
Comprehensive Detoxification Mechanism Assessment of Red Imported Fire Ant (Solenopsis invicta) against Indoxacarb

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Toxicity Bioassay
2.2. Enzyme Activity
2.3. Transcriptional Analysis of Indoxacarb Treated S. invicta
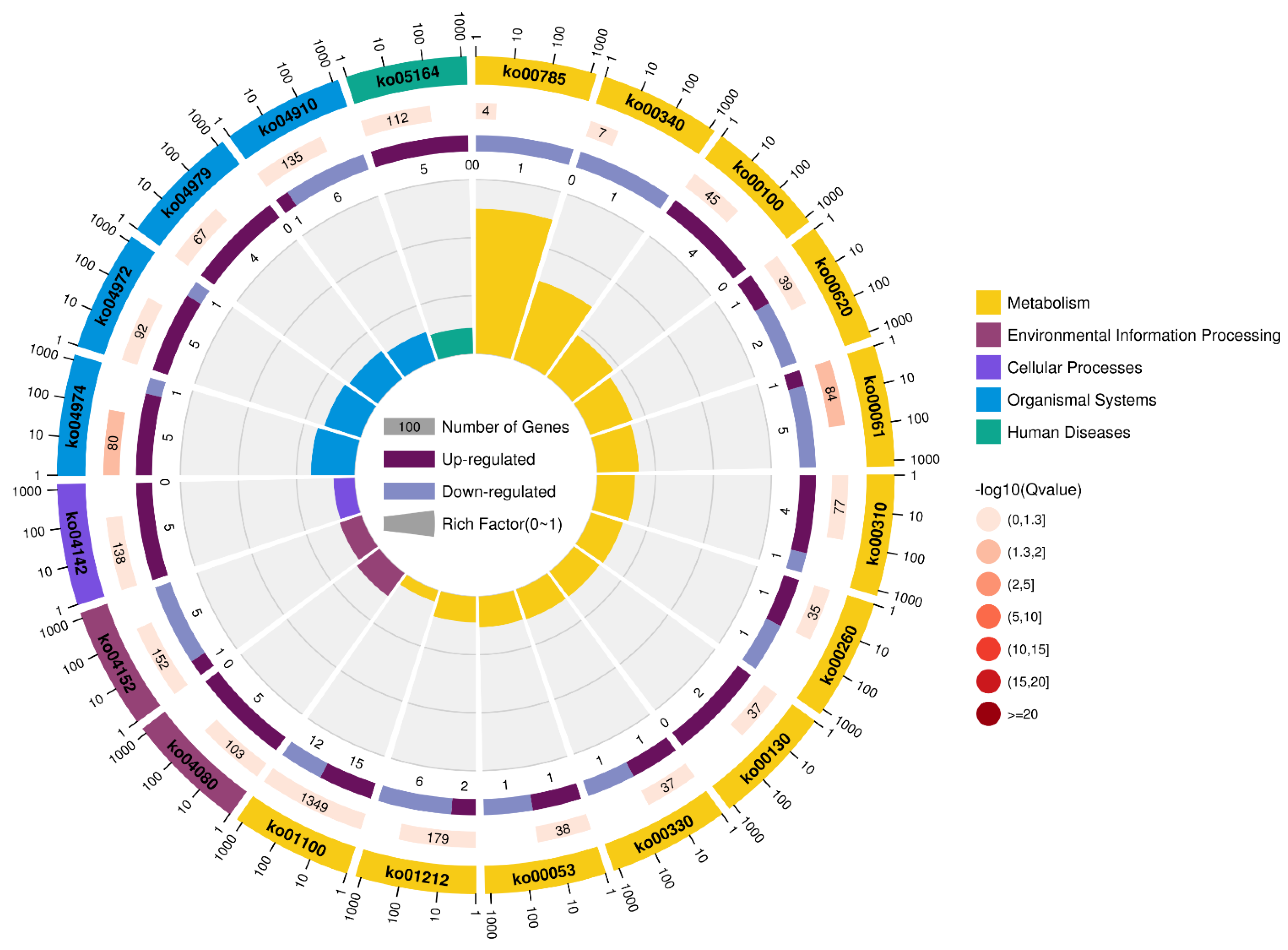
2.4. KEGG Analysis
2.5. KEGG Analysis of DEGs
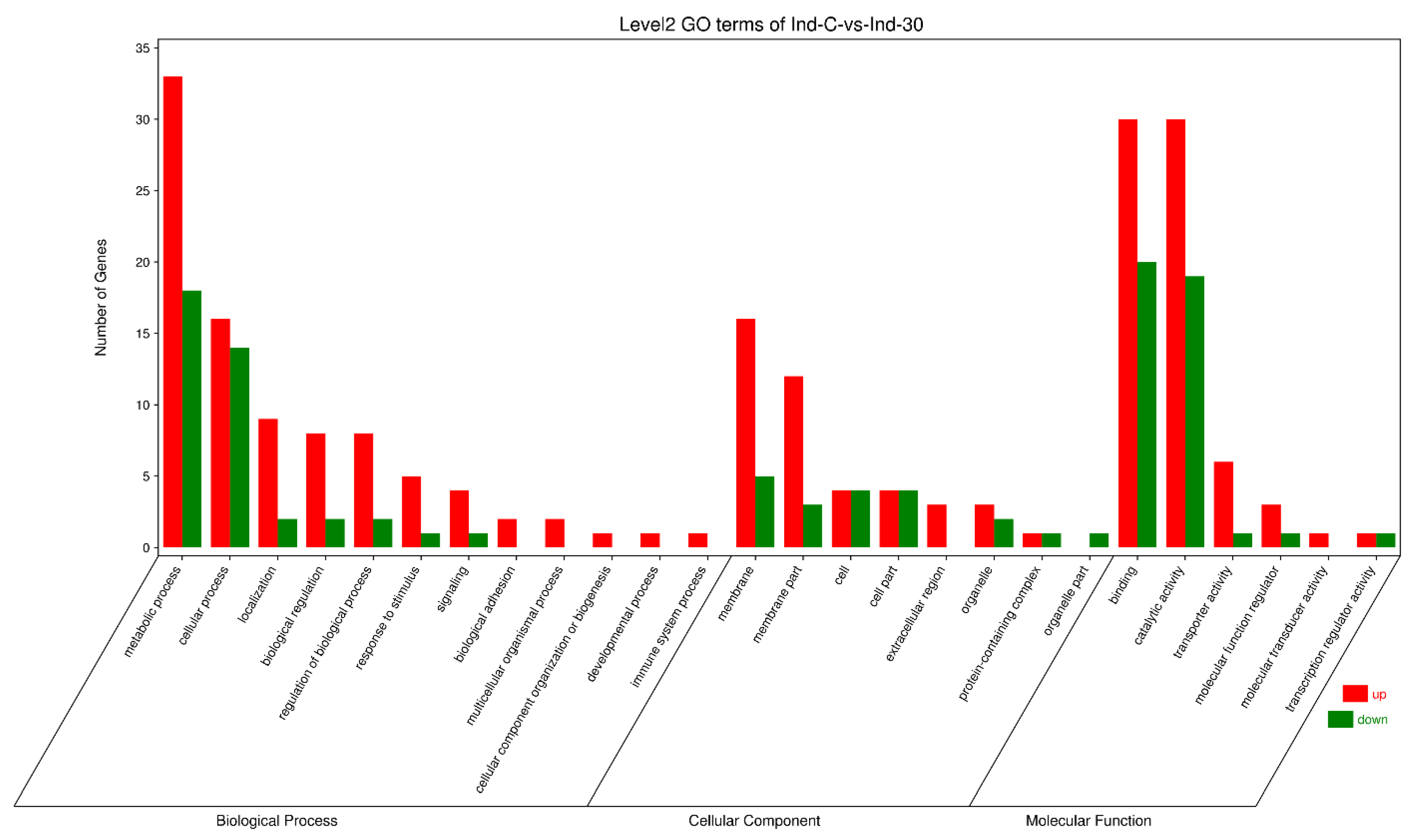
2.6. Highly Regulated GO Terms
2.7. Highly Regulated GO Terms in Biological Processes
2.8. Highly Regulated GO Term in the Cellular Component
2.9. Highly Regulated GO Terms in Molecular Function
2.10. Detoxification Mechanisms Related to Genes
2.11. Validation of Transcriptome via RT-qPCR
3. Discussion
4. Materials and Methods
4.1. Test Insects
4.2. Toxicity Bioassay
4.3. Enzyme Assay
4.4. RNA Extraction, Library Construction, and Sequencing
4.5. Bioinformatics Analysis
4.5.1. Alignment of Ribosome RNA (rRNA)
4.5.2. Quantification of Gene Abundance
4.5.3. Differentially Expressed Genes (DEGs)
4.5.4. Relationship Analysis of Samples
GO Enrichment Analysis
Pathway Enrichment Analysis
4.6. RT-qPCR for Transcriptome Validation
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Siddiqui, J.A.; Bamisile, B.S.; Khan, M.M.; Islam, W.; Hafeez, M. Impact of invasive ant species on native fauna across similar habitats under global environmental changes. Environ. Sci. Pollut. Res. 2021, 28, 54362–54382. [Google Scholar] [CrossRef]
- Bertelsmeier, C.; Ollier, S.; Liebhold, A.; Keller, L. Recent human history governs global ant invasion dynamics. Nat. Ecol. Evol. 2017, 1, 0184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, A.V.; McGlynn, T.P.; Tsutsui, N.D. Biogeographic and taxonomic patterns of introduced ants. Ant Ecol. 2010, 18, 233–244. [Google Scholar]
- Wang, L.; Zeng, L.; Xu, Y.; Lu, Y. Prevalence and management of Solenopsis invicta in China. NeoBiota 2020, 54, 89–124. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.M.; Tseng, Y.C.; Chen, C.T.; Lin, C.C.; Lee, Y.T.; Chen, Y.Y. Use of liquid nitrogen to treat Solenopsis invicta (Hymenoptera: Formicidae) nests. Fla. Entomol. 2013, 96, 871–876. [Google Scholar] [CrossRef]
- Zeng, L.; Lu, Y.; He, X.; Zhang, W.; Liang, G. Identification of red imported fire ant, Solenopsis invicta, to invade mainland China and infestation in Wuchuan, Guangdong. Chin. Bull. Entomol. 2005, 42, 144–148. [Google Scholar]
- Wu, D.; Zeng, L.; Lu, Y.; Xu, Y. Effects of Solenopsis invicta (Hymenoptera: Formicidae) and its interaction with aphids on the seed productions of mungbean and rapeseed plants. J. Econ. Entomol. 2014, 107, 1758–1764. [Google Scholar] [CrossRef]
