


UPLC-ESI-QTOF-MS Profiling of Phenolic Compounds from Eriocephalus africanus: In Vitro Antioxidant, Antidiabetic, and Anti-Inflammatory Potentials

Abstract
:1. Introduction
2. Results and Discussion
2.1. Phytochemical Compounds and Antioxidant Activity
2.2. Phytochemical Analysis of the Methanol and Chloroform Extracts of E. africanus
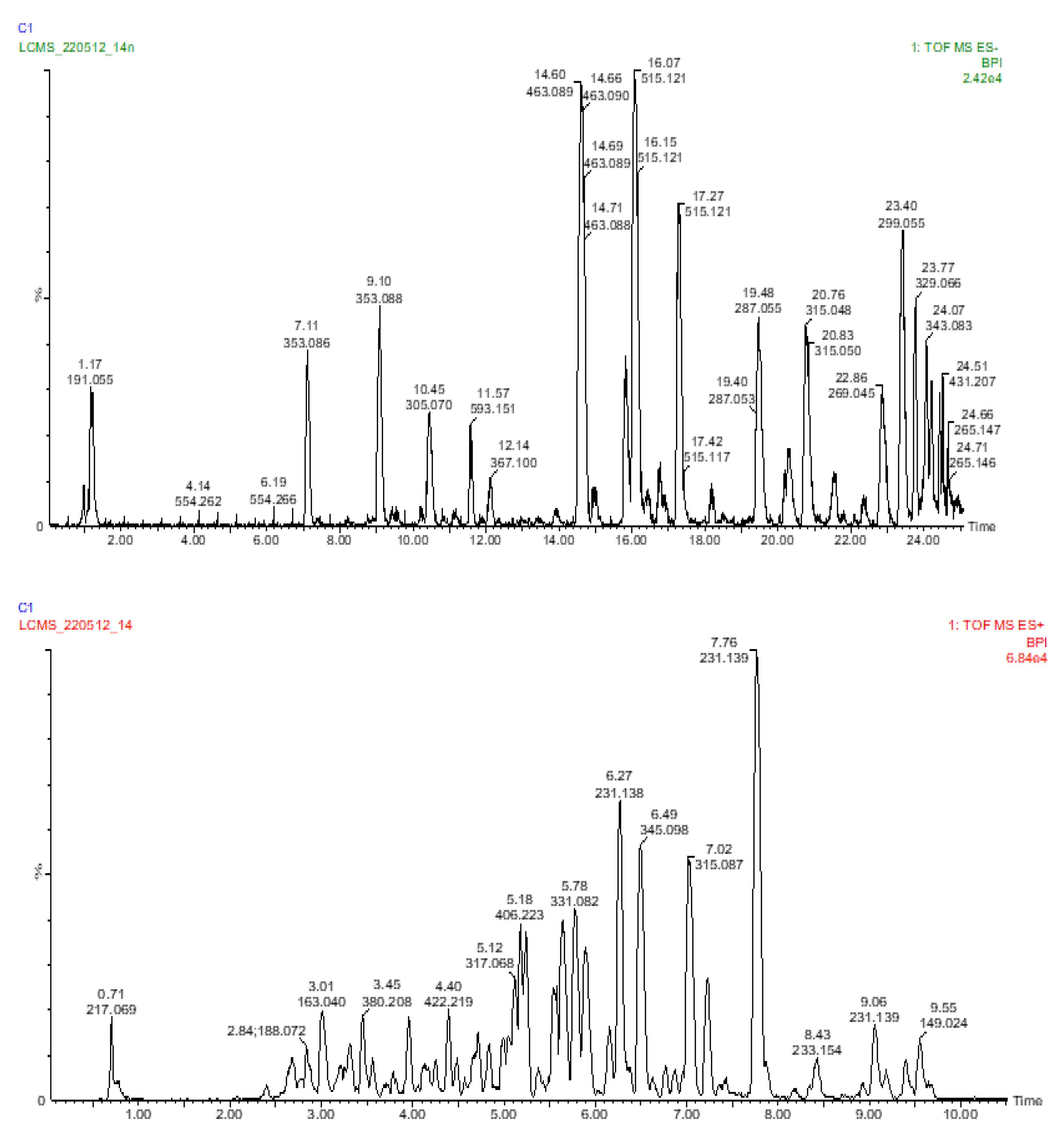
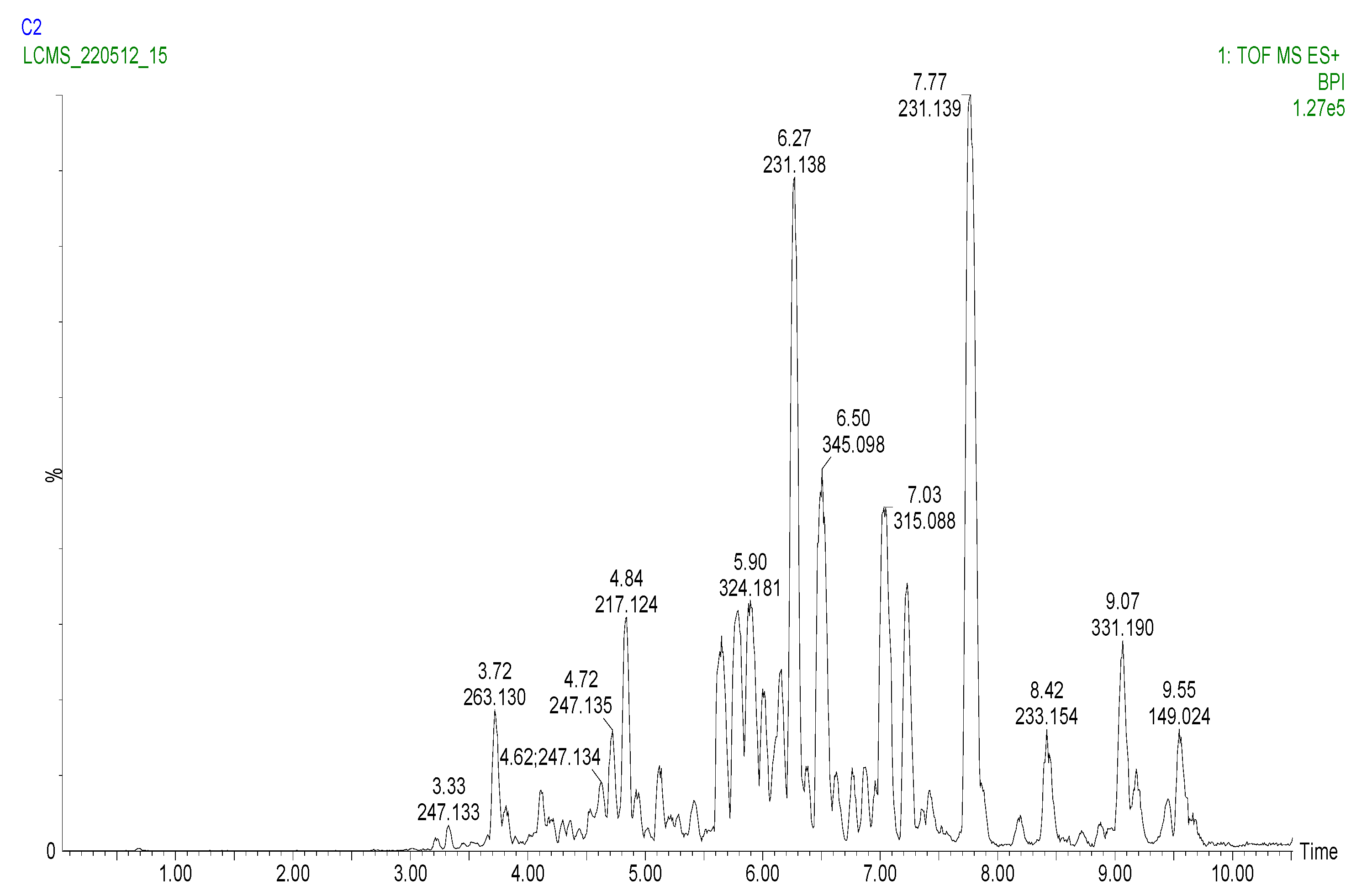
2.3. Analysis of the Phenolic Compounds and Other Metabolites
2.3.1. Analysis of the Carboxylic Acids
2.3.2. Analysis of the Hydroxycinnamic Acids
2.3.3. Analysis of the Flavanones, Flavones, Flavonols, and Flavan-3-ols
2.3.4. Analysis of the Coumarins and Furanocoumarins
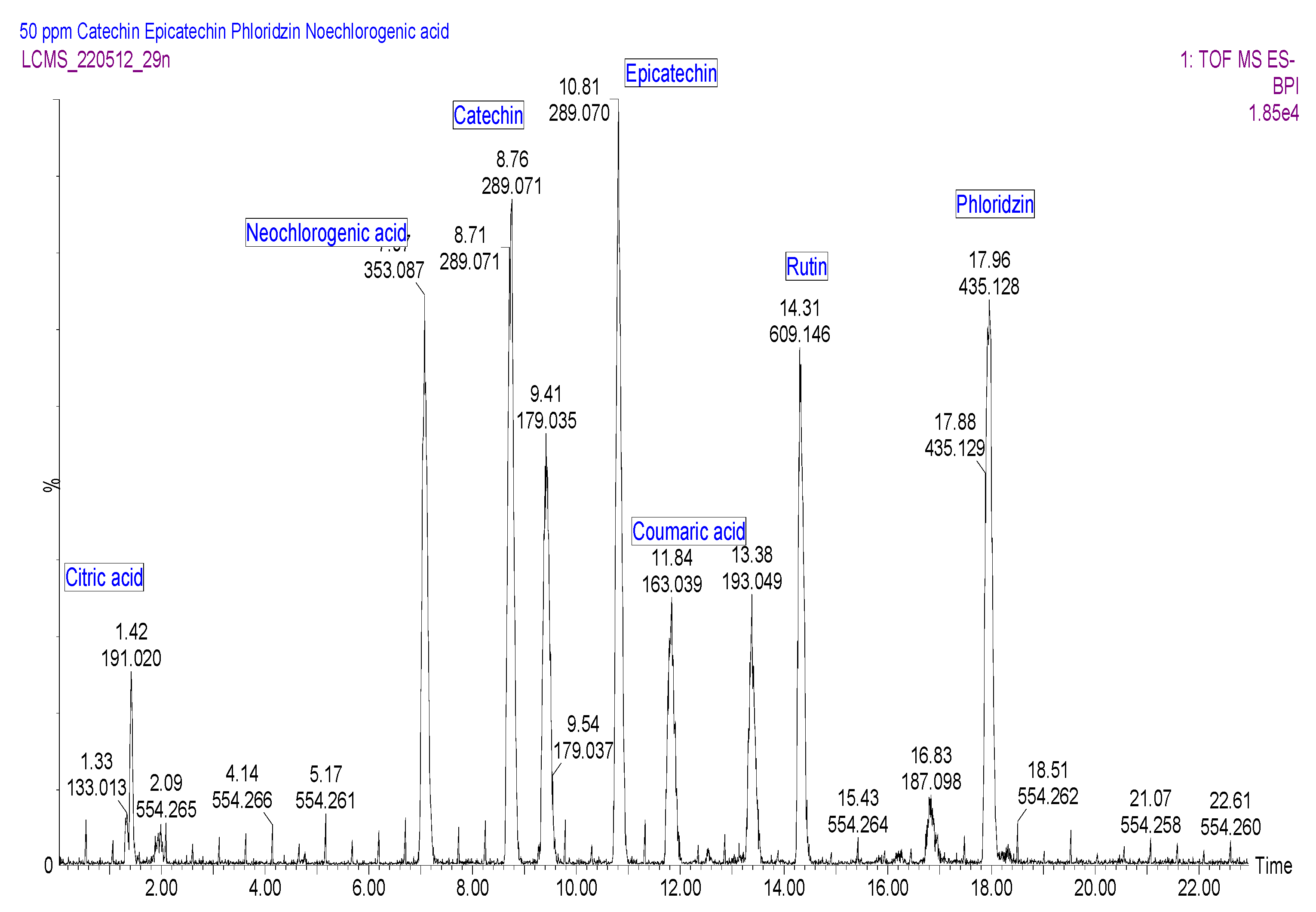
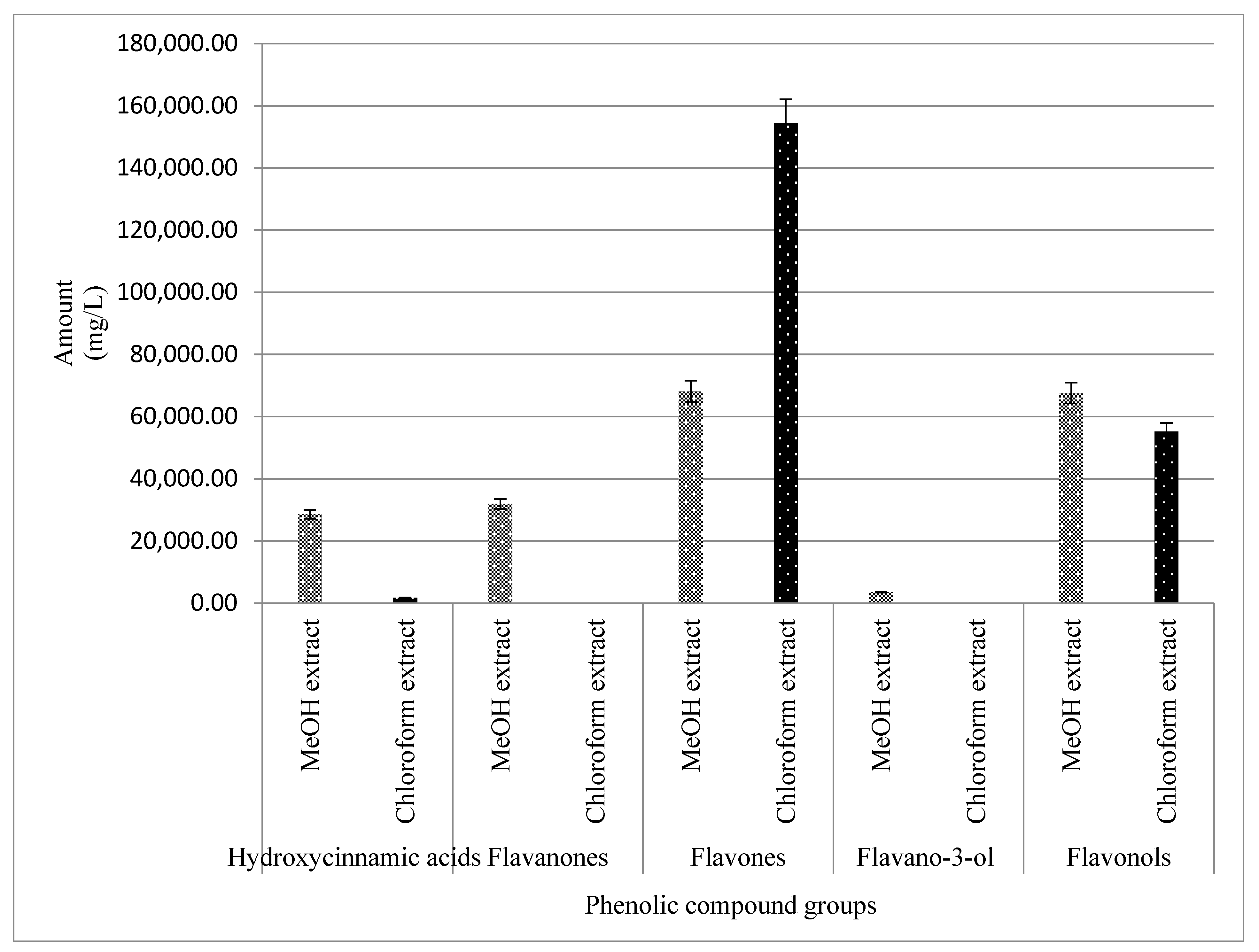
2.3.5. UPLC-QTOF-MS Quantitation of the Phenolic Compounds
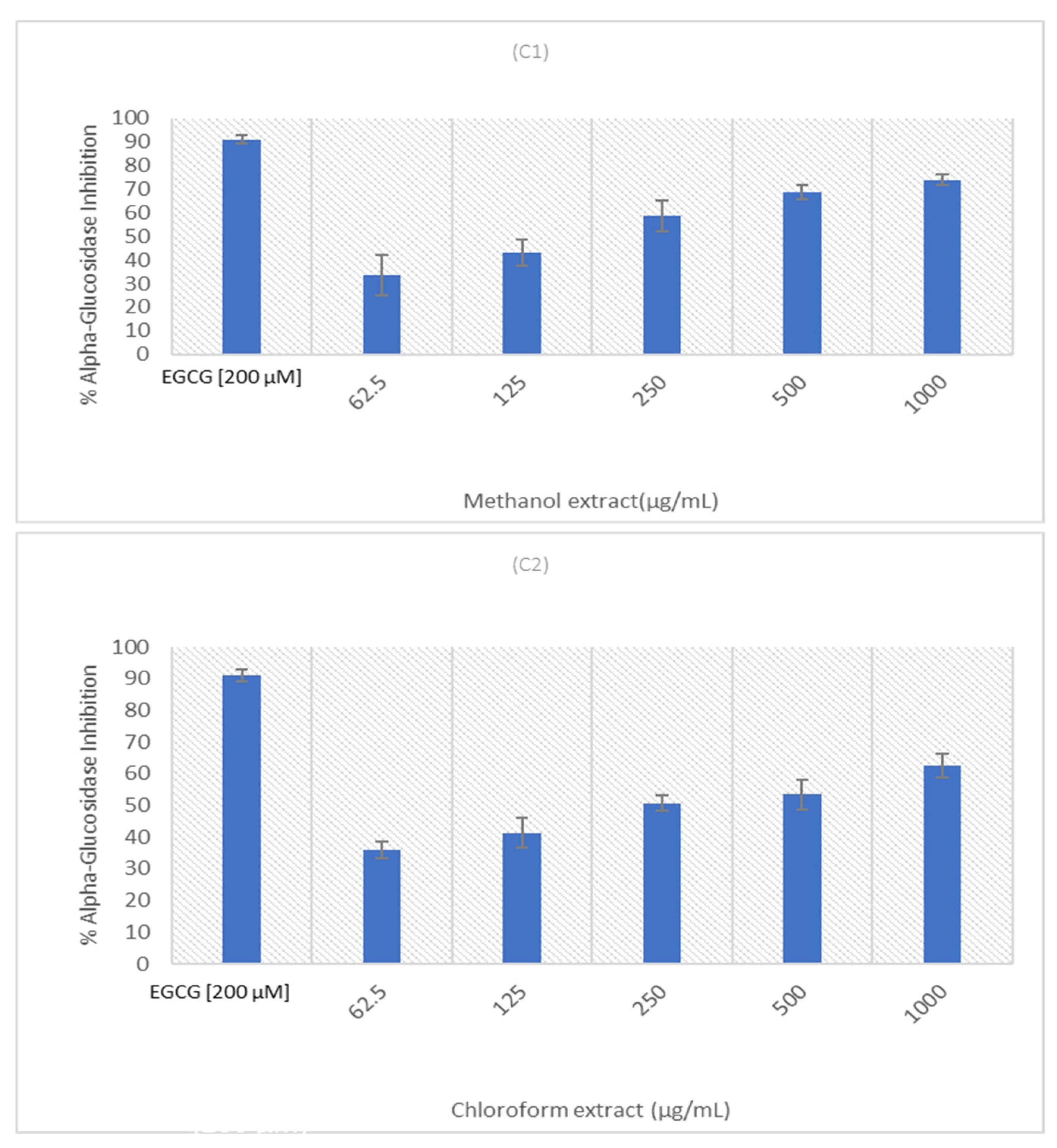
2.4. Alpha-Glucosidase Inhibition
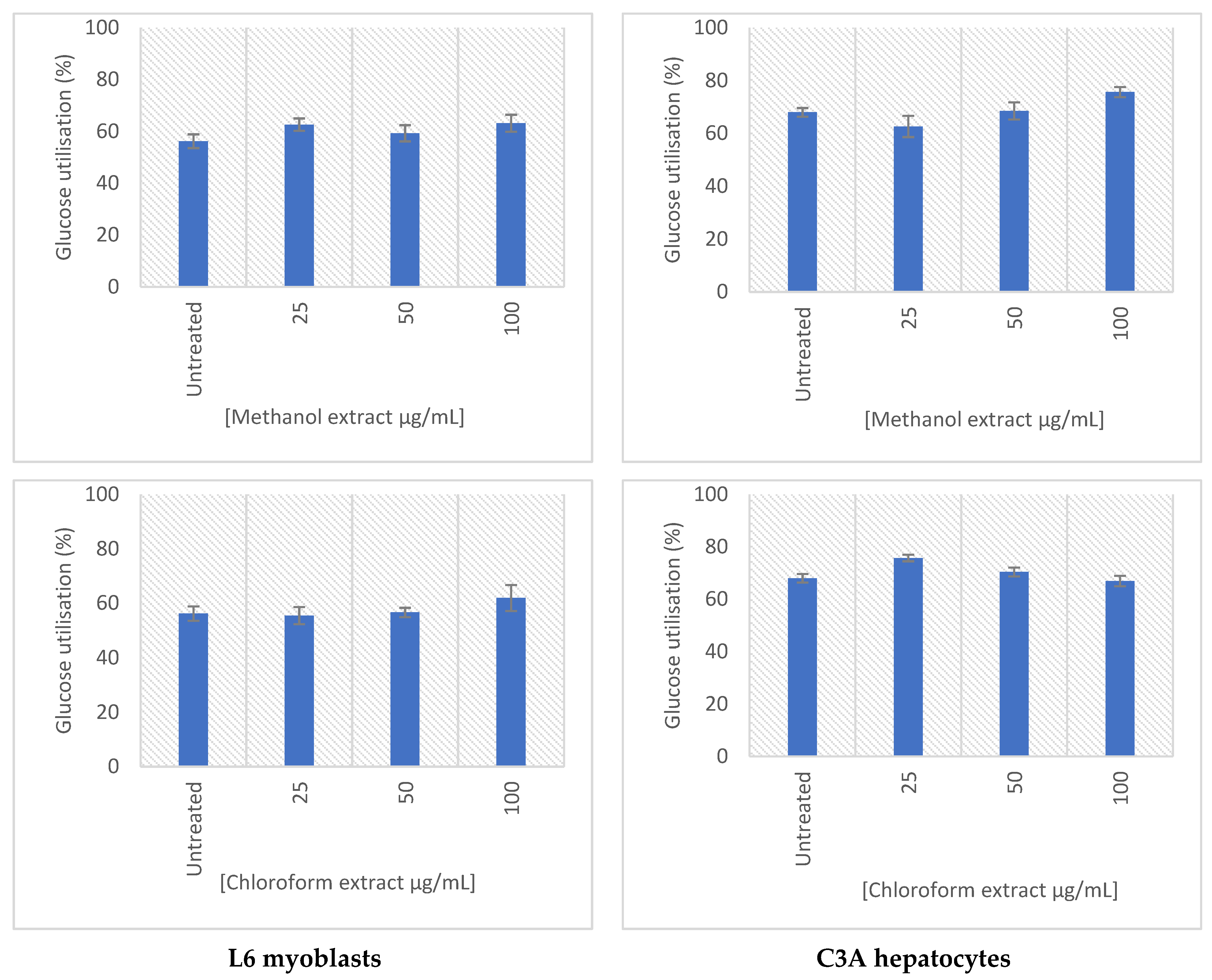
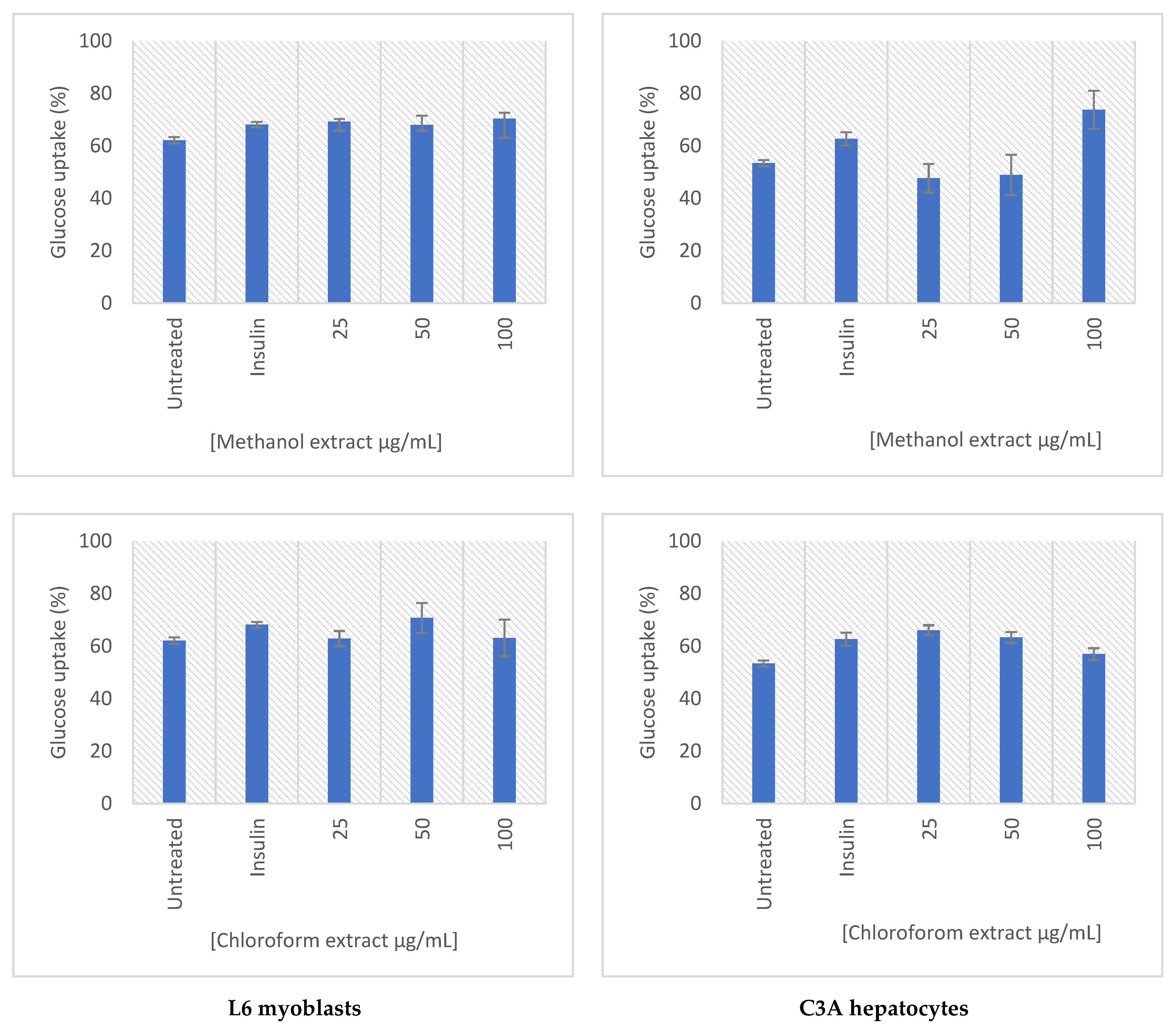
2.5. Glucose Uptake
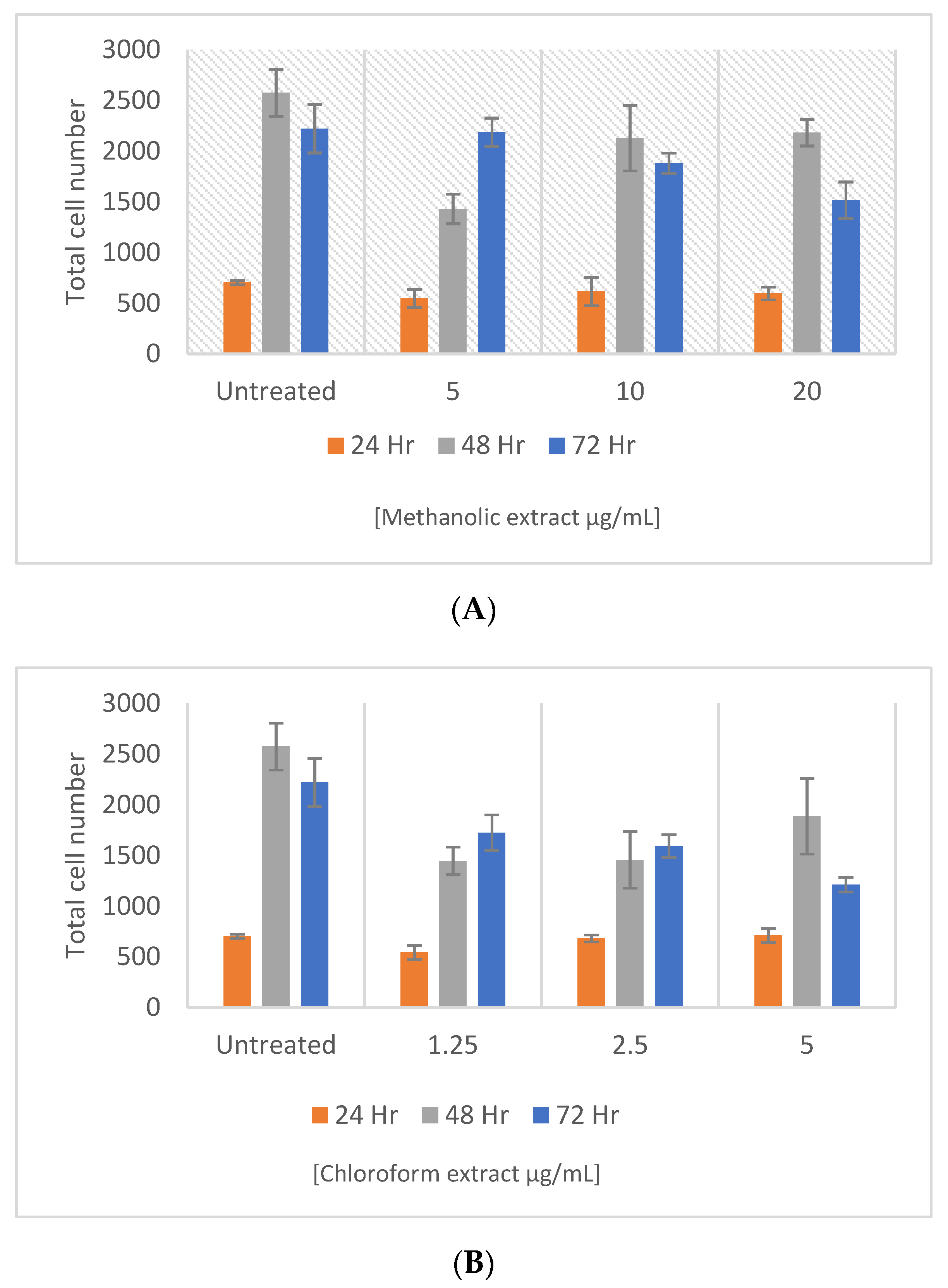
2.6. Pancreatic β-Cell Proliferation
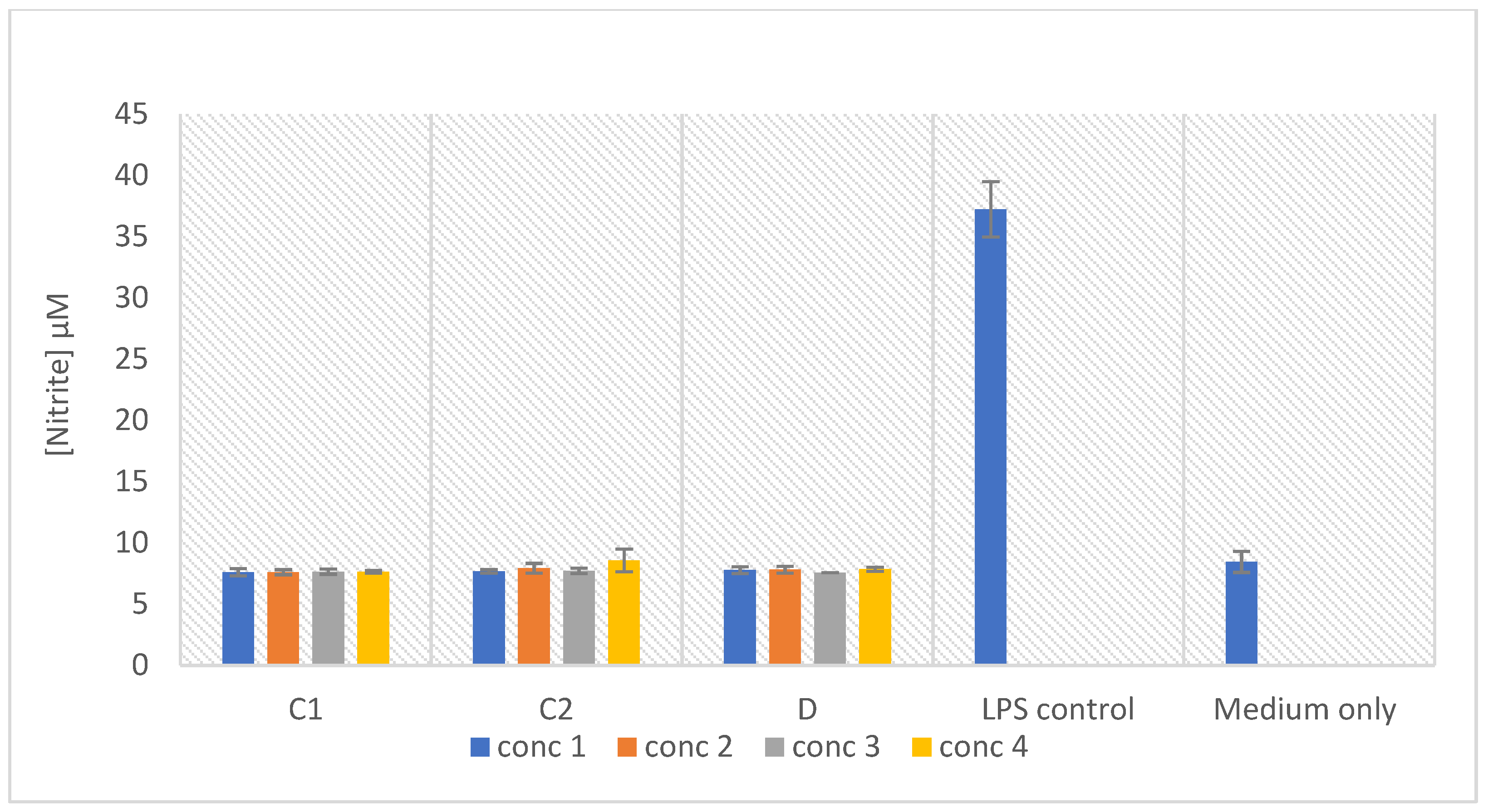
2.7. In Vitro Anti-Inflammatory Activity
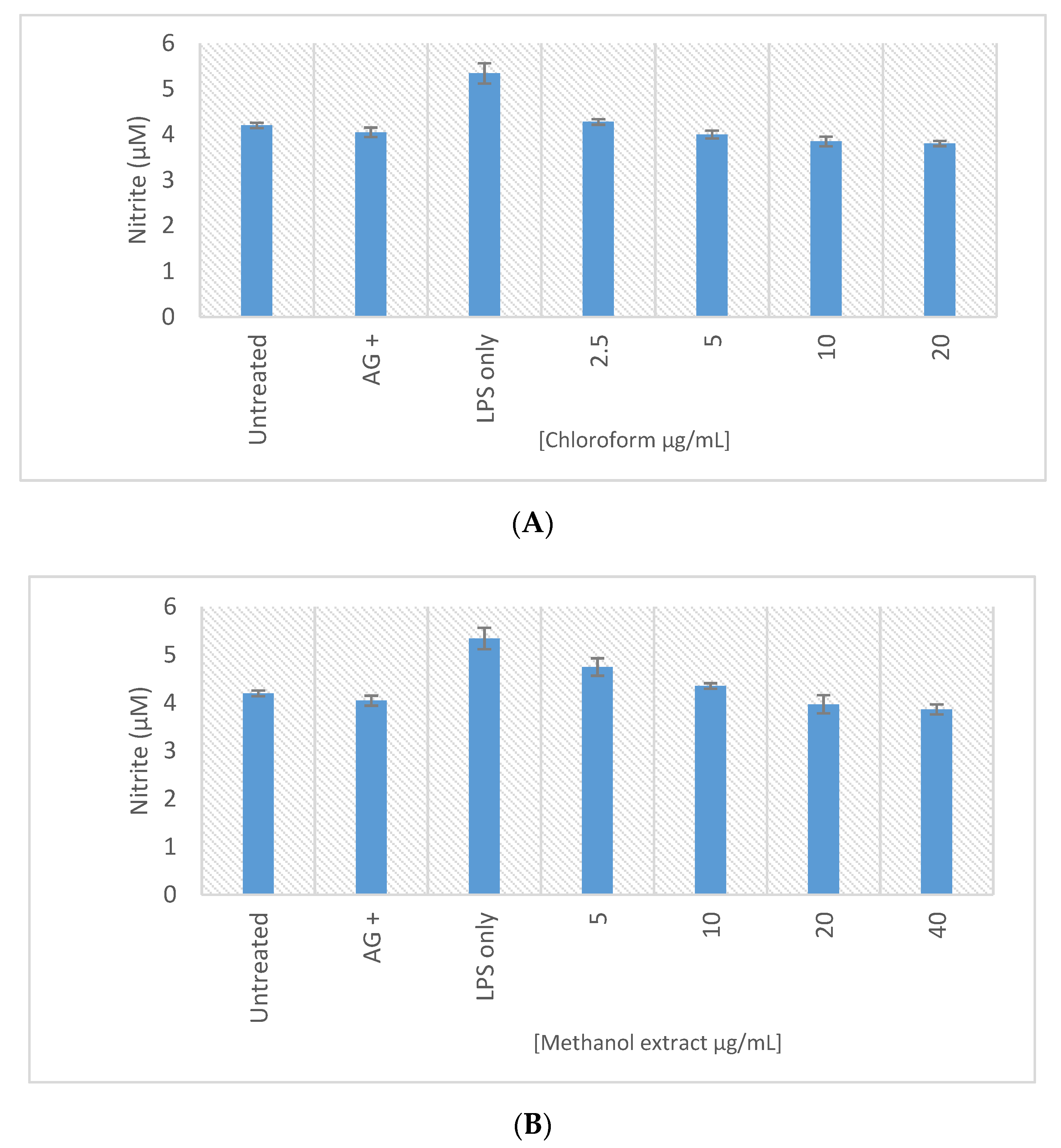
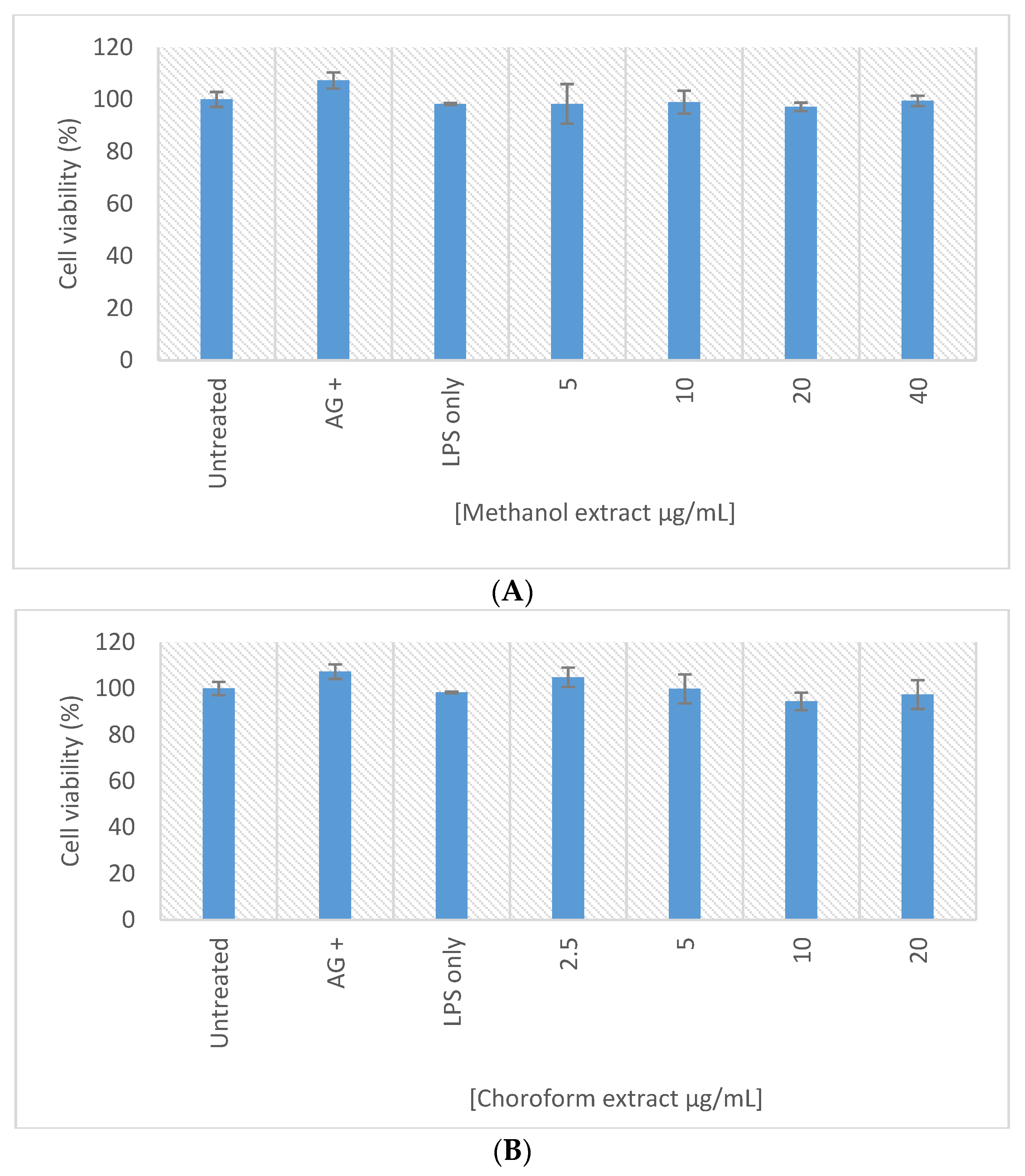
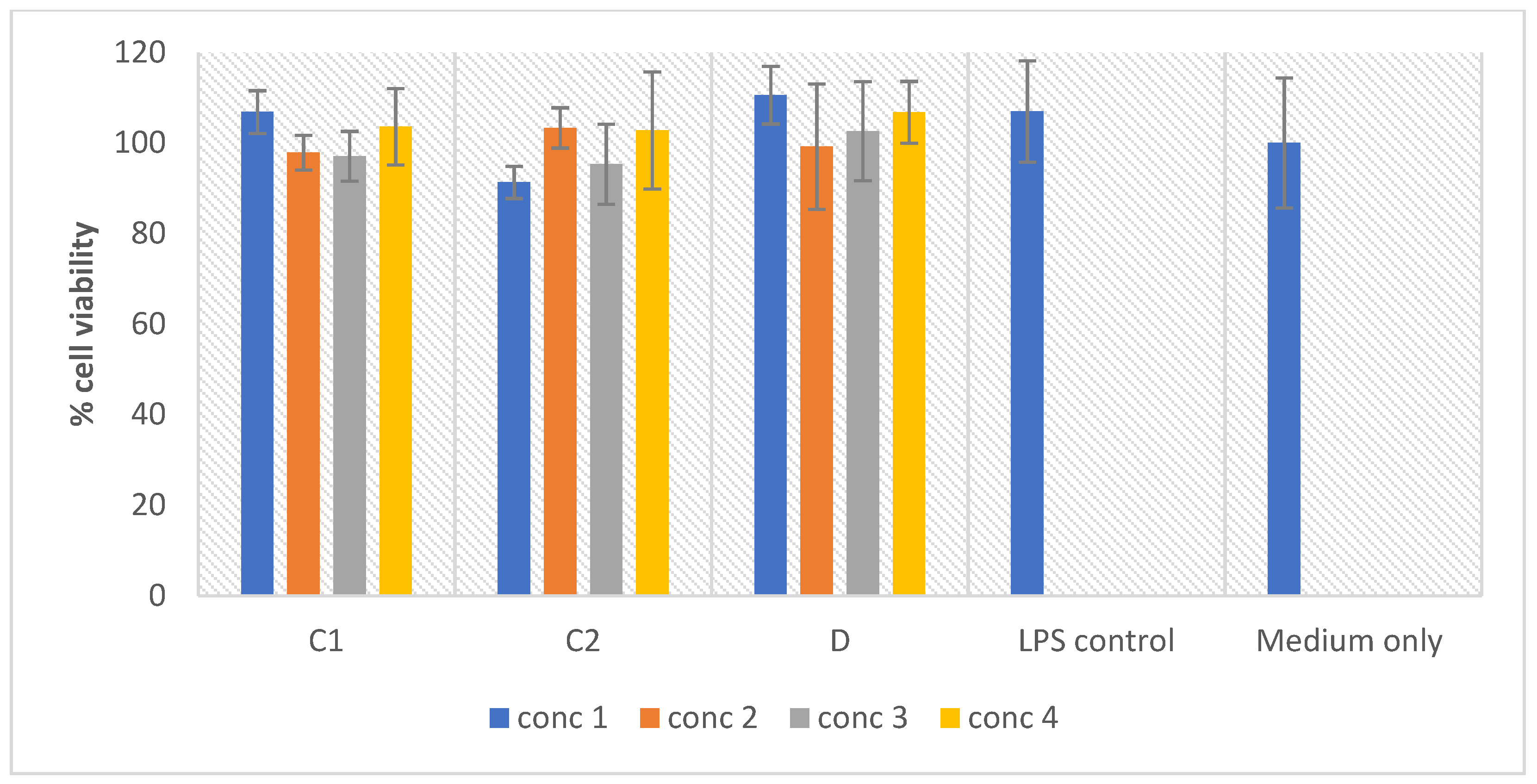
2.8. In Vitro Macrophage Activation Screening
3. Materials and Methods
3.1. Plant Collection and Identification
3.2. Plant Extraction
3.3. Determination of the Total Polyphenol and Flavonol Contents
3.4. Estimation of the Antioxidant Capacity
3.5. Alpha-Glucosidase Inhibition
3.6. Glucose Uptake
3.6.1. Cell Line Maintenance
3.6.2. Glucose Utilization (A) and Uptake (B)
3.7. 3-4,5-dimethylthiazol-2,5-diphenyltetrazolium Bromide (MTT) Assay
3.8. Pancreatic β-Cell Proliferation
3.8.1. Cell Line Maintenance
3.8.2. β-Cell Proliferation Assay
3.9. In Vitro Macrophage Activation
3.10. UPLC-ESI-QTOF-MS Analysis of the Methanol and Chloroform Extracts of E. africanus
3.11. Identification of the Compounds Using UPLC-ESI-QTOF-MS
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Wang, W.; Xu, H.; Chen, H.; Tai, K.; Liu, F.; Gao, Y. In vitro antioxidant, anti-diabetic and antilipemic potentials of quercetagetin extracted from marigold (Tagetes erecta L.) inflorescence residues. J. Food Sci. Technol. 2016, 53, 2614–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Diabetes Federation. IDF Diabetes Atlas 2017, 8th ed.; International Diabetes Federation: Brussels, Belgium, 2017. [Google Scholar]
- Hoyas, I.; Leon-Sanz, M. Nutritional Challenges in Metabolic Syndrome. J. Clin. Med. 2019, 8, 1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45. [Google Scholar] [PubMed]
- Upadhyay, J.; Polyzos, S.A.; Perakakis, N.; Thakkar, B.; Paschou, S.A.; Katsiki, N.; Underwood, P.; Park, K.-H.; Seufert, J.; Kang, E.S.; et al. Pharmacotherapy of type 2 diabetes: An update. Metabolism 2018, 78, 13–42. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Overview of food products and dietary constituents with antidiabetic properties and their putative mechanisms of action: A natural approach to complement pharmacotherapy in the management of diabetes. Mol. Nutr. Food Res. 2013, 58, 61–78. [Google Scholar] [CrossRef]
- Hossain, M.K.; Dayem, A.A.; Han, J.; Yin, Y.; Kim, K.; Saha, S.K.; Yang, G.-M.; Choi, H.Y.; Cho, S.-G. Molecular Mechanisms of the Anti-Obesity and Anti-Diabetic Properties of Flavonoids. Int. J. Mol. Sci. 2016, 17, 569. [Google Scholar] [CrossRef] [Green Version]
- Chang, A.M.; Smith, M.J.; Bloem, C.J.; Galecki, A.T.; Halter, J.B. Effect of lowering postprandial hyperglycemia on insulin secretion in older people with impaired glucose tolerance. Am. J. Physiol. Metab. 2004, 287, E906–E911. [Google Scholar] [CrossRef]
- Chawla, A.; Chawla, R.; Jaggi, S. Microvasular and macrovascular complications in diabetes mellitus: Distinct or continuum? Ind. J. Endocrinol. Metab. 2016, 20, 546–551. [Google Scholar] [CrossRef]
- Forcados, G.E.; James, D.B.; Sallau, A.B.; Muhammad, A.; Mabeta, P. Oxidative stress and carcinogenesis: Potential of phy-tochemicals in breast cancer therapy. Nutr. Cancer. 2017, 69, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Sil, P. Role of Plant-Derived Polyphenols in Reducing Oxidative Stress-Mediated Diabetic Complications. React. Oxyg. Species 2018, 5, 15–34. [Google Scholar] [CrossRef]
- Al-Aubaidy, H.A.; Jelinek, H.F. Oxidative DNA damage: Antioxidant response in postprandial hyperglycaemia in type 2 diabetes mellitus. Brit. J. Diab. Vasc. Dis. 2011, 11, 53–54. [Google Scholar] [CrossRef] [Green Version]
