ying, G.; Yang, M.; Nian, Y.; Wei, F.; Kong, W. Antifungal evaluation of plant essential oils and their major components against toxigenic fungi. Ind. Crop. Prod. 2018, 120, 180–186. [Google Scholar] [CrossRef]
- Hosseini, S.; Amini, J.; Saba, M.K.; Karimi, K.; Pertot, I. Preharvest and postharvest application of garlic and rosemary essential oils for controlling anthracnose and quality assessment of strawberry fruit during cold storage. Front. Microbiol. 2020, 11, 1855. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Number | Retention Time (min) | Formula | Compound | Molecular Weight | Retention Indices a | Retention Indices b | Areas c (%) |
|---|---|---|---|---|---|---|---|
| 1 | 4.78 | C10H16 | 2-thujene | 136 | 968 | 978 | 0.1 |
| 2 | 4.9 | C10H16 | (-)-α-pinene | 136 | 942 | 936 | 0.25 |
| 3 | 5.3 | C8H16O | 2,4-dimethyl-cyclohexanol | 128 | 1032 | 1032 | 0.12 |
| 4 | 5.46 | C10H16 | sabinene | 136 | 967 | 946 | 5.79 |
| 5 | 5.53 | C10H16 | (-)-β-pinene | 136 | 943 | 0.27 | |
| 6 | 5.65 | C10H16 | β-myrcene | 136 | 983 | 984 | 2.04 |
| 7 | 5.82 | C8H16O | octanal | 128 | 982 | 1004 | 0.06 |
| 8 | 6.08 | C10H16 | α-terpinene | 136 | 1010 | 1010 | 0.33 |
| 9 | 6.27 | C10H16 | D-limonene | 136 | 1018 | 11.31 | |
| 10 | 6.5 | C10H16 | β-ocimene | 136 | 1037 | 1259 | 0.41 |
| 11 | 6.7 | C10H16 | γ-terpinene | 136 | 1050 | 1240 | 0.5 |
| 12 | 6.84 | C10H18O | 4-thujanol | 154 | 1093 | 1109 | 0.16 |
| 13 | 7.15 | C10H16 | α-terpinolene | 136 | 1079 | 1089 | 0.23 |
| 14 | 7.33 | C10H18O | linalool | 154 | 1086 | 1103 | 73.57 |
| 15 | 7.47 | C10H18O | isothujol | 154 | 1152 | 1170 | 0.1 |
| 16 | 7.61 | C10H16O | thujone | 152 | 1089 | 1105 | 0.07 |
| 17 | 7.67 | C10H18O | trans-p-2-menthen-1-ol | 154 | 1112 | 1108 | 0.09 |
| 18 | 7.92 | C10H18O | cis-p-menth-2-en-1-ol | 154 | 1111 | 1126 | 0.07 |
| 19 | 8.07 | C10H18O | citronellal | 154 | 1134 | 1153 | 0.08 |
| 20 | 8.29 | C10H20O | levomenthol | 156 | 1160 | 1172 | 0.12 |
| 21 | 8.49 | C10H18O | terpinen-4-ol | 154 | 1164 | 1167 | 1.39 |
| 22 | 8.68 | C10H18O | α-terpineol | 154 | 1175 | 1183 | 0.37 |
| 23 | 9.51 | C11H18O2 | linalyl formate | 182 | 1206 | 1215 | 0.08 |
| 24 | 9.61 | C10H16O | piperiton | 152 | 1233 | 1255 | 0.09 |
| 25 | 11.91 | C15H24 | caryophyllene | 204 | 1419 | 1423 | 0.26 |
| 26 | 12.34 | C15H24 | humulene | 204 | 1451 | 1436 | 0.07 |
| 27 | 12.68 | C15H24 | germacrene D | 204 | 1477 | 1482 | 0.92 |
