

Effect of Common Cooking and Drying Methods on Phytochemical and Antioxidant Properties of Corchorus olitorius Identified Using Liquid Chromatography-Mass Spectrometry (LC-MS)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Preparation
2.3. Solvent Extraction
2.3.1. Determination of Total Phenolic Content
2.3.2. Determination of Total Flavonoid Content
2.4. Determination of Antioxidant Activity
2.4.1. ABTS Radical Scavenging Activity Assay
2.4.2. DPPH Radical Scavenging Activity Assay
2.4.3. Ferric Reducing Antioxidant Power Assay (FRAP)
2.5. Fourier Transform Infrared Spectroscopy (FTIR)
2.6. Extraction of Metabolites
2.7. Analysis Using Liquid Chromatography-Mass Spectrometry (LC-MS)
2.8. Statistical Analysis
3. Results and Discussion
3.1. Effect of Common Cooking and Drying Methods on Total Phenolic and Flavonoid Content
3.2. Antioxidant Activity
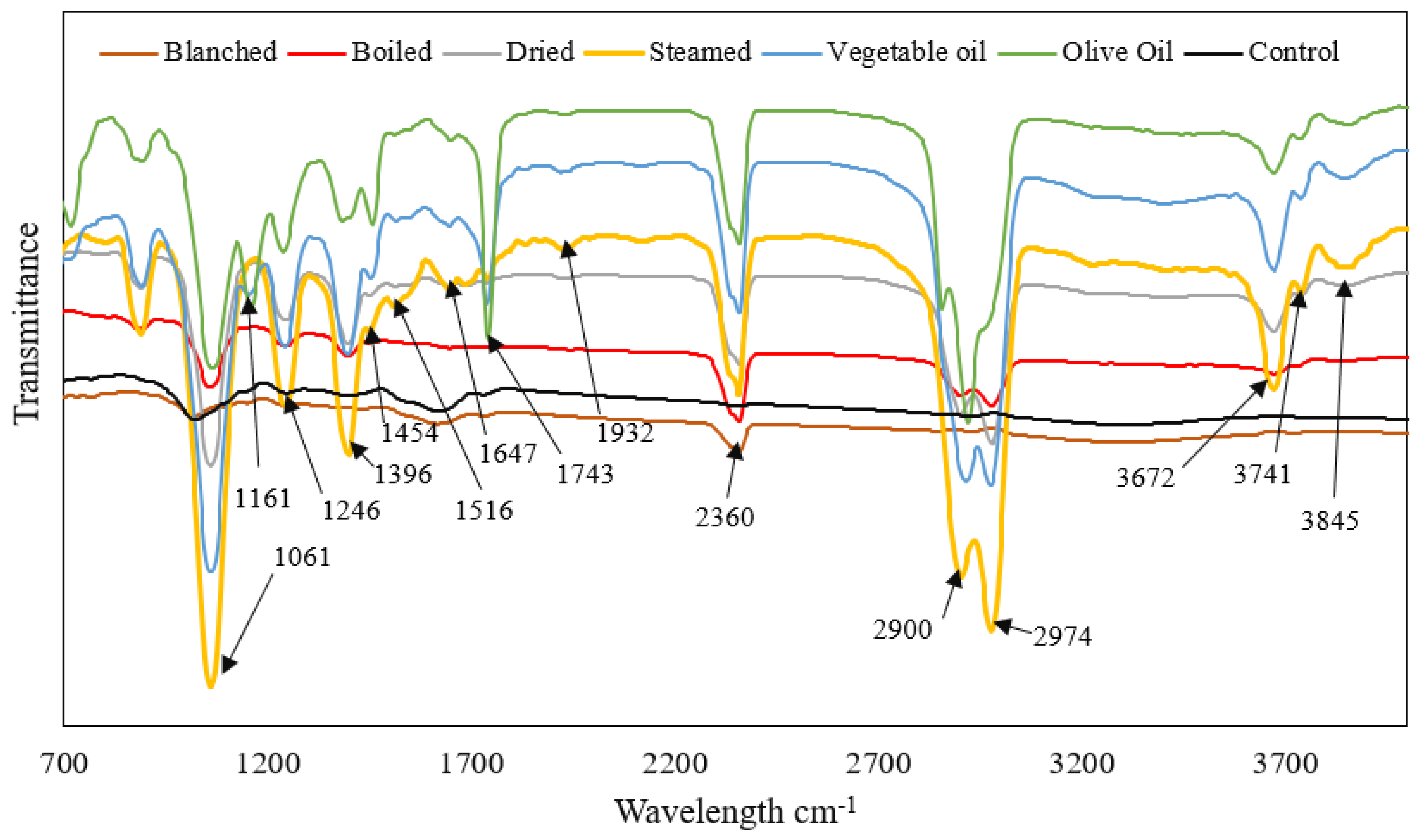
3.3. Fourier Transform Infrared Spectroscopy (FTIR)
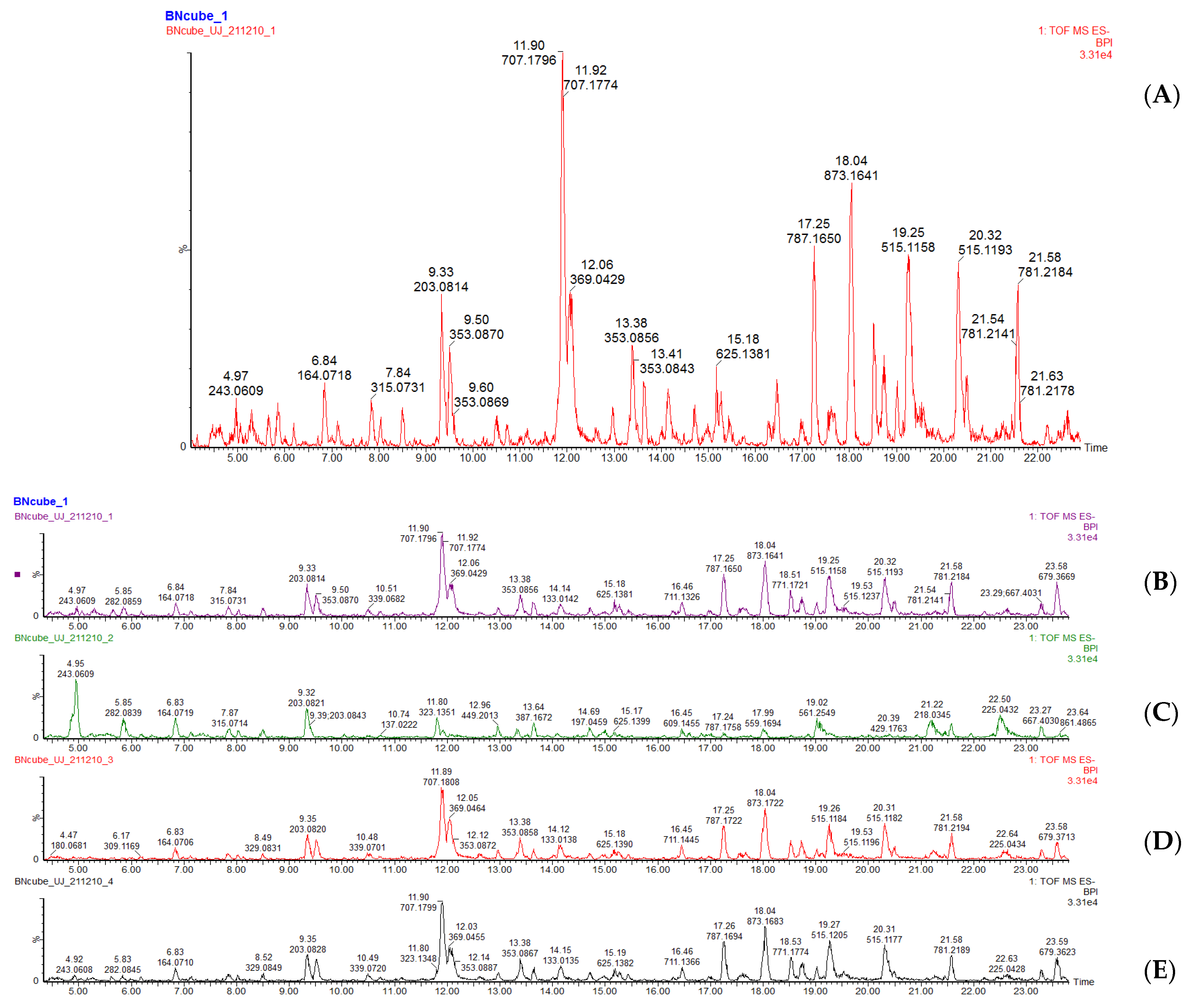
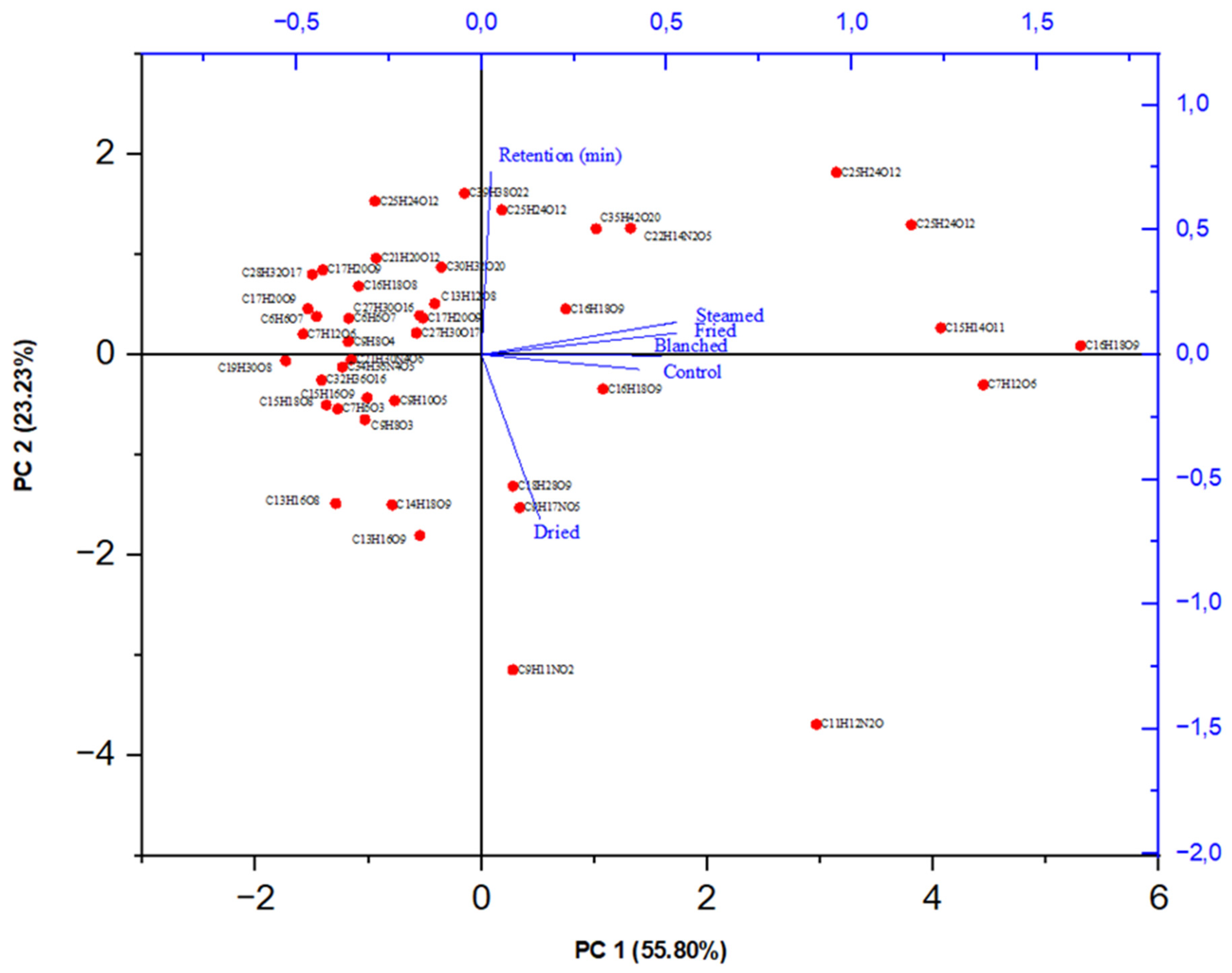
3.4. Quantification and Qualification of Metabolites Using LC-MS
3.4.1. Characterization of Amino Acids
3.4.2. Characterization of Phenolic Acids
3.4.3. Characterization of Flavonoid Derivatives
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Traoré, K.; Parkouda, C.; Guissou, A.W.B.D.; Traoré, Y.; Savadogo, A. Effect of Cooking Methods on the β-carotene Content of Jute Mallow Leaves (Corchorus olitorius). Am. J. Food Sci. Technol. 2019, 7, 223–226. [Google Scholar]
- Uusiku, N.P.; Oelofse, A.; Duodu, K.G.; Bester, M.J.; Faber, M. Nutritional value of leafy vegetables of sub-Saharan Africa and their potential contribution to human health: A review. J. Food Compos. Anal. 2010, 23, 499–509. [Google Scholar] [CrossRef]
- Maseko, I.; Mabhaudhi, T.; Tesfay, S.; Araya, H.; Fezzehazion, M.; Plooy, C. African Leafy Vegetables: A Review of Status, Production and Utilization in South Africa. Sustainability 2017, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Moyo, S.M.; Mavumengwana, V.; Kayitesi, E. Effects of cooking and drying on phenolic compounds and antioxidant activity of African green leafy vegetables. Food Rev. Int. 2018, 34, 248–264. [Google Scholar] [CrossRef]
- Kochanek, K.D.; Murphy, S.L.; Xu, J. Deaths: Final Data for 2011. Natl. Vital Stat. Rep. 2015, 63, 1–120. [Google Scholar]