- Lu, Y.Y. Long-distance spreading speed and trend prediction of red imported fire ant, Solenopsis invicta Buren, in mainland China. Guangdong Agric. Sci. 2014, 10, 70–72. [Google Scholar]
- Wang, L.; Xu, Y.; Zeng, L.; Lu, Y. Impact of the red imported fire ant Solenopsis invicta Buren on biodiversity in South China: A review. J. Integr. Agric. 2019, 18, 788–796. [Google Scholar] [CrossRef]
- Jenkins, M.A. Impact of the balsam woolly adelgid (Adelges piceae Ratz.) on an Abies fraseri (Pursh) Poir. dominated stand near the summit of Mount LeConte, Tennessee. Castanea 2003, 68, 109–118. [Google Scholar]
- Boettner, G.H.; Elkinton, J.S.; Boettner, C.J. Effects of a biological control introduction on three non target native species of saturniid moths. Conserv. Biol. 2000, 14, 1798–1806. [Google Scholar] [CrossRef]
- Snyder, W.E.; Evans, E.W. Ecological effects of invasive arthropod generalist predators. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 95–122. [Google Scholar] [CrossRef] [Green Version]
- Jensen, A.B.; Palmer, K.A.; Boomsma, J.J.; Pedersen, B.V. Varying degrees of Apis mellifera ligustica introgression in protected populations of the black honeybee, Apis mellifera mellifera, in northwest Europe. Mol. Ecol. 2005, 14, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Fournier, A.; Penone, C.; Pennino, M.G.; Courchamp, F. Predicting future invaders and future invasions. Proc. Natl. Acad. Sci. USA 2019, 116, 7905–7910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelsmeier, C.; Ollier, S.; Liebhold, A.M.; Brockerhoff, E.G.; Ward, D.; Keller, L. Recurrent bridgehead effects accelerate global alien ant spread. Proc. Natl. Acad. Sci. USA 2018, 115, 5486–5491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drees, B.M.; Calixto, A.A.; Nester, P.R. Integrated pest management concepts for red imported fire ants Solenopsis invicta (Hymenoptera: Formicidae). Insect Sci. 2013, 20, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Grodowitz, M.J.; Chen, J. Insecticidal and enzyme inhibitory activities of isothiocyanates against red imported fire ants, Solenopsis invicta. Biomolecules 2020, 10, 716. [Google Scholar] [CrossRef] [PubMed]
- Wing, K.D.; Sacher, M.; Kagaya, Y.; Tsurubuchi, Y.; Mulderig, L.; Connair, M.; Schnee, M. Bioactivation and mode of action of the oxadiazine indoxacarb in insects. Crop Prot. 2000, 19, 537–545. [Google Scholar] [CrossRef]
- Wang, X.; Su, W.; Zhang, J.; Yang, Y.; Dong, K.; Wu, Y. Two novel sodium channel mutations associated with resistance to indoxacarb and metaflumizone in the diamondback moth, Plutella xylostella. Insect Sci. 2016, 23, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- De França, S.M.; Breda, M.O.; Barbosa, D.R.S.; Araujo, A.M.N.; Guedes, C.A. The Sublethal Effects of Insecticides in Insects. In Biological Control of Pest and Vector Insects; BoD—Books on Demand: Norderstedt, Germany, 2017; pp. 23–39. [Google Scholar]