- Butkowski, E.G.; Jelinek, H.F. Hyperglycaemia, oxidative stress and inflammation markers. Redox Rep. 2017, 22, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, K.; Akash, M.S.H. Mechanism of generation of oxidative stress and pathophysiology of type 2 diabetes mellitus: How are they interlinked? J. Cell Biochem. 2017, 118, 3577–3585. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Donath, M.Y.; Mandrup-Poulsen, T. Role of IL-1beta in type 2 diabetes. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 314–321. [Google Scholar] [CrossRef]
- Rains, J.L.; Jain, S.K. Oxidative stress, insulin signaling, and diabetes. Free. Radic. Biol. Med. 2011, 50, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donath, M.Y.; Dalmas, É.; Sauter, N.S.; Böni-Schnetzler, M. Inflammation in Obesity and Diabetes: Islet Dysfunction and Therapeutic Opportunity. Cell Metab. 2013, 17, 860–872. [Google Scholar] [CrossRef] [Green Version]
- Engwa, G.A. Free Radicals and the Role of Plant Phytochemicals as Antioxidants Against Oxidative Stress-Related Diseases. Phytochemicals: Source of Antioxidants and Role in Disease Prevention. BoD-Books Demand 2018, 7, 49–74. [Google Scholar]
- Gerber, P.A.; Rutter, G.A. The Role of Oxidative Stress and Hypoxia in Pancreatic Beta-Cell Dysfunction in Diabetes Mellitus. Antioxid. Redox Signal. 2017, 26, 501–518. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Zhao, C.; Guven, E.C.; Paoli, P.; Simal-Gandara, J.; Ramkumar, K.M.; Wang, S.; Buleu, F.; Pah, A.; Turi, V.; et al. Dietary polyphenols as antidiabetic agents: Advances and opportunities. Food Front. 2020, 1, 18–44. [Google Scholar] [CrossRef] [Green Version]
- Okaiyeto, K.; Kerebba, N.; Rautenbach, F.; Singh, S.K.; Dua, K.; Oguntibeju, O.O. UPLC-ESI-QTOF-MS phenolic compounds identification and quantification from ethanolic extract of Myrtus communis ‘Variegatha’: In vitro antioxidant and antidiabetic potentials. Arab. J. Chem. 2022, 16, 104447. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Lebelo, S.L. Antioxidant Effects and Mechanisms of Medicinal Plants and Their Bioactive Compounds for the Prevention and Treatment of Type 2 Diabetes: An Updated Review. Oxidative Med. Cell. Longev. 2020, 2020, 1356893. [Google Scholar] [CrossRef] [PubMed]
- Rupasinghe, H.V.; Sekhon-Loodu, S.; Mantso, T.; Panayiotidis, M.I. Phytochemicals in regulating fatty acid β-oxidation: Potential underlying mechanisms and their involvement in obesity and weight loss. Pharmacol. Ther. 2016, 165, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Olvera-Sandoval, C.; Fabela-Illescas, H.E.; Fernández-Martínez, E.; Ortiz-Rodríguez, M.A.; Cariño-Cortés, R.; Ariza-Ortega, J.A.; Hernández-González, J.C.; Olivo, D.; Valadez-Vega, C.; Belefant-Miller, H.; et al. Potential Mechanisms of the Improvement of Glucose Homeostasis in Type 2 Diabetes by Pomegranate Juice. Antioxidants 2022, 11, 553. [Google Scholar] [CrossRef]
- Mahnashi, M.H.; Alqahtani, Y.S.; Alyami, B.A.; Alqarni, A.O.; Alqahl, S.A.; Ullah, F.; Sadiq, A.; Zeb, A.; Ghufran, M.; Kuraev, A.; et al. HPLC-DAD phenolics analysis, α-glucosidase, α-amylase inhibitory, molecular docking and nutritional profiles of Persicaria hydropiper L. BMC Complement. Altern. Med. 2022, 22, 26. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.; Pham, B.; Le, L. Bioactive Compounds in Anti-Diabetic Plants: From Herbal Medicine to Modern Drug Discovery. Biology 2020, 9, 252. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Silva, A.M.; Saraiva, S.C.; Sobral, A.J.; Cardoso, S.M. Characterization of phenolic constituents and evaluation of antioxidant properties of leaves and stems of Eriocephalus africanus. Arab. J. Chem. 2018, 11, 62–69. [Google Scholar] [CrossRef] [Green Version]
- van Wyk, B.E.; van Oudtshoorn, B.; Gericke, N. Medicinal Plants of South Africa; Briza Publications: Pretoria, South Africa, 1997. [Google Scholar]
- Behiry, S.I.; El-Hefny, M.; Salem, M.Z.M. Toxicity effects of Eriocephalus africanus L. leaf essential oil against some molecularly identified phytopathogenic bacterial strains. Nat. Prod. Res. 2020, 34, 3394–3398. [Google Scholar] [CrossRef]
- Magura, J.; Moodley, R.; Maduray, K.; Mackraj, I. Phytochemical constituents and in vitro anticancer screening of isolated compounds from Eriocephalus africanus. Nat. Prod. Res. 2021, 35, 4173–4176. [Google Scholar] [CrossRef]