- Cencic, A.; Chingwaru, W. The Role of Functional Foods, Nutraceuticals, and Food Supplements in Intestinal Health. Nutrients 2010, 2, 611–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lima, A.C.S.; da Rocha Viana, J.D.; de Sousa Sabino, L.B.; da Silva, L.M.R.; da Silva, N.K.V.; de Sousa, P.H.M. Processing of three different cooking methods of cassava: Effects on in vitro bioaccessibility of phenolic compounds and antioxidant activity. LWT—Food Sci. Technol. 2017, 76, 253–258. [Google Scholar] [CrossRef]
- Van Rensburg, W.S.J.; Van Averbeke, W.; Slabbert, R.; Faber, M.; Van Jaarsveld, P.; Van Heerden, I.; Wenhold, F.; Oelofse, A. African leafy vegetables in South Africa. Water SA 2007, 33, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Idirs, S.; Yisa, J.; Ndamitso, M. Nutritional composition of Corchorus olitorius leaves. Anim. Prod. Res. Adv. 2009, 5, 49827. [Google Scholar] [CrossRef]
- Moyo, M.; Amoo, S.O.; Ncube, B.; Ndhlala, A.R.; Finnie, J.F.; van Staden, J. Phytochemical and antioxidant properties of unconventional leafy vegetables consumed in southern Africa. South Afr. J. Bot. 2013, 84, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.M. Biochemistry, Medicinal and Food values of Jute (Corchorus capsularis L. and C. olitorius L.) leaf: A Review. Int. J. Enhanc. Res. Sci. Technol. Eng. 2013, 2, 35–44. [Google Scholar]
- Adebo, H.O.; Ahoton, L.E.; Quenum, F.J.B.; Adoukonou-Sagbadja, H.; Bello, D.O.; Chrysostome, C.A.A.M. Ethnobotanical Knowledge of Jute (Corchorus olitorius L.) in Benin. Eur. J. Med. Plants 2018, 26, 1–11. [Google Scholar] [CrossRef]
- Onwordi, C.T.; Ogungbade, A.M.; Wusu, A.D. The Proximate and Mineral Composition of Three Leafy Vegetables Commonly Consumed in Lagos, Nigeria. Afr. J. Pure Appl. Chem. 2009, 3, 102–107. [Google Scholar]
- Ndlovu, J.; Afolayan, A.J. Nutritional Analysis of the South African Wild Vegetable Corchorus olitorius L. Asian J. Plant Sci. 2008, 7, 615–618. [Google Scholar] [CrossRef]
- Dansi, A.; Adjatin, A.; Adoukonou-Sagbadja, H.; Faladé, V.; Yedomonhan, H.; Odou, D.; Dossou, B. Traditional leafy vegetables and their use in the Benin Republic. Genet. Resour. Crop Evol. 2008, 55, 1239–1256. [Google Scholar] [CrossRef]
- Youssef, M.; Mokhtar, S.; Morsy, N. Effect of Hot Air Drying Variables on Phytochemicals and Antioxidant Capacityof Jew’s Mallow (Corchorus olitorius L.) Leaves. Suez Canal Univ. J. Food Sci. 2014, 2, 1–8. [Google Scholar] [CrossRef]
- Lewu, M.; Adebola, P.O.; Afolayan, A.J. Effect of cooking on the proximate composition of the leaves of some accessions of Colocasia esculenta (L.) Schott in KwaZulu-Natal province of South Africa. Afr. J. Biotechnol. 2009, 8, 1619–1622. [Google Scholar]