| 28 | 12.87 | C15H24 | bicylogermacrene | 204 | 1492 | 1057 | 0.25 |
| 29 | 14.89 | C15H26O | α-cadinol | 222 | 1642 | 1635 | 0.06 |
| Total | 99.16 |
| Species | Organs | Growth Inhibition Rate (%) | ||||
|---|---|---|---|---|---|---|
| P. capsici | P. sojae | P. parasitica | Py. aphanidermatum | Py. ultimum | ||
| T. ruticarpum | Leaf | 39.02 ± 5.41 | 9.40 ± 2.55 | 20.90 ± 5.72 | 0 | 60.90 ± 5.02 |
| Fruit | 14.63 ± 2.17 | 5.20 ± 2.91 | 25.00 ± 3.28 | 0 | 43.50 ± 5.44 | |
| T. daniellii | Leaf | 27.91 ± 4.62 | 34.67 ± 4.64 | 0 | 22.00 ± 3.27 | 11.90 ± 2.76 |
| Fruit | 61.67 ± 1.06 | 35.52 ± 2.12 | 77.74 ± 2.03 | 0 | 22.17 ± 4.21 | |
| T. fraxinifolium | Leaf | 25.00 ± 2.58 | 31.50 ± 3.68 | 47.10 ± 5.70 | 5.00 ± 3.25 | 9.78 ± 2.83 |
| Fruit | 55.10 ± 4.71 | 25.80 ± 2.87 | 83.80 ± 6.45 | 15.00 ± 2.76 | 29.35 ± 3.16 | |
| Z. armatum | Leaf | 27.89 ± 3.84 | 51.68 ± 4.21 | 33.80 ± 3.98 | 25.85 ± 3.66 | 43.38 ± 3.75 |
| Fruit | 100 | 92.83 ± 5.24 | 95.75 ± 5.32 | 100 | 100 | |
| R. graveolens | Leaf | 0 | 0 | 0 | 12.37 ± 3.23 | 11.62 ± 2.12 |
| Fruit | 17.52 ± 3.71 | 10.32 ± 1.37 | 27.37 ± 1.94 | 3.71 ± 0.77 | 42.17 ± 4.60 | |
| C. medica | Leaf | 39.02 ± 4.38 | 9.40 ± 2.10 | 80.90 ± 4.35 | 0 | 60.90 ± 5.77 |
| Fruit | 42.19 ± 3.63 | 23.71 ± 2.76 | 54.31 ± 2.96 | 9.84 ± 0.31 | 39.25 ± 1.03 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Wang, Q.; Li, L.; Li, P.; Yin, M.; Xu, S.; Chen, Y.; Feng, X.; Wang, B. Chemical Composition and Antifungal Activity of Zanthoxylum armatum Fruit Essential Oil against Phytophthora capsici. Molecules 2022, 27, 8636. https://doi.org/10.3390/molecules27238636
Yang J, Wang Q, Li L, Li P, Yin M, Xu S, Chen Y, Feng X, Wang B. Chemical Composition and Antifungal Activity of Zanthoxylum armatum Fruit Essential Oil against Phytophthora capsici. Molecules. 2022; 27(23):8636. https://doi.org/10.3390/molecules27238636
Chicago/Turabian StyleYang, Jingjing, Qizhi Wang, Linwei Li, Pirui Li, Min Yin, Shu Xu, Yu Chen, Xu Feng, and Bi Wang. 2022. "Chemical Composition and Antifungal Activity of Zanthoxylum armatum Fruit Essential Oil against Phytophthora capsici" Molecules 27, no. 23: 8636. https://doi.org/10.3390/molecules27238636
APA StyleYang, J., Wang, Q., Li, L., Li, P., Yin, M., Xu, S., Chen, Y., Feng, X., & Wang, B. (2022). Chemical Composition and Antifungal Activity of Zanthoxylum armatum Fruit Essential Oil against Phytophthora capsici. Molecules, 27(23), 8636. https://doi.org/10.3390/molecules27238636