- Khalil, N.; Elhady, S.S.; Diri, R.M.; Fekry, M.I.; Bishr, M.; Salama, O.; El-Zalabani, S.M. Salicylic Acid Spraying Affects Sec-ondary Metabolites and Radical Scavenging Capacity of Drought-Stressed Eriocephalus africanus L. Agronomy 2022, 12, 2278. [Google Scholar] [CrossRef]
- Babbar, N.; Oberoi, H.S.; Sandhu, S.K.; Bhargav, V.K. Influence of different solvents in extraction of phenolic compounds from vegetable residues and their evaluation as natural sources of antioxidants. J. Food Sci. Technol. 2014, 51, 2568–2575. [Google Scholar] [CrossRef] [Green Version]
- Piasecka, A.; Jedrzejczak-Rey, N.; Bednarek, P. Secondary metabolites in plant innate immunity: Conserved function of di-vergent chemicals. New Phytol. 2015, 206, 948–964. [Google Scholar] [CrossRef] [PubMed]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: An overview. Futur. J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef]
- Ullrich, C.I.; Aloni, R.; Saeed, M.E.; Ullrich, W.; Efferth, T. Comparison between tumors in plants and human beings: Mechanisms of tumor development and therapy with secondary plant metabolites. Phytomedicine 2019, 64, 153081. [Google Scholar] [CrossRef]
- Fakhri, S.; Moradi, S.Z.; Farzaei, M.H.; Bishayee, A. Modulation of dysregulated cancer metabolism by plant secondary metabolites: A mechanistic review. Semin. Cancer Biol. 2020, 80, 276–305. [Google Scholar] [CrossRef]
- Cosme, P.; Rodríguez, A.B.; Espino, J.; Garrido, M. Plant Phenolics: Bioavailability as a Key Determinant of Their Potential Health-Promoting Applications. Antioxidants 2020, 9, 1263. [Google Scholar] [CrossRef]
- Zekeya, N.; Ibrahim, M.; Mamiro, B.; Ndossi, H.; Kilonzo, M.; Mkangara, M.; Chacha, M.; Chilongola, J.; Kideghesho, J. Potential of natural phenolic antioxidant compounds from Bersama abyssinica (Meliathacea) for treatment of chronic diseases. Saudi J. Biol. Sci. 2022, 29, 103273. [Google Scholar] [CrossRef]
- Archana, T.; Soumya, K.; James, J.; Sudhakaran, S. Root extracts of Anacardium occidentale reduce hyperglycemia and oxidative stress in vitro. Clin. Phytosci. 2021, 7, 57. [Google Scholar] [CrossRef]
- Harborne, J.B. Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis; Chapman and Hall: London, UK, 1998. [Google Scholar]
- Kerebba, N.; Oyedeji, A.O.; Byamukama, R.; Kuria, S.K.; Oyedeji, O.O. UHPLC-ESI-QTOF-MS/MS characterisation of phenolic compounds from Tithonia diversifolia (Hemsl.) A. gray and antioxidant activity. ChemistrySelect 2022, 7, e202104406. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free. Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef] [PubMed]
- Magura, J. The Effect of Isolated and Nanoencapsulated Flavonoids from Eriocephalus africanus on Apoptotic Factors and Microrna Expression in Cancer. Doctoral Dissertation, University of KwaZulu-Natal, Durban, South Africa, 2020. [Google Scholar]
- Clifford, M.N.; Knight, S.; Kuhnert, N. Discriminating between the six isomers of dicaffeoylquinic acid by LC-MS. J. Agric. Food Chem. 2005, 53, 3821. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.G.; Zhang, L.J.; Li, C.B.; Lan, Y.Y.; Wang, A.M.; Huang, Y.; Zhen, L.; Fu, X.Z.; Zhou, W.; Qi, X.L.; et al. Rapid screening and identification of caffeic acid and its esters in Erigeron breviscapus by ultra-performance liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2533–2541. [Google Scholar] [CrossRef]
- Schram, K.; Miketova, P.; Slanina, J.; Humpa, O.; Taborska, E. Mass spectrometry of 1,3-and 1,5-dicaffeoylquinic acids. J. Mass Spectrom. 2004, 39, 384–395. [Google Scholar] [CrossRef]
- Park, J.S.; Woo, J.W.; Choi, G.-H.; Choi, D.-S.; Jung, M.Y. Chlorogenic Acid Profiles and Antioxidant Potentials of 17 Sweet Potato Varieties Cultivated in Korea: Impact of Extraction Condition and Classification by Hierarchical Clustering Analysis. J. Food Chem. Nanotechnol. 2015, 1, 3–12. [Google Scholar] [CrossRef]
- Tsimogiannis, D.; Samiotaki, M.; Panayotou, G.; Oreopoulou, V. Characterization of Flavonoid Subgroups and Hydroxy Substitution by HPLC-MS/MS. Molecules 2007, 12, 593–606. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Ye, M.; Liu, M.; Kong, D.; Shi, R.; Shi, X.; Zhang, K.; Wang, Q.; Lantong, Z. A practical strategy for the characterization of coumarins in Radix Glehniae by liquid chromatography coupled with triple quadrupole-linear ion trap mass spectrometry. J. Chromatogr. A 2010, 1217, 4587–4600. [Google Scholar] [CrossRef]
- Göttingerová, M.; Kumšta, M.; Rampáčková, E.; Kiss, T.; Nečas, T. Analysis of phenolic compounds and some important analytical properties in selected apricot genotypes. HortScience 2021, 56, 1446–1452. [Google Scholar] [CrossRef]