- Khan, M.M.; Khan, A.H.; Ali, M.W.; Hafeez, M.; Ali, S.; Du, C.; Fan, Z.; Sattar, M.; Hua, H. Emamectin benzoate induced enzymatic and transcriptional alternation in detoxification mechanism of predatory beetle Paederus fuscipes (Coleoptera: Staphylinidae) at the sublethal concentration. Ecotoxicology 2021, 30, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Hafeez, M.; Siddiqui, J.A.; Ullah, F.; Shah, S.; Iftikhar, A.; Rehman, S.U.; Rajput, S.A.; Ali, D.; Almarzoug, M.H.A.; et al. Residual toxicity and low lethal effects of fenvalerate on the development and physiology of Spodoptera exigua reared on different hosts. J. King Saud Univ.-Sci. 2021, 33, 101593. [Google Scholar] [CrossRef]
- Rasheed, M.A.; Khan, M.M.; Hafeez, M.; Zhao, J.; Islam, Y.; Ali, S.; Ur-Rehman, S.; Zhou, X. Lethal and sublethal effects of chlorpyrifos on biological traits and feeding of the aphidophagous predator Harmonia axyridis. Insects 2020, 11, 491. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, L.; Cui, R.; Zeng, X.; Gao, X. Cloning and expression of multiple cytochrome P450 genes: Induction by fipronil in workers of the red imported fire ant (Solenopsis invicta Buren). PLoS ONE 2016, 11, e0150915. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yang, Y.; Yuan, G.; Campbell, P.M.; Teese, M.G.; Russell, R.J.; Oakeshott, J.G.; Wu, Y. Overexpressed esterases in a fenvalerate resistant strain of the cotton bollworm, Helicoverpa armigera. Insect Biochem. Mol. Biol. 2011, 41, 14–21. [Google Scholar] [CrossRef]
- Ishaaya, I. Insect detoxifying enzymes: Their importance in pesticide synergism and resistance. Arch. Insect Biochem. Physiol. 1993, 22, 263–276. [Google Scholar] [CrossRef]
- Lin, T.; Cai, Z.; Wu, H. Transcriptome analysis of the Japanese pine sawyer beetle, Monochamus alternatus (Coleoptera: Cerambycidae) by high-throughput Illumina sequencing. J. Asia-Pac. Entomol. 2015, 18, 439–445. [Google Scholar] [CrossRef]
- Wang, J.J.; Cheng, W.X.; Ding, W.; Zhao, Z.M. The effect of the insecticide dichlorvos on esterase activity extracted from the psocids, Liposcelis bostrychophila and L. entomophila. J. Insect Sci. 2004, 4, 23. [Google Scholar] [CrossRef]
- Wu, H.; Gao, C.; Guo, Y.; Zhang, Y.; Zhang, J.; Ma, E. Acute toxicity and sublethal effects of fipronil on detoxification enzymes in juvenile zebrafish (Danio rerio). Pestic. Biochem. Physiol. 2014, 115, 9–14. [Google Scholar] [CrossRef]