- Musa, A.; Ogbadoyi, E.O. Effect of Cooking and Sun Drying on Micronutrients, Antinutrients an Toxic Substances in Corchorus olitorius (Jute Mallow). J. Nutr. Food Sci. 2012, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Adefegha, S.A.; Oboh, G. Enhancement of total phenolics and antioxidant properties of some tropical green leafy vegetables by steam cooking. J. Food Process. Preserv. 2011, 35, 615–622. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Al-Farsi, M.A.; Lee, C.Y. Optimization of phenolics and dietary fibre extraction from date seeds. Food Chem. 2008, 108, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Awika, J.M.; Rooney, L.W.; Wu, X.; Prior, R.L.; Cisneros-Zevallos, L. Screening Methods to Measure Antioxidant Activity of Sorghum (Sorghum bicolor) and Sorghum Products. J. Agric. Food Chem. 2003, 51, 6657–6662. [Google Scholar] [CrossRef] [PubMed]
- Awwad, A.M.; Salem, N.M.; Abdeen, A.O. Green synthesis of silver nanoparticles using carob leaf extract and its antibacterial activity. Int. J. Ind. Chem. 2013, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Adedapo, A.A.; Jimoh, F.O.; Afolayan, A.J.; Masika, P.J. Antioxidant Properties of the Methanol Extracts of the Leaves and Stems of Celtis africana. Rec. Nat. Prod. 2009, 3, 23–31. [Google Scholar]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.M.; Arráez-Román, D.; Segura-Carretero, A. Comprehensive metabolite profiling of Arum palaestinum (Araceae) leaves by using liquid chromatography–tandem mass spectrometry. Food Res. Int. 2015, 70, 74–86. [Google Scholar] [CrossRef]
- Ramabulana, T.; Mavunda, R.D.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A.; Madala, N.E. Perturbation of pharmacologically relevant polyphenolic compounds in Moringa oleifera against photo-oxidative damages imposed by gamma radiation. J. Photochem. Photobiol. B Biol. 2016, 156, 79–86. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; Vander Gheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef]
- Andabati, B.; Muyonga, J. Phenolic content and antioxidant activity of selected Ugandan traditional medicinal foods. Afr. J. Food Sci. 2014, 8, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Thi, N.D.; Hwang, E.S. Effects of drying methods on contents of bioactive compounds and antioxidant activities of black chokeberries (Aronia melanocarpa). Food Sci. Biotechnol. 2016, 25, 55–61. [Google Scholar] [CrossRef]
- Provost, J.J.; Colabroy, K.L.; Kelly, B.S.; Wallert, M.A. The Science of Cooking: Understanding the Biology and Chemistry Behind Food and Cooking; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 5–20. [Google Scholar]