- Bhatia, A.; Singh, B.; Arora, R.; Arora, S. In vitro evaluation of the α-glucosidase inhibitory potential of methanolic extracts of traditionally used antidiabetic plants. BMC Complement. Altern. Med. 2019, 19, 74. [Google Scholar] [CrossRef] [Green Version]
- Nair, S.S.; Kavrekar, V.; Mishra, A. In vitro studies on alpha amylase and alpha glucosidase inhibitory activities of selected plant extracts. Eur. J. Exp. Biol. 2013, 3, 128–132. [Google Scholar]
- Ojo, O.A.; Amanze, J.C.; Oni, A.I.; Grant, S.; Iyobhebhe, M.; Elebiyo, T.C.; Rotimi, D.; Asogwa, N.T.; Oyinloye, B.E.; Ajiboye, B.O.; et al. Antidiabetic activity of avocado seeds (Persea americana Mill.) in diabetic rats via activation of PI3K/AKT signaling pathway. Sci. Rep. 2022, 12, 2919. [Google Scholar] [CrossRef] [PubMed]
- Daou, M.; Elnaker, N.A.; Ochsenkühn, M.A.; Amin, S.A.; Yousef, A.F.; Yousef, L.F. In vitro α-glucosidase inhibitory activity of Tamarix nilotica shoot extracts and fractions. PLoS ONE 2022, 17, e0264969. [Google Scholar] [CrossRef] [PubMed]
- Jadalla, B.M.I.S.; Moser, J.J.; Sharma, R.; Etsassala, N.G.E.R.; Egieyeh, S.A.; Badmus, J.A.; Marnewick, J.L.; Beukes, D.; Cupido, C.N.; Hussein, A.A. In Vitro Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities and Antioxidant Capacity of Helichrysum cymosum and Helichrysum pandurifolium Schrank Constituents. Separations 2022, 9, 190. [Google Scholar] [CrossRef]
- Arshad, M.; Chaudhary, A.R.; Mumtaz, M.W.; Raza, S.A.; Ahmad, M.; Mukhtar, H.; Bashir, R. Polyphenol fingerprinting and hypoglycemic attributes of optimized Cycas circinalis leaf extracts. J. Sci. Food Agric. 2021, 101, 1530–1537. [Google Scholar] [CrossRef]
- William, J.; John, P.; Mumtaz, M.W.; Ch, A.R.; Adnan, A.; Mukhtar, H.; Sharif, S.; Raza, S.A.; Akhtar, M.T. Antioxidant activity, α-glucosidase inhibition and phytochemical profiling of Hyophorbela genicaulis leaf extracts. PeerJ 2019, 7, e7022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niaz, A.; Adnan, A.; Bashir, R.; Mumtaz, M.W.; Raza, S.A.; Rashid, U.; Tan, C.P.; Tan, T.B. The In Vitro α-Glucosidase Inhibition Activity of Various Solvent Fractions of Tamarix dioica and 1H-NMR Based Metabolite Identification and Molecular Docking Analysis. Plants 2021, 10, 1128. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.U.; Mumtaz, M.W.; Mukhtar, H.; Rashid, U.; Akhtar, M.T.; Raza, S.A.; Nadeem, M. UHPLC-QTOF-MS/MS based phytochemical characterization and anti-hyperglycemic prospective of hydro-ethanolic leaf extract of Butea monosperma. Sci. Rep. 2020, 10, 3530. [Google Scholar] [CrossRef] [Green Version]
- William, J.; John, P.; Mumtaz, M.W.; Ch, A.R.; Adnan, A.; Mukhtar, H.; Sharif, S.; Raza, S.A. Antioxidant activity, hypoglycemic potential and metabolite profiling of Hyophorbe indica leaf extract. Pak. J. Pharm. Sci. 2018, 31, 2737–2742. [Google Scholar]
- Tundis, R.; Loizzo, M.R.; Menichini, F. Natural products as alpha-amylase and alpha-glucosidase inhibitors and their hypo-glycaemic potential in the treatment of diabetes: An update. Mini Rev. Med. Chem. 2010, 4, 315–331. [Google Scholar] [CrossRef]
- Rose, D.R.; Chaudet, M.M.; Jones, K. Structural Studies of the Intestinal α-Glucosidases, Maltase-glucoamylase and Sucrase-isomaltase. JPGN 2018, 66, S11–S13. [Google Scholar] [CrossRef] [PubMed]
- Roskar, I.; Molek, P.; Vodnik, M.; Stempelj, M.; Strukelj, B.; Lunder, M. Peptide modulators of alpha-glucosidase. J. Diabetes Investig. 2015, 6, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Bian, Y.; Liu, T.; Xu, Z.; Song, Z.; Wang, F.; Li, T.; Li, S. Antioxidant potential and in vitro inhibition of starch digestion of flavonoids from Crataegus pinnatifida. Heliyon 2022, 8, e11058. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, M.W.; Al-Zuaidy, M.H.; Abdul Hamid, A.; Danish, M.; Akhtar, M.T.; Mukhtar, H. Metabolite profiling and inhibitory properties of leaf extracts of Ficus benjamina towards α-glucosidase and α-amylase. Int. J. Food Prop. 2018, 21, 1560–1574. [Google Scholar] [CrossRef] [Green Version]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.; Sousa, J.L.; Tome, S.M.; Ramos, M.J.; Silva, A.M.; Fernandes, P.A.; Fernandes, E. α-Glucosidase inhibition by flavonoids: An in vitro and in silico structure–activity relationship study. J. Enzym. Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef] [Green Version]
