
The Antiviral Effects of Jasminin via Endogenous TNF-α and the Underlying TNF-α-Inducing Action

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
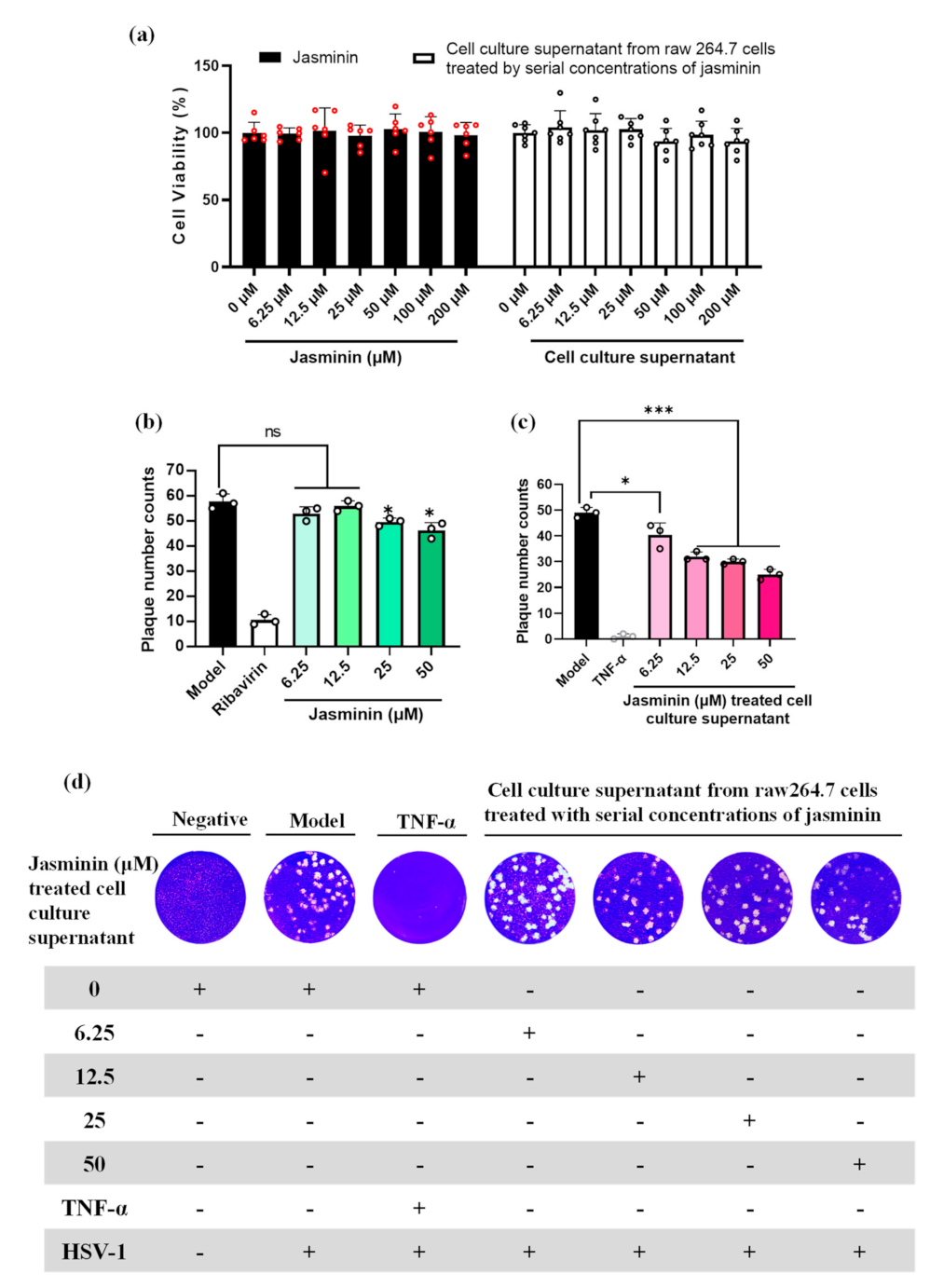
2.1. Jasminin Can Activate RAW 264.7 Cells and Induce TNF-α Production