- Craft, B.D.; Kerrihard, A.L.; Amarowicz, R.; Pegg, R.B. Phenol-Based Antioxidants and the In Vitro Methods Used for Their Assessment. Compr. Rev. Food Sci. Food Saf. 2012, 11, 148–173. [Google Scholar] [CrossRef]
- Mazzeo, T.; N’Dri, D.; Chiavaro, E.; Visconti, A.; Fogliano, V.; Pellegrini, N. Effect of two cooking procedures on phytochemical compounds, total antioxidant capacity and colour of selected frozen vegetables. Food Chem. 2011, 128, 627–633. [Google Scholar] [CrossRef]
- Kunyanga, C.N.; Imungi, J.K.; Okoth, M.W.; Biesalski, H.K.; Vadivel, V. Total phenolic content, antioxidant and antidiabetic properties of methanolic extract of raw and traditionally processed Kenyan indigenous food ingredients. LWT—Food Sci. Technol. 2012, 45, 269–276. [Google Scholar] [CrossRef]
- Miglio, C.; Chiavaro, E.; Visconti, A.; Fogliano, V.; Pellegrini, N. Effects of Different Cooking Methods on Nutritional and Physicochemical Characteristics of Selected Vegetables. J. Agric. Food Chem. 2008, 56, 139–147. [Google Scholar] [CrossRef]
- Tuck, K.L.; Hayball, P.J. Major phenolic compounds in olive oil: Metabolism and health effects. J. Nutr. Biochem. 2002, 13, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Coates, J. Interpretation of Infrared Spectra, A Practical Approach. In Encyclopedia of Analytical Chemistry; Meyers, R.A., Ed.; John Wiley & Sons Ltd.: Chichester, UK, 2000; pp. 10881–10882. [Google Scholar]
- Pop, R.M.; Buzoianu, A.D.; Rați, I.V.; Socaciu, C. Untargeted Metabolomics for Sea Buckthorn (Hippophae rhamnoides ssp. carpatica) Berries and Leaves: Fourier Transform Infrared Spectroscopy as a Rapid Approach for Evaluation and Discrimination. Not. Bot. Horti Agrobot. Cluj Napoca 2014, 42, 545–550. [Google Scholar] [CrossRef]
- Pop, C.; Apostu, S.; Rotar, A.M.; Semeniuc, C.A.; Sindic, M.; Mabon, N. FTIR spectroscopic characterization of a new biofilm obtained from kefiran. J. Agroaliment. Process. Technol. 2013, 19, 157–159. [Google Scholar]
- Ito, H.; Kikuzaki, H.; Ueno, H. Effects of Cooking Methods on Free Amino Acid Contents in Vegetables. J. Nutr. Sci. Vitaminol. 2019, 65, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Richard, D.M.; Dawes, M.A.; Mathias, C.W.; Acheson, A.; Hill-Kapturczak, N.; Dougherty, D.M. L-Tryptophan: Basic Metabolic Functions, Behavioral Research and Therapeutic Indications. Int. J. Tryptophan Res. IJTR 2009, 2, 45–60. [Google Scholar] [CrossRef] [Green Version]