- Cox, D.J.; Fang, K.; McCall, A.L.; Conaway, M.R.; Banton, T.A.; Moncrief, M.A.; Diamond, A.M.; Taylor, A.G. Behavioral Strategies to Lower Post-prandial Glucose in Those with Type 2 Diabetes May Also Lower Risk of Coronary Heart Disease. Diabetes Ther. 2019, 10, 277–281. [Google Scholar] [CrossRef] [Green Version]
- You, Q.; Chen, F.; Wang, X.; Jiang, Y.; Lin, S. Anti-diabetic activities of phenolic compounds in muscadine against alpha-glucosidase and pancreatic lipase. LWT-Food Sci. Technol. 2012, 46, 164–168. [Google Scholar] [CrossRef]
- EscandÓn-Rivera, S.; González-Andrade, M.; Bye, R.; Linares, E.; Navarrete, A.; Mata, R. α-Glucosidase inhibitors from Brickellia cavanillesii. J. Nat. Prod. 2012, 75, 968–974. [Google Scholar] [CrossRef]
- Shibano, M.; Kakutani, K.; Taniguchi, M.; Yasuda, M.; Baba, K. Antioxidant constituents in the dayflower (Commelina communis L.) and their α-glucosidase-inhibitory activity. J. Nat. Med. 2008, 62, 349–353. [Google Scholar] [CrossRef]
- Funke, I.; Melzig, M.F. Effect of different phenolic compounds on α-amylase activity: Screening by microplate-reader based kinetic assay. Pharmazie 2005, 60, 796–797. [Google Scholar]
- Kim, J.-S.; Kwon, C.-S.; Son, K.H. Inhibition of Alpha-glucosidase and Amylase by Luteolin, a Flavonoid. Biosci. Biotechnol. Biochem. 2000, 64, 2458–2461. [Google Scholar] [CrossRef]
- Zhang, W.-Y.; Lee, J.-J.; Kim, Y.; Kim, I.-S.; Han, J.-H.; Lee, S.-G.; Ahn, M.-J.; Jung, S.-H.; Myung, C.-S. Effect of Eriodictyol on Glucose Uptake and Insulin Resistance in Vitro. J. Agric. Food Chem. 2012, 60, 7652–7658. [Google Scholar] [CrossRef]
- Thao, N.P.; Luyen, B.T.T.; Tai, B.H.; Yang, S.Y.; Jo, S.H.; Cuong, N.X.; Nam, N.H.; Kwon, Y.I.; Van Minh, C.; Kim, Y.H. Rat intestinal sucrase inhibition of constituents from the roots of Rosa rugosa Thunb. Bioorg. Med. Chem. Lett. 2014, 24, 1192–1196. [Google Scholar] [CrossRef]
- Zhao, J.; Zhou, X.-W.; Chen, X.-B.; Wang, Q.-X. α-glucosidase inhibitory constituents from Toona sinensis. Chem. Nat. Compd. 2009, 45, 244–246. [Google Scholar] [CrossRef]
- Wansi, J.D.; Lallemand, M.-C.; Chiozem, D.D.; Toze, F.A.A.; Mbaze, L.M.; Naharkhan, S.; Iqbal, M.C.; Tillequin, F.; Wandji, J.; Fomum, Z.T. α-Glucosidase inhibitory constituents from stem bark of Terminalia superba (Combretaceae). Phytochemistry 2007, 68, 2096–2100. [Google Scholar] [CrossRef]
- Asghari, B.; Salehi, P.; Sonboli, A.; Ebrahimi, S.N. Flavonoids from Salvia chloroleuca with α-amylsae and α-glucosidase inhibitory effect. Iran. J. Pharm. Res. IJPR 2015, 14, 609. [Google Scholar]
- Koistinen, H.A.; Galuska, D.; Chibalin, A.V.; Yang, J.; Zierath, J.R.; Holman, G.D.; Wallberg-Henriksson, H. 5-Amino-Imidazole Carboxamide Riboside Increases Glucose Transport and Cell-Surface GLUT4 Content in Skeletal Muscle from Subjects with Type 2 Diabetes. Diabetes 2003, 52, 1066–1072. [Google Scholar] [CrossRef] [Green Version]
- Taderera, T.; Chagonda, L.S.; Gomo, E.; Katerere, D.; Shai, L.J. Annona stenophylla aqueous extract stimulate glucose uptake in established C2Cl2 muscle cell lines. Afr. Health Sci. 2019, 19, 2219–2229. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.; Ahmed, Q.U.; Soad, S.Z.M.; Latip, J.; Taher, M.; Syafiq, T.M.F.; Sarian, M.N.; Alhassan, A.M.; Zakaria, Z.A. Flavonoids from Tetracera indica Merr. induce adipogenesis and exert glucose uptake activities in 3T3-L1 adipocyte cells. BMC Complement. Altern. Med. 2017, 17, 431. [Google Scholar] [CrossRef] [Green Version]
- Moin, A.S.; Butler, A.E. Alterations in Beta Cell Identity in Type 1 and Type 2 Diabetes. Curr. Diabetes Rep. 2019, 19, 83. [Google Scholar] [CrossRef] [Green Version]
- Tomita, T. Apoptosis in pancreatic β-islet cells in Type 2 diabetes. Bosn. J. Basic Med. Sci. 2016, 16, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, H.; Saisho, Y.; Inaishi, J.; Itoh, H. Revisiting Regulators of Human β-cell Mass to Achieve β-cell–centric Approach Toward Type 2 Diabetes. J. Endocr. Soc. 2021, 5, bvab128. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.E.; McKenzie, M.D.; Angstetra, E.; Campbell, P.D.; Kay, T.W. Beta cell apoptosis in diabetes. Apoptosis 2009, 14, 1389. [Google Scholar] [CrossRef] [PubMed]
- Pringle, N.A.; van de Venter, M.; Koekemoer, T.C. Comprehensive in vitro antidiabetic screening of Aspalathus linearis using a target-directed screening platform and cellomics. Food Funct. 2020, 12, 1020–1038. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Shailendra, G.; Ribnicky, D.M.; Burk, D.; Karki, N.; Wang, M.Q. An extract of Artemisia dracunculus L. stimulates insulin secretion from β cells, activates AMPK and suppresses inflammation. J. Ethnopharmacol. 2015, 170, 98–105. [Google Scholar] [CrossRef]