- Ismail, S.M. Effect of sublethal doses of some insecticides and their role on detoxication enzymes and protein-content of Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae). Bull. Natl. Res. Cent. 2020, 44, 35. [Google Scholar] [CrossRef]
- Gang, W.; Shuren, J. Resistance mechanisms to methamidosphos and methomyl in the parasitoid Diaeretiella rapae (M’Intosh). Kun Chong xue bao. Acta Entomol. Sin. 2003, 46, 292–298. [Google Scholar]
- Hu, Z.; Xia, F.; Lin, Q.; Chen, H.; Li, Z.; Fei, Y.I.N.; Liang, P.; Gao, X. Biochemical mechanism of chlorantraniliprole resistance in the diamondback moth, Plutella xylostella Linnaeus. J. Integr. Agric. 2014, 13, 2452–2459. [Google Scholar] [CrossRef] [Green Version]
- Feyereisen, R. Molecular biology of insecticide resistance. Toxicol. Lett. 1995, 82, 83–90. [Google Scholar] [CrossRef]
- Tang, Z.H.; Gong, K.Y.; You, Z.P. Present status and countermeasures of insecticide resistance in agricultural pests in China. Pestic. Sci. 1988, 23, 189–198. [Google Scholar] [CrossRef]
- Mao, K.; Ren, Z.; Li, W.; Cai, T.; Qin, X.; Wan, H.; Jin, B.R.; He, S.; Li, J. Carboxylesterase genes in nitenpyram-resistant brown planthoppers, Nilaparvata lugens. Insect Sci. 2021, 28, 1049–1060. [Google Scholar] [CrossRef]
- Wang, K.; Huang, Y.; Li, X.; Chen, M. Functional analysis of a carboxylesterase gene associated with isoprocarb and cyhalothrin resistance in Rhopalosiphum padi (L.). Front. Physiol. 2018, 9, 992. [Google Scholar] [CrossRef]
- Zibaee, A.; Bandani, A.R.; Tork, M. Effect of the entomopathogenic fungus, Beauveria bassiana, and its secondary metabolite on detoxifying enzyme activities and acetylcholinesterase (AChE) of the sunn pest, Eurygaster integriceps (Heteroptera: Scutellaridae). Biocontrol Sci. Technol. 2009, 19, 485–498. [Google Scholar] [CrossRef]
- Simon, J.Y. Insect glutathione S-transferases. Zool. Stud. 1996, 35, 9–19. [Google Scholar]
- Zibaee, A.; Sendi, J.J.; Etebari, K.; Alinia, F.; Ghadamyari, M. The effect of diazinon on some biochemical characteristics of Chilo suppressalis Walker (Lepidoptera: Pyralidae), rice striped stem borer. Munis Entomol. Zool. 2008, 3, 255–265. [Google Scholar]
- Xiong, T.; Ling, S.; Liu, J. Insecticidal and P450 mediate mechanism of furalaner against red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae). Res. Sq. 2020, 25. [Google Scholar]
- Serebrov, V.V.; Gerber, O.N.; Malyarchuk, A.A.; Martemyanov, V.V.; Alekseev, A.A.; Glupov, V.V. Effect of entomopathogenic fungi on detoxification enzyme activity in greater wax moth Galleria mellonella L. (Lepidoptera, Pyralidae) and role of detoxification enzymes in development of insect resistance to entomopathogenic fungi. Biol. Bull. 2006, 33, 581–586. [Google Scholar] [CrossRef]