- Akram, M.; Munir, N.; Daniyal, M.; Egbuna, C.; Găman, M.A.; Onyekere, P.F.; Olatunde, A. Vitamins and Minerals: Types, Sources and their Functions. In Functional Foods and Nutraceuticals, 1st ed.; Egbuna, C., Dable Tupas, G., Eds.; Springer: Cham, Switzerland, 2020; pp. 149–172. [Google Scholar]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.M.; Kumar, C.S. Syringic acid (SA)—A Review of Its Occurrence, Biosynthesis, Pharmacological and Industrial Importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Khoza, B.S.; Gbashi, S.; Steenkamp, P.A.; Njobeh, P.B.; Madala, N.E. Identification of hydroxylcinnamoyl tartaric acid esters in Bidens pilosa by UPLC-tandem mass spectrometry. S. Afr. J. Bot. 2016, 103, 95–100. [Google Scholar] [CrossRef]
- Azuma, K.; Nakayama, M.; Koshioka, M.; Ippoushi, K.; Yamaguchi, Y.; Kohata, K.; Yamauchi, Y.; Ito, H.; Higashio, H. Phenolic Antioxidants from the Leaves of Corchorus olitorius L. J. Agric. Food Chem. 1999, 47, 3963–3966. [Google Scholar] [CrossRef]
- Yakoub, A.R.B.; Abdehedi, O.; Jridi, M.; Elfalleh, W.; Nasri, M.; Ferchichi, A. Flavonoids, phenols, antioxidant, and antimicrobial activities in various extracts from Tossa jute leave (Corchorus olitorus L.). Ind. Crops Prod. 2018, 118, 206–213. [Google Scholar] [CrossRef]
- Guzzetti, L.; Panzeri, D.; Ulaszewska, M.; Sacco, G.; Forcella, M.; Fusi, P.; Tommasi, N.; Fiorini, A.; Campone, L.; Labra, M. Assessment of Dietary Bioactive Phenolic Compounds and Agricultural Sustainability of an African Leafy Vegetable Corchorus olitorius L. Front. Nutr. 2021, 8, 667812. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Anaya, J.; Samaniego-Sánchez, C.; Castañeda-Saucedo, M.C.; Villalón-Mir, M.; de la Serrana, H.L. Phenols and the antioxidant capacity of Mediterranean vegetables prepared with extra virgin olive oil using different domestic cooking techniques. Food Chem. 2015, 188, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Nicoli, M.C.; Anese, M.; Parpinel, M. Influence of processing on the antioxidant properties of fruit and vegetables. Trends Food Sci. Technol. 1999, 10, 94–100. [Google Scholar] [CrossRef]
- Mediani, A.; Abas, F.; Khatib, A.; Tan, C.P. Cosmos caudatus as a potential source of polyphenolic compounds: Optimisation of oven drying conditions and characterisation of its functional properties. Molecules 2013, 18, 10452–10464. [Google Scholar] [CrossRef]
- Birková, A.; Hubková, B.; Bolerázska, B.; Mareková, M.; Čižmárová, B. Caffeic acid: A brief overview of its presence, metabolism, and bioactivity. Bioact. Compd. Health Dis. 2020, 3, 74–81. [Google Scholar] [CrossRef]
- Iwashina, T.; Matsumoto, S. Flavonoid Glycosides from the Fern, Schizaea (Schizaeaceae) in South Pacific Region, and their Distribution Pattern. Bull. Natl. Mus. Nat. Sci. Ser. B 2013, 39, 195–201. [Google Scholar]