- Đorđević, M.; Grdović, N.; Mihailović, M.; Jovanović, J.A.; Uskoković, A.; Rajić, J.; Sinadinović, M.; Tolić, A.; Mišić, D.; Šiler, B.; et al. Centaurium erythraea extract improves survival and functionality of pancreatic beta-cells in diabetes through multiple routes of action. J. Ethnopharmacol. 2019, 242, 112043. [Google Scholar] [CrossRef]
- Oh, Y.S. Plant-Derived Compounds Targeting Pancreatic Beta Cells for the Treatment of Diabetes. Evid.-Based Complement. Altern. Med. 2015, 2015, 629863. [Google Scholar] [CrossRef] [Green Version]
- Smoak, P.; Burke, S.J.; Collier, J.J. Botanical Interventions to Improve Glucose Control and Options for Diabetes Therapy. SN Compr. Clin. Med. 2021, 3, 2465–2491. [Google Scholar] [CrossRef]
- Li, K.; Yao, F.; Xue, Q.; Fan, H.; Yang, L.; Li, X.; Sun, L.; Liu, Y. Inhibitory effects against α-glucosidase and α-amylase of the flavonoids-rich extract from Scutellaria baicalensis shoots and interpretation of structure–activity relationship of its eight flavonoids by a refined assign-score method. Chem. Central J. 2018, 12, 82. [Google Scholar] [CrossRef]
- Lapidot, T.; Walker, M.D.; Kanner, J. Antioxidant and Prooxidant Effects of Phenolics on Pancreatic β-Cells in Vitro. J. Agric. Food Chem. 2002, 50, 7220–7225. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raziyeva, K.; Kim, Y.; Zharkinbekov, Z.; Kassymbek, K.; Jimi, S.; Saparov, A. Immunology of Acute and Chronic Wound Healing. Biomolecules 2021, 11, 700. [Google Scholar] [CrossRef] [PubMed]
- Ghuman, S.; Ncube, B.; Finnie, J.F.; McGaw, L.J.; Njoya, E.M.; Coopoosamy, R.M.; Van Staden, J. Antioxidant, anti-inflammatory and wound healing properties of medicinal plant extracts used to treat wounds and dermatological disorders. S. Afr. J. Bot. 2019, 126, 232–240. [Google Scholar] [CrossRef]
- Kusminski, C.M.; Shetty, S.; Orci, L.; Unger, R.H.; Scherer, P.E. Diabetes and apoptosis: Lipotoxicity. Apoptosis 2009, 14, 1484–1495. [Google Scholar] [CrossRef]
- Ghemrawi, R.; Battaglia-Hsu, S.-F.; Arnold, C. Endoplasmic Reticulum Stress in Metabolic Disorders. Cells 2018, 7, 63. [Google Scholar] [CrossRef] [Green Version]
- Okaiyeto, K.; Oguntibeju, O. African Herbal Medicines: Adverse Effects and Cytotoxic Potentials with Different Therapeutic Applications. Int. J. Environ. Res. Public Health 2021, 18, 5988. [Google Scholar] [CrossRef]
- Sarbadhikary, P.; George, B.P. A Review on Traditionally Used African Medicinal Plant Annickia chlorantha, Its Phytochemistry, and Anticancer Potential. Plants 2022, 11, 2293. [Google Scholar] [CrossRef]
- Okafor, J.N.; Rautenbauch, F.; Meyer, M.; Le Roes-Hill, M.; Harris, T.; Jideani, V.A. Phenolic content, antioxidant, cytotoxic and antiproliferative effects of fractions of Vigna subterraenea (L.) verdc from Mpumalanga, South Africa. Heliyon 2021, 7, e08397. [Google Scholar] [CrossRef]
- Yermakov, A.I.; Arasimov, V.V.; Yarosh, N.P. Methods of Biochemical Analysis of Plants; Agropromizdat: Leningrad, Russia, 1987. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolourization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Ngxabi, S.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. Growth characteristics, phytochemical contents, and antioxidant capacity of Trachyandra ciliata (Lf) Kunth grown in hydroponics under varying degrees of salinity. Horticulturae 2021, 7, 244. [Google Scholar] [CrossRef]
- van de Venter, M.; Roux, S.; Bungu, L.C.; Louw, J.; Crouch, N.R.; Grace, O.M.; Maharaj, V.; Pillay, P.; Sewnarian, P.; Bhagwandin, N.; et al. Antidiabetic screening and scoring of 11 plants traditionally used in South Africa. J. Ethnopharmacol. 2008, 119, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Trinder, P. Determination of glucose in blood using glucose oxidase with an alternative oxygen acceptor. Ann. Clin. Biochem. Int. J. Biochem. Lab. Med. 1969, 6, 24–27. [Google Scholar] [CrossRef]
- Twentyman, P.R.; Luscombe, M. A study of some variables in a tetrazolium dye (MTT) based assay for cell growth and chemosensitivity. Br. J. Cancer 1987, 56, 279–285. [Google Scholar] [CrossRef]
- Okaiyeto, K.; Ojemaye, M.O.; Hoppe, H.; Mabinya, L.V.; Okoh, A.I. Phytofabrication of silver/silver chloride nanoparticles using aqueous leaf extract of Oedera genistifolia: Characterization and antibacterial potential. Molecules 2019, 24, 4382. [Google Scholar] [CrossRef] [Green Version]