- Khan, M.M.; Kaleem-Ullah, R.M.; Siddiqui, J.A.; Ali, S. Insecticide resistance and detoxification enzymes activity in Nilaparvata lugens Stål against neonicotinoids. J. Agric. Sci. 2020, 12, 24–36. [Google Scholar] [CrossRef]
- Barres, B.; Corio-Costet, M.F.; Debieu, D.; Délye, C.; Fillinger-David, S.; Grosman, J.; Micoud, A.; Siegwart, M.; Walker, A.S. Trends and challenges in pesticide resistance detection. Trends Plant Sci. 2016, 21, 834–853. [Google Scholar]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhang, L.; Yang, S.; Xie, B.; An, S.; Liang, G. Assessment of the lethal and sublethal effects by spinetoram on cotton bollworm. PLoS ONE 2018, 13, e0204154. [Google Scholar] [CrossRef]
- Li, Y.H.; Wang, X.Y.; Hua, W.; Zhang, H. Studies on dissipations and residues of indoxacarb under different field and environmental conditions. J. Anal. Methods Chem. 2020, 2020, 8874759. [Google Scholar] [CrossRef]
- Jayakrishnan, S.; Madhuban, G. Evaluation of synthetic and natural insecticides for the management of insect pest control of eggplant (Solanum melongena L.) and pesticide residue dissipation pattern. Am. J. Plant Sci. 2012, 3, 214–227. [Google Scholar]
- Wing, K.D. It takes a team: Reflections on insecticide discoveries, toxicological problems and enjoying the unexpected. Pest Manag. Sci. 2017, 73, 666–671. [Google Scholar] [CrossRef]
- Mahmoudvand, M.; Abbasipour, H.; Garjan, A.S.; Bandani, A.R. Sublethal effects of indoxacarb on the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae). Appl. Entomol. Zool. 2011, 46, 75–80. [Google Scholar] [CrossRef]
- Shono, T.; Zhang, L.; Scott, J.G. Indoxacarb resistance in the house fly, Musca domestica. Pestic. Biochem. Physiol. 2004, 80, 106–112. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Shen, J.; Li, D.; Wan, H.; You, H.; Li, J. Cross-resistance and biochemical mechanisms of resistance to indoxacarb in the diamondback moth, Plutella xylostella. Pestic. Biochem. Physiol. 2017, 140, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.J.; McCord Jr, E. Lack of cross-resistance to indoxacarb in insecticide-resistant Spodoptera frugiperda (Lepidoptera: Noctuidae) and Plutella xylostella (Lepidoptera: Yponomeutidae). Pest Manag. Sci. Former Pestic. Sci. 2007, 63, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Sayyed, A.H.; Ahmad, M.; Saleem, M.A. Cross-resistance and genetics of resistance to indoxacarb in Spodoptera litura (Lepidoptera: Noctuidae). J. Econ. Entomol. 2014, 101, 472–479. [Google Scholar] [CrossRef]
- Su, J.; Sun, X.-X. High level of metaflumizone resistance and multiple insecticide resistance in field populations of Spodoptera exigua (Lepidoptera: Noctuidae) in Guangdong Province, China. Crop Prot. 2014, 61, 58–63. [Google Scholar] [CrossRef]