- Simões, A.N.; Moreira, S.I.; Mosquim, P.R.; Soares, N.F.F.; Puschmann, R. The effects of storage temperature on the quality and phenolic metabolism of whole and minimally processed kale leaves. Acta Sci. Agron. 2014, 37, 101. [Google Scholar] [CrossRef] [Green Version]
- Medoua, G.N.; Oldewage-Theron, W.H. Bioactive compounds and antioxidant properties of selected fruits and vegetables available in the Vaal region, South Africa. J. Food Biochem. 2011, 35, 1424–1433. [Google Scholar] [CrossRef]
- Singla, R.K.; Dhir, V.; Madaan, R.; Kumar, D.; Singh Bola, S.; Bansal, M.; Kumar, S.; Dubey, A.K.; Singla, S.; Shen, B. The Genus Alternanthera: Phytochemical and Ethnopharmacological Perspectives. Front. Pharmacol. 2022, 13, 769111. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Tao, X.; Zhang, C.; Lu, Y.; Wei, D. Protective effects of hyperoside (quercetin-3-o-galactoside) to PC12 cells against cytotoxicity induced by hydrogen peroxide and tert-butyl hydroperoxide. Biomed. Pharmacother. 2005, 59, 481–490. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample Type | TPC (mg GAE/g) | TFC (mg QE//g) |
|---|---|---|
| Control | 47.40 a ± 1.02 | 25.50 a ± 0.86 |
| Blanched | 88.35 c ± 1.76 | 36.65 c ± 3.33 |
| Boiled | 52.93 a ± 5.79 | 39.47 c ± 1.18 |
| Dried | 99.02 d ± 2.37 | 44.57 d ± 4.29 |
| Fried Olive Oil | 106.86 e ± 3.66 | 31.75 b ± 1.37 |
| Fried Vegetable Oil | 80.64 b ± 1.62 | 55.31 e ± 1.20 |
| Steamed | 118.89 f ± 3.32 | 78.42 f ± 2.59 |
| Sample Type | DPPH (µM TE/g) | FRAP (µM TE/g) | ABTS (µM TE/g) |
|---|---|---|---|
| Control | 66.46 b ± 0.30 | 139.24 b ± 10.12 | 75.49 a ± 2.89 |
| Blanched | 82.28 d ± 0.35 | 146.61 d ± 2.24 | 95.30 c ± 6.67 |
| Boiled | 56.70 a ± 0.66 | 72.62 a ± 0.17 | 107.29 d ± 0.58 |
| Dried | 87.77 e ± 0.47 | 151.69 e ± 5.77 | 111.20 e ± 0.73 |
| Fried Olive Oil | 99.12 f ± 1.40 | 159.96 f ± 6.29 | 119.45 f ± 3.00 |
| Fried Vegetable Oil | 70.24 c ± 10.12 | 143.22 c ± 7.44 | 81.89 b ± 5.72 |
| Steamed | 119.58 g ± 4.47 | 167.31 g ± 9.23 | 122.23 g ± 5.77 |
| Molecule No. | Metabolite Name | Retention Time/min | Average m/z | Molecular Formula | Control | Blanched (mg/kg) | Dried (mg/kg) | Fried (mg/kg) | Steamed (mg/kg) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | L-Phenylalanine | 6.8 | 164 | C9H11NO2 | 110.6 | 120.9 | 184.9 | 102.6 | 115.0 |
| 2 | Pseudolaroside A | 7.1 | 299 | C13H16O8 | 52.8 | 42.7 | 47.3 | 29.7 | 37.1 |
| 3 | Gentesic acid 5-O-glucoside | 7.9 | 315 | C13H16O9 | 110.3 | 85.7 | 86.4 | 57.1 | 60.7 |