- Rampa, K.M.; Van De Venter, M.; Koekemoer, T.C.; Swanepoel, B.; Venables, L.; Hattingh, A.C.; Viljoen, A.M.; Kamatou, G.P. Exploring four south African croton species for potential anti-inflammatory properties: In vitro activity and toxicity risk as-sessment. J. Ethnopharmacol. 2022, 282, 114596. [Google Scholar] [CrossRef]
- Stander, M.A.; Van Wyk, B.-E.; Taylor, M.J.C.; Long, H.S. Analysis of Phenolic Compounds in Rooibos Tea (Aspalathus linearis) with a Comparison of Flavonoid-Based Compounds in Natural Populations of Plants from Different Regions. J. Agric. Food Chem. 2017, 65, 10270–10281. [Google Scholar] [CrossRef]
- Kind, T.; Tsugawa, H.; Cajka, T.; Ma, Y.; Lai, Z.; Mehta, S.S.; Wohlgemuth, G.; Barupal, D.K.; Showalter, M.R.; Arita, M.; et al. Identification of small molecules using accurate mass MS/MS search. Mass Spectrom. Rev. 2018, 37, 513–532. [Google Scholar] [CrossRef]














{kind=link}
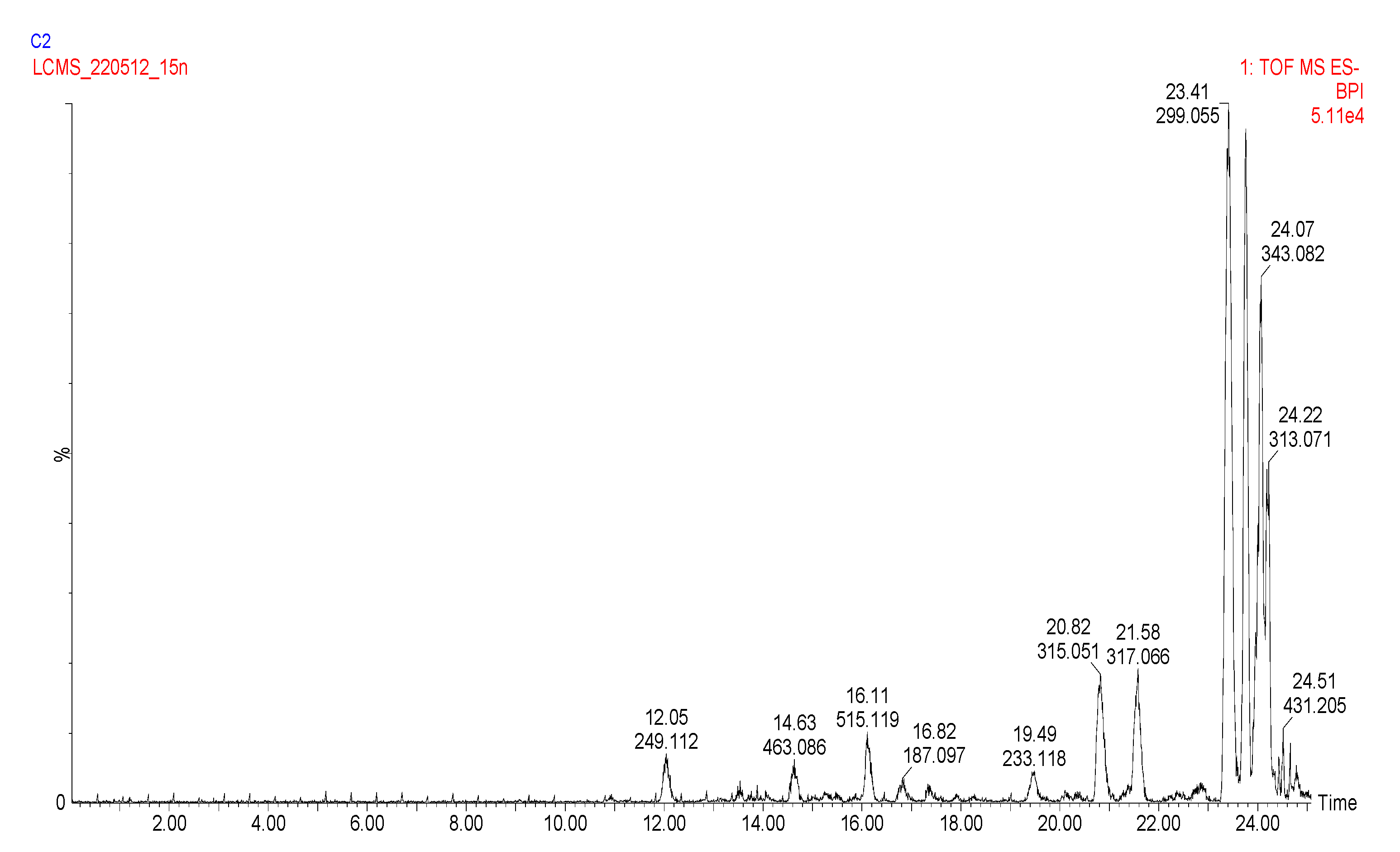{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts | Polyphenols (mg GAE/g) | Flavonols (mg QE/g) | FRAP (µmol AAE/g) | DPPH (µmol TE/g) | TEAC (µmol TE/g) |
|---|---|---|---|---|---|
| Methanolic extract | 112 ± 2.81 | 76.12 ± 7.95 | 752.64 ± 89.0 | 812.18 ± 51.12 | 631.63 ± 17.42 |
| Chloroform extract | 39.93 ± 1.36 | 44.81 ± 3.74 | 107.10 ± 2.41 | 58.70 ± 5.18 | 118.63 ± 3.74 |
| No | tR (min) | UV λmax (nm) | m/z [M − H]− | MS/MS | Tentative Name | Identification | |
|---|---|---|---|---|---|---|---|
| 1 | 1.13 | 191.0549 | 119 | p-Coumaric acid ethyl ester | C1 | New | |
| 2 | 4.14 | 295 | 554.2621 Cl− | 355, 401, 113 | Caffeoyl-hexuronide derivative | C1 | [27] |
| 3 | 7.11 | 243, 326 | 353.0859 | 191, 179 | 3-Caffeoylquinic acid (Neochlorogenic acid) | C1 | Standard |
| 4 | 9.10 | 242, 326 sh | 353.0879 | 191 | 1-Caffeoylquinic acid/ Chlorogenic acid | C1 | [27] |
| 5 | 10.45 | 249, 318 | 305.0705 | 198 | Epigallocatechin | C1 | New |
| 6 | 11.57 | 237, 270, 330 | 593.1514 | 171, 479, 353 | Apigenin-6,8-di-C-glycoside | C1 | New |
| 7 | 12.05 | 249.1122 | 205, 231, 187 | Gemfibrozil | C2 | New | |
| 8 | 12.14 | 245, 330 | 367.1002 | 191, 173, 134 | Feruloyl quinic acid | C1 | New |
| 9 | 14.60 | 281, 284 | 463.0887 | 287, 151, 135, 113 | Eriodictyol-O-hexuronide | C1, C2 | [27] |
| 10 | 14.66 | 284 sh | 463.0890 | 287 | Eriodictyol-O-hexuronide | C1 | [27] |
| 11 | 15.00 | 255, 347 | 461.0755 | 285, 217, 175 | Kaempferol-7-O-glucuronide | C1 | New |
| 12 | 16.11 | 245, 327 sh | 515.1200 | 173 > 179, 191, 353 | 1,5-Dicaffeoylquinic acid | C1, C2 | New |
| 13 | 16.50 | 243, 320 | 515.1211 | 353 | 3,4-Dicaffeoylquinic acid | C2 | [27] |
| 14 | 16.82 | 447.0944 | 187 | 6-Hydroxyluteolin 7-O-rhamnoside | C2 | New | |
| 15 | 17.21 | 243, 326 | 515.1209 | 353 | 3,5-Dicaffeoylquinic acid | C1 | [27] |
| 16 | 17.42 | 245, 323 sh | 515.1176 | 173 > 179, 135, 191, 353 | 1,4-Dicaffeoylquinic acid | C1 | [27] |
| 17 | 18.20 | 288, 325 | 477.1053 | 301, 171, 145 | Quercetin 31-O-glucuronide | C1 | New |
| 18 | 19.40 | 288 | 287.0539 | 151, 135 | Eriodictyol | C1 | [27] |
| 19 | 19.48 | 234, 289 | 287.0551 | 151, 135 | Eriodictyol | C1 | [27] |
| 20 | 19.49 | 288 | 233.1181 | 189, 215, 221, 159, 203, 177 | unknown | C2 | New |
| 21 | 20.20 | 251, 347 | 523.2188 | 285 | Luteolin derivative | C1 | New |
| 22 | 20.30 | 255, 338 | 285.0404 | 187, 195 | Luteolin | C1 | [45] |
| 23 | 20.76 | 250, 347 | 315.0484 | 300 | Isorhamnetin | C1 | New |
| 24 | 20.82 | 252, 273, 346 | 315.0503 | 161, 300 | Isorhamnetin | C2 | New |
| 25 | 21.58 | 236, 288 | 317.0667 | 289, 299, 195 | Myricetin | C2 | New |
| 26 | 22.40 | 289 | 271.0585 | 227 | Narigenin | C1 | New |
| 27 | 22.86 | 265, 337 | 269.0450 | 161, 151, 117 | Apigenin | C1 | [45] |
| 28 | 23.40 | 237, 272, 333 | 299.0551 | 284 | Chrysoeriol (3′-O-methylluteolin) | C1, C2 | New |
| 29 | 23.71 | 242, 275, 340 | 329.0661 | 315 | Tricin | C1 | New |
| 30 | 24.07 | 247, 273, 339 | 343.0826 | 315, 299, 297, 227 | 3′-O-methyl tricin/3″,4″,5,7-tetramethoxyluteolin (methlut) | C1, C2 | New |
| 31 | 24.22 | 235, 275, 329 | 313.0720 | 253 | 31,41-dimethyluteolin | C2 | New |
| 32 | 24.40 | 255 | 445.2216 | 325, 339, 265, 311, 287, 150, 221 | Apigenin-7-O-glucuronide | C1 | New |
| 33 | 24.51 | 252 | 431.2066 | 265, 150 | Apigenin-6-C-hexose | C1, C2 | New |
| No | tR (min) | UV λmax (nm) | m/z [M + H]+ | MS/MS | Assignment | Identification | |
|---|---|---|---|---|---|---|---|