- Xiong, T.; Qiu, X.; Ling, S.; Liu, J.; Zeng, X. Interaction of fipronil and the red imported fire ant (Solenopsis invicta): Toxicity differences and detoxification responses. J. Insect Physiol. 2019, 115, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Wang, Q.; Qi, H.; Wang, Q.; Yuan, H.; Rui, C. Resistance selection of indoxacarb in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae): Cross-resistance, biochemical mechanisms and associated fitness costs. Pest Manag. Sci. 2018, 74, 2636–2644. [Google Scholar] [CrossRef]
- Salman, S.Y.; Recep, A.Y. Analysis of hexythiazox resistance mechanisms in a laboratory selected predatory mite Neoseiulus californicus (Acari: Phytoseiidae). Turk. Entomol. Derg. 2013, 37, 409–422. [Google Scholar]
- Pang, S.; You, W.; Duan, L.; Song, X.; Li, X.; Wang, C. Resistance selection and mechanisms of oriental tobacco budworm (Helicoverpa assulta Guenee) to indoxacarb. Pestic. Biochem. Physiol. 2012, 103, 219–223. [Google Scholar] [CrossRef]
- Gao, M.; Mu, W.; Wang, W.; Zhou, C.; Li, X. Resistance mechanisms and risk assessment regarding indoxacarb in the beet armyworm, Spodoptera exigua. Phytoparasitica 2014, 42, 585–594. [Google Scholar] [CrossRef]
- Nehare, S.; Moharil, M.P.; Ghodki, B.S.; Lande, G.K.; Bisane, K.D.; Thakare, A.S.; Barkhade, U.P. Biochemical analysis and synergistic suppression of indoxacarb resistance in Plutella xylostella L. J. Asia Pac. Entomol. 2010, 13, 91–95. [Google Scholar] [CrossRef]
- Hafeez, M.; Li, X.; Zhang, Z.; Huang, J.; Wang, L.; Zhang, J.; Shah, S.; Khan, M.M.; Xu, F.; Fernández-Grandon, G.M.; et al. De novo transcriptomic analyses revealed some detoxification genes and related pathways responsive to poposion yihaogong® 5% EC (Lambda-cyhalothrin 5%) exposure in Spodoptera frugiperda third-instar larvae. Insects 2021, 12, 132. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, J.; Shen, G.; Xu, Z.; Wei, P.; Zhang, Y.; Xu, Q.; He, L. Silencing NADPH-cytochrome P450 reductase results in reduced acaricide resistance in Tetranychus cinnabarinus (Boisduval). Sci. Rep. 2015, 5, 15581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Shi, Y.; Zhang, Y.; Liao, X. A systemic study of indoxacarb resistance in Spodoptera litura revealed complex expression profiles and regulatory mechanism. Sci. Rep. 2019, 9, 14997. [Google Scholar] [CrossRef] [Green Version]