| 4 | Pantothenic acid | 8.0 | 218 | C9H17NO5 | 25.9 | 50.5 | 67.0 | 42.8 | 47.0 |
| 5 | 1-O-vanilloyl-beta-D-glucose | 8.5 | 329 | C14H18O9 | 80.4 | 69.7 | 72.5 | 51.1 | 62.4 |
| 6 | L-Tryptophan | 9.3 | 203 | C11H12N2O | 285.3 | 264.1 | 278.1 | 232.3 | 251.5 |
| 7 | Neochlorogenic acid | 9.5 | 353 | C16H18O9 | 173.0 | 194.6 | 9.6 | 189.8 | 209.8 |
| 8 | Aesculin | 10.5 | 339 | C15H16O9 | 79.7 | 55.7 | 17.1 | 47.7 | 61.4 |
| 9 | Kaempferide 3-rhamnoside-7-(6”-succinylglucoside) | 10.5 | 707 | C32H36O16 | 60.7 | 30.3 | 0.4 | 43.5 | 22.1 |
| 10 | 4-Hydroxybenzoic acid | 10.7 | 137 | C7H6O3 | 71.6 | 43.8 | 26.1 | 27.6 | 30.8 |
| 11 | 1-O-p-Coumaroyl-beta-D-glucose | 11.8 | 325 | C15H18O8 | 48.9 | 29.0 | 41.3 | 29.4 | 29.3 |
| 12 | 4-Hydroxycinnamic acid | 11.8 | 163 | C9H8O3 | 60.5 | 49.7 | 55.3 | 48.4 | 52.2 |
| 13 | Quinic acid | 11.9 | 191 | C7H12O6 | 338.9 | 405.9 | 63.7 | 393.2 | 416.2 |
| 14 | Crypto-chlorogenic acid | 11.9 | 353 | C16H18O9 | 65.2 | 601.9 | 68.8 | 563.6 | 580.8 |
| 15 | 2-O-Caffeoylhy- droxycitric acid | 12.0 | 69 | C15H14O11 | 450.8 | 307.1 | 5.9 | 402.9 | 315.9 |
| 16 | Caffeic acid | 12.6 | 179 | C9H8O4 | 71.7 | 35.6 | 4.1 | 57.1 | 42.9 |
| 17 | 5-Hydroxy-6-methoxycoumaric acid | 13.0 | 369 | C7H12O6 | 32.0 | 26.1 | 3.2 | 27.4 | 27.4 |
| 18 | Chlorogenic acid | 13.4 | 353 | C16H18O9 | 120.2 | 196.2 | 6.7 | 185.6 | 186.4 |
| 19 | 7-Epi-12-hydroxyjasmonic acid glucoside | 13.6 | 387 | C18H28O9 | 148.5 | 132.4 | 139.6 | 82.8 | 106.6 |
| 20 | Oxalosuccinic acid | 14.0 | 189 | C6H6O7 | 28.2 | 31.7 | 6.6 | 43.3 | 35.6 |
| 21 | Citroside A | 14.1 | 431 | C19H30O8 | 20,1 | 6.0 | 40.6 | 8.2 | 11.1 |
| 22 | Caffeoylmalic acid | 14.1 | 295 | C13H12O8 | 110.7 | 81.3 | 2.2 | 102.7 | 99.0 |
| 23 | Syringic acid | 14.7 | 197 | C9H10O5 | 39.2 | 79.1 | 89.3 | 56.0 | 78.0 |
| 24 | 3-hydroxy-2-(3-methyl-2-{[3-oxo-2-(propan-2-yl)-1,2,3,4-tetrahydroquinoxaline-1-carbonyl]amino}butanamido)butanoic acid | 14.9 | 433 | C21H30N4O6 | 59.4 | 33.4 | 54.4 | 44.0 | 37.6 |
| 25 | N-(3-butoxypropyl)-2-[10-(4-methoxyphenyl)-12,14-dioxo-8,11,13-triazatetracyclo-hexadeca-1(9),2,4,6-tetraen-13-yl]benzamide | 15.0 | 579 | C34H36N4O5 | 29.9 | 37.9 | 63.6 | 41.0 | 43.9 |
| 26 | 3-O-Caffeoyl-4-O-methylquinic acid | 15.0 | 367 | C17H20O9 | 40.6 | 28.0 | 13.2 | 20.9 | 23.1 |
| 27 | Oxalosuccinic acid | 15.2 | 189 | C6H6O7 | 50.8 | 43.6 | 27.1 | 50.1 | 46.5 |
| 28 | Quercetin 3-glucosyl-(1-2)-galactoside | 15.2 | 625 | C27H30O17 | 60,1 | 100.9 | 43.8 | 75.7 | 94.9 |
| 29 | 3-O-Caffeoyl-4-O-methylquinic acid | 15.3 | 367 | C17H20O9 | 98.9 | 95.9 | 30.4 | 73.2 | 83.2 |