| 34 | 0.71 | 217.069 | 203 | Xanthotoxin | C1 | New | |
| 35 | 2.70 | 163.0404 | 135, 145 | Hydroxy coumarin | New | ||
| 36 | 2.88 | 236, 326 | 188.0727 | 146 | Trans-3-indoleacrylic acid | C1 | New |
| 37 | 3.01 | 242, 286, 304, 340 | 163.040 | 145, 135 | Hydroxy coumarin | C1 | New |
| 38 | 3.33 | 271, 331 | 247.133 | 229, 183, 201 | Isopimpinellin | C2 | New |
| 39 | 3.45 | 321 sh | 380.208 | 163, 364 | Xanthotoxin-Hydroxy coumarin adduct | C1 | New |
| 40 | 3.72 | 291, 323 | 263.130 | 217, 187, 203, 91, 245 | Leptophyllin | C2 | New |
| 41 | 4.40 | 286, 327 | 422.219 | 211, 163 | Unknown dimer | C1 | New |
| 42 | 4.62 | 312 | 247.134 | 217, 229, 183, 171 | Isopimpinellin | C2 | New |
| 43 | 4.72 | 291, 321 | 247.135 | 217, 229 | Isopimpinellin | C2 | New |
| 44 | 4.84 | 312, 323 | 217.124 | 171, 143 | Xanthotoxin | C2 | New |
| 45 | 5.00 | 287 | 289.0723 | 229, 163 | Dihydrokaempferol | C1 | New |
| 46 | 5.05 | 250, 293, 346 | 287.0566 | 229 | Kaempferol | C1 | New |
| 47 | 5.12 | 250, 293, 346 | 317.068 | 302, 163 | Isorhamnetin | C1 | New |
| 48 | 5.18 | 273, 346 | 406.223 | 317 | Isorhamnetin-4″-O-malonyl | C1 | New |
| 49 | 5.60 | 268, 291, 331 | 271.0615 | 153, 163 | 3″,4″,7-trihydroxy flavanone | New | |
| 50 | 5.64 | 274, 330 | 301.0721 | 247, 229 | Methoxy kaempferol | C1, C2 | New |
| 51 | 5.78 | 273, 346 | 331.08185 | 316, 301, 168 | Tricin | C1, C2 | New |
| 52 | 5.90 | 271, 347 | 324.181 Na | 229, 247, 307, 183, 289, 247 | Methoxy kaempferol | C1, C2 | New |
| 53 | 6.20 | 235 289, 329 | 289.1440 | 229, 249, 185 | Dihydrokaempferol | C1, C2 | New |
| 54 | 6.27 | 287, 325 | 231.1384 | 185, 157 | 6-(3,3-Dimethylallyl)-7-hydroxycoumarin | C1, C2 | New |
| 55 | 6.50 | 274, 346 | 345.0994 | 229, 247 | 3″,4″,5,7-tetramethoxyluteolin (methlut) | C1, C2 | New |
| 56 | 7.02 | 275, 334 | 315.087 | 251 | 31,41-dimethyluteolin | C1, C2 | New |
| 57 | 7.20 | 244, 276, 335 | 359.1140 | 217, 235 | Xanthotoxin derivative | C1, C2 | New |
| 58 | 7.76 | 241, 331 | 231.1385 | 185, 157, 213 | 6-(3,3-Dimethylallyl)-7-hydroxycoumarin | C1, C2 | New |
| 59 | 8.43 | 244, 329, 331 | 233.1543 | 187, 215 | 6-(3,3-Dimethyl)-7-hydroxycoumarin | C1, C2 | New |
| 60 | 9.06 | 247, 331 | 231.1385 | 185, 157 | 6-(3,3-Dimethylallyl)-7-hydroxycoumarin | C1 | New |
| 61 | 9.07 | 312, 335 | 331.1905 | 231, 175, 185, 213 | 6-(3,3-Dimethylallyl)-7-hydroxycoumarinderivative | C1, C2 | New |
| 62 | 9.55 | 242, 274 | 149.0248 231.1385 | 93 | 6-(3,3-Dimethylallyl)-7-hydroxycoumarin | C1, C2 | New |
| No | Name | tR (min) | UVλmax (nm) | Regression Equation | Linear Range mg/L | R2 | LOD * mg/L | LOQ * mg/L |
|---|---|---|---|---|---|---|---|---|
| 1 | Neochlorogenic acid | 7.1 | 300, 325 | y = 3.0x + 7.9 | 3.9–31.3 | 0.994 | 10.0 | 30.4 |
| 2 | Catechin | 8.7 | 278 | y = 1.6x + 94.2 | 3.9–31.3 | 0.999 | 5.3 | 16.0 |
| 3 | Caffeic acid | 9.4 | 300, 325 | y =1.9x − 1497.4 | 3.9–15.6 | 0.993 | 6.2 | 18.9 |
| 4 | Epicatechin | 10.8 | 277 | y = 1.2x + 1859.3 | 3.9–31.3 | 0.996 | 4.0 | 12.2 |
| 5 | p-Coumaric acid | 11.8 | 300, 308 | y = 3.9x + 2855.3 | 7.8–62.5 | 0.993 | 12.9 | 39.1 |
| 6 | Ferulic acid | 13.4 | 300, 323 | y = 4.2x + 1098.7 | 7.8–31.3 | 1.000 | 13.8 | 41.7 |
| 7 | Rutin | 14.5 | 254, 255, 354 | y = 1.9x + 449.7.3 | 7.8–31.3 | 0.994 | 6.1 | 18.6 |
| 8 | Phloridzin | 17.9 | 284 | y = 1.4x + 1603.3 | 3.9–15.6 | 1.000 | 4.7 | 14.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okaiyeto, K.; Kerebba, N.; Oguntibeju, O.O. UPLC-ESI-QTOF-MS Profiling of Phenolic Compounds from Eriocephalus africanus: In Vitro Antioxidant, Antidiabetic, and Anti-Inflammatory Potentials. Molecules 2022, 27, 8912. https://doi.org/10.3390/molecules27248912
Okaiyeto K, Kerebba N, Oguntibeju OO. UPLC-ESI-QTOF-MS Profiling of Phenolic Compounds from Eriocephalus africanus: In Vitro Antioxidant, Antidiabetic, and Anti-Inflammatory Potentials. Molecules. 2022; 27(24):8912. https://doi.org/10.3390/molecules27248912
Chicago/Turabian StyleOkaiyeto, Kunle, Nasifu Kerebba, and Oluwafemi O. Oguntibeju. 2022. "UPLC-ESI-QTOF-MS Profiling of Phenolic Compounds from Eriocephalus africanus: In Vitro Antioxidant, Antidiabetic, and Anti-Inflammatory Potentials" Molecules 27, no. 24: 8912. https://doi.org/10.3390/molecules27248912
APA StyleOkaiyeto, K., Kerebba, N., & Oguntibeju, O. O. (2022). UPLC-ESI-QTOF-MS Profiling of Phenolic Compounds from Eriocephalus africanus: In Vitro Antioxidant, Antidiabetic, and Anti-Inflammatory Potentials. Molecules, 27(24), 8912. https://doi.org/10.3390/molecules27248912