- Pan, P.L.; Ye, Y.X.; Lou, Y.H.; Lu, J.B.; Cheng, C.; Shen, Y.; Moussian, B.; Zhang, C.X. A comprehensive omics analysis and functional survey of cuticular proteins in the brown planthopper. Proc. Natl. Acad. Sci. USA 2018, 115, 5175–5180. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Wang, W.; Zhang, D.; Lv, Y.; Zhou, D.; Ma, L.; Shen, B.; Sun, Y.; Zhu, C. The cuticle proteins: A putative role for deltamethrin resistance in Culex pipiens pallens. Parasitol. Res. 2015, 114, 4421–4429. [Google Scholar] [CrossRef]
- Wei, N.; Zhong, Y.; Lin, L.; Xie, M.; Zhang, G.; Su, W.; Li, C.; Chen, H. Transcriptome analysis and identification of insecticide tolerance-related genes after exposure to insecticide in Sitobion avenae. Genes 2019, 10, 951. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Zhou, C.; Yang, X.-B.; Long, G.Y.; Jin, D.C. Effects of insecticide stress on expression of NLABCG transporter gene in the brown planthopper, Nilaparvata lugens. Insects 2019, 10, 334. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Yang, H.; Wang, Z.; Long, G.Y.; Jin, D.C. Protective and detoxifying enzyme activity and ABCG subfamily gene expression in Sogatella furcifera under insecticide stress. Front. Physiol. 2019, 9, 1890. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Yan, Z.; Si, F.; Zhou, Y.; Fu, W.; Chen, B. ATP-binding cassette (ABC) transporter genes involved in pyrethroid resistance in the malaria vector Anopheles sinensis: Genome-wide identification, characteristics, phylogenetics, and expression profile. Int. J. Mol. Sci. 2019, 20, 1409. [Google Scholar] [CrossRef] [Green Version]
- Nadal, A.; Quesada, I.; Tudurí, E.; Nogueiras, R.; Alonso-Magdalena, P. Endocrine-disrupting chemicals and the regulation of energy balance. Nat. Rev. Endocrinol. 2017, 13, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Bandookwala, M.; Nemani, K.S.; Chatterjee, B.; Sengupta, P. Reactive metabolites: Generation and estimation with electrochemistry based analytical strategy as an emerging screening tool. Curr. Anal. Chem. 2020, 16, 811–825. [Google Scholar] [CrossRef]
- Kafle, L.; Wu, W.J.; Shiha, C.J. A new fire ant (Hymenoptera: Formicidae) bait base carrier for moist conditions. Pest Manag. Sci. 2010, 66, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. Advancement on techniques for the separation and maintenance of the red imported fire ant colonies. Insect Sci. 2007, 14, 1–4. [Google Scholar] [CrossRef]
- Ning, D.; Yang, F.; Xiao, Q.; Ran, H.; Xu, Y. A simple and efficient method for preventing ant escape (Hymenoptera: Formicidae). Myrmecol. News 2019, 29, 57–65. [Google Scholar]
- Seagraves, M.P.; McPherson, R.M. Residual susceptibility of the red imported fire ant (Hymenoptera: Formicidae) to four agricultural insecticides. J. Econ. Entomol. 2003, 96, 645–648. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | n | LC10 (%) (95% CL) | LC30 (%) (95% CL) | LC50 (%) (95% CL) | Slope ± SE | X2 (df) |