| 30 | 3-O-p-Coumaroylquinic acid | 15.4 | 337 | C16H18O8 | 74.0 | 49.6 | 3.9 | 45.8 | 58.0 |
| 31 | 3-O-Caffeoyl-4-O-methylquinic acid | 16.4 | 367 | C17H20O9 | 49.2 | 37.7 | 4.7 | 29.8 | 32.2 |
| 32 | Isorhamnetin 3-glucosyl-(1-2)-galactoside | 16.3 | 639 | C28H32O17 | 50.5 | 42.1 | 5.2 | 1.8 | 33.5 |
| 33 | Quercetin-O-malonylglucoside-o-glycoside | 16.5 | 711 | C30H32O20 | 72.9 | 119.5 | 13.0 | 108.1 | 105.3 |
| 34 | Quercetin 3-O-robinobioside | 16.5 | 609 | C27H30O16 | 48.6 | 100.8 | 51.8 | 76.6 | 106.2 |
| 35 | Quercetin 3-galactoside | 17.6 | 463 | C21H20O12 | 104.6 | 75.9 | 12.6 | 27.8 | 48.9 |
| 36 | Kaempferol-o-glycoside-o-sophoroside | 18.53 | 771 | C22H14N2O5 | 191.7 | 229.8 | 19.7 | 161.6 | 226.6 |
| 37 | Tricin 7-[feruloyl-(2)-glucuronyl-(1-2)-glucuronide] | 19.3 | 857 | C39H38O22 | 5.91 | 153.9 | 9.2 | 75.4 | 231.4 |
| 38 | 1,4-dicaffeoylquinic acid | 18.7 | 515 | C25H24O12 | 123.4 | 56.1 | 7.9 | 166.4 | 172.8 |
| 39 | 3,5-dicaffeoylquinic acid | 19.3 | 515 | C25H24O12 | 312.9 | 398.1 | 44.5 | 313.7 | 379.8 |
| 40 | 1,3-dicaffeoylquinic acid | 19.5 | 515 | C25H24O12 | 50.3 | 74.8 | 3.2 | 55.6 | 81.3 |
| 41 | 4,5-dicaffeoylquinic acid | 20.3 | 515 | C25H24O12 | 223.8 | 362.8 | 20.1 | 341.4 | 333.5 |
| 42 | Kaempferol 3-(2”-rhamnosyl-6”-acetylgalactoside) 7-rhamnoside | 21.6 | 781 | C35H42O20 | 59.3 | 266.1 | 75.0 | 175.3 | 208.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ncube, B.; Dlamini, B.; Beswa, D. Effect of Common Cooking and Drying Methods on Phytochemical and Antioxidant Properties of Corchorus olitorius Identified Using Liquid Chromatography-Mass Spectrometry (LC-MS). Molecules 2022, 27, 9052. https://doi.org/10.3390/molecules27249052
Ncube B, Dlamini B, Beswa D. Effect of Common Cooking and Drying Methods on Phytochemical and Antioxidant Properties of Corchorus olitorius Identified Using Liquid Chromatography-Mass Spectrometry (LC-MS). Molecules. 2022; 27(24):9052. https://doi.org/10.3390/molecules27249052
Chicago/Turabian StyleNcube, Buyile, Bhekisisa Dlamini, and Daniso Beswa. 2022. "Effect of Common Cooking and Drying Methods on Phytochemical and Antioxidant Properties of Corchorus olitorius Identified Using Liquid Chromatography-Mass Spectrometry (LC-MS)" Molecules 27, no. 24: 9052. https://doi.org/10.3390/molecules27249052
APA StyleNcube, B., Dlamini, B., & Beswa, D. (2022). Effect of Common Cooking and Drying Methods on Phytochemical and Antioxidant Properties of Corchorus olitorius Identified Using Liquid Chromatography-Mass Spectrometry (LC-MS). Molecules, 27(24), 9052. https://doi.org/10.3390/molecules27249052