|---|---|---|---|---|---|---|
| GZ-1 | 1350 | 0.012 (0.010–0.013) a | 0.015 (0.014–0.016) a | 0.018 (0.016–0.019) a | 12.174 ± 0.0.64 | 24.607 (8) |
| GZ-2 | 2430 | 0.013 (0.011–0.014) ab | 0.017 (0.015–0.018) ab | 0.020 (0.018–0.021) a | 12.281 ± 0.480 | 42.373 (8) |
| HY | 1200 | 0.005 (0.004–0.006) ab | 0.008 (0.007–0.009) ab | 0.011 (0.011–0.012) ab | 07.255 ± 0.518 | 05.280 (7) |
| HZ | 1200 | 0.007 (0.005–0.008) ab | 0.010 (0.009–0.012) ab | 0.014 (0.012–0.015) ab | 07.552 ± 0.522 | 13.966 (7) |
| SZ | 960 | 0.004 (0.003–0.005) b | 0.007 (0.006–0.008) b | 0.011 (0.010–0.012) ab | 06.439 ± 0.564 | 06.680 (7) |
| ZS | 2160 | 0.007 (0.006–0.009) ab | 0.010 (0.008–0.011) ab | 0.012 (0.011–0.013) ab | 11.659 ± 0.466 | 33.651 (7) |
| ZH | 2430 | 0.004 (0.003–0.005) b | 0.007 (0.005–0.008) b | 0.009 (0.008–0.010) b | 08.045 ± 0.319 | 42.538 (8) |
| DG | 2160 | 0.007 (0.006–0.008) ab | 0.010 (0.008–0.010) ab | 0.012 (0.011–0.013) ab | 11.501 ± 0.464 | 23.284 (7) |
| JM | 2160 | 0.007 (0.005–0.009) ab | 0.010 (0.009–0.012) ab | 0.013 (0.012–0.015) ab | 09.860 ± 0.426 | 44.507 (7) |
| Insecticide | Detoxification Enzyme Family | Total Genes | Upregulated Genes | Downregulated Genes |
|---|---|---|---|---|
| Indoxacarb | Acetylcholinesterase | 8 | 6 | 2 |
| Carboxylesterase | 2 | 2 | 0 | |
| Glutathione S-transferases | 13 | 6 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddiqui, J.A.; Zhang, Y.; Luo, Y.; Bamisile, B.S.; Rehman, N.U.; Islam, W.; Qasim, M.; Jiang, Q.; Xu, Y. Comprehensive Detoxification Mechanism Assessment of Red Imported Fire Ant (Solenopsis invicta) against Indoxacarb. Molecules 2022, 27, 870. https://doi.org/10.3390/molecules27030870
Siddiqui JA, Zhang Y, Luo Y, Bamisile BS, Rehman NU, Islam W, Qasim M, Jiang Q, Xu Y. Comprehensive Detoxification Mechanism Assessment of Red Imported Fire Ant (Solenopsis invicta) against Indoxacarb. Molecules. 2022; 27(3):870. https://doi.org/10.3390/molecules27030870
Chicago/Turabian StyleSiddiqui, Junaid Ali, Yuping Zhang, Yuanyuan Luo, Bamisope Steve Bamisile, Naveed Ur Rehman, Waqar Islam, Muhammad Qasim, Qiuying Jiang, and Yijuan Xu. 2022. "Comprehensive Detoxification Mechanism Assessment of Red Imported Fire Ant (Solenopsis invicta) against Indoxacarb" Molecules 27, no. 3: 870. https://doi.org/10.3390/molecules27030870
APA StyleSiddiqui, J. A., Zhang, Y., Luo, Y., Bamisile, B. S., Rehman, N. U., Islam, W., Qasim, M., Jiang, Q., & Xu, Y. (2022). Comprehensive Detoxification Mechanism Assessment of Red Imported Fire Ant (Solenopsis invicta) against Indoxacarb. Molecules, 27(3), 870. https://doi.org/10.3390/molecules27030870