2.2. Endogenous TNF-α Induced by Jasminin Has Potent Antiviral Activity against HSV-1
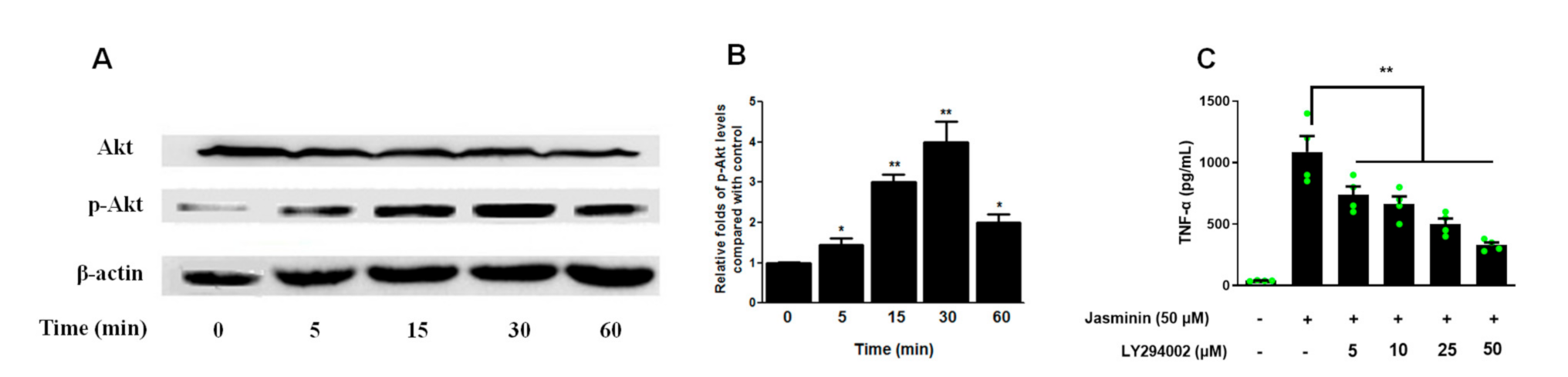
2.3. Akt Phosphorylation Mediates TNF-α Production Induced by Jasminin
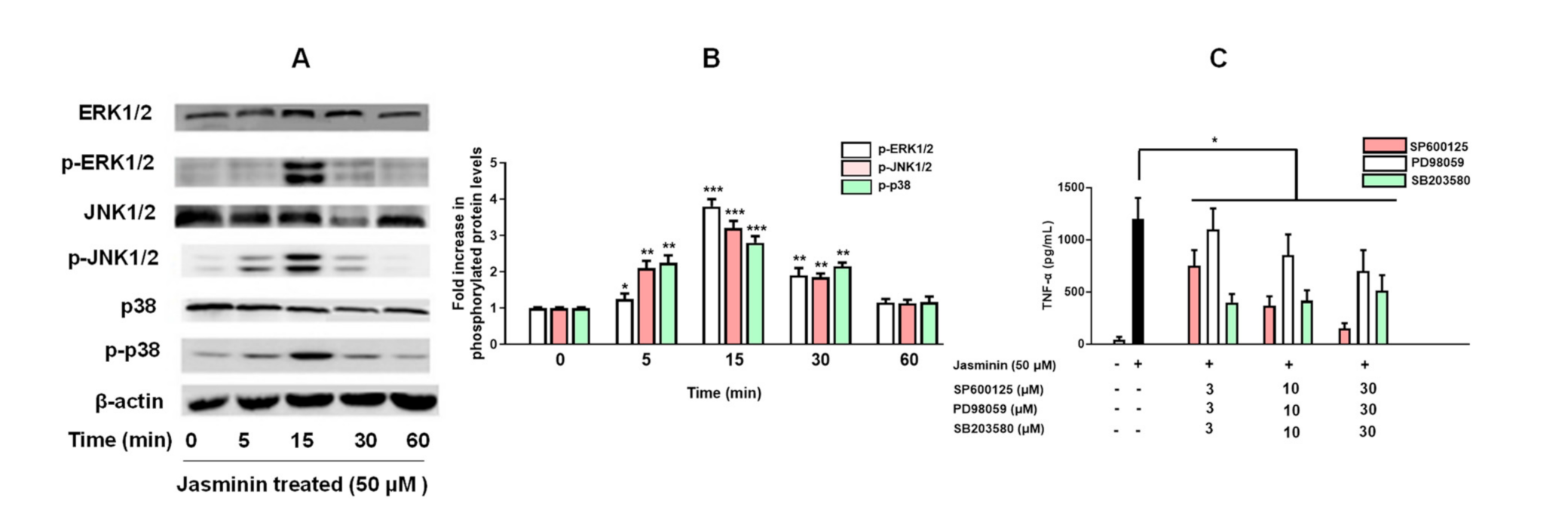
2.4. Phosphorylation of MAPKs by Jasminin to Induce TNF-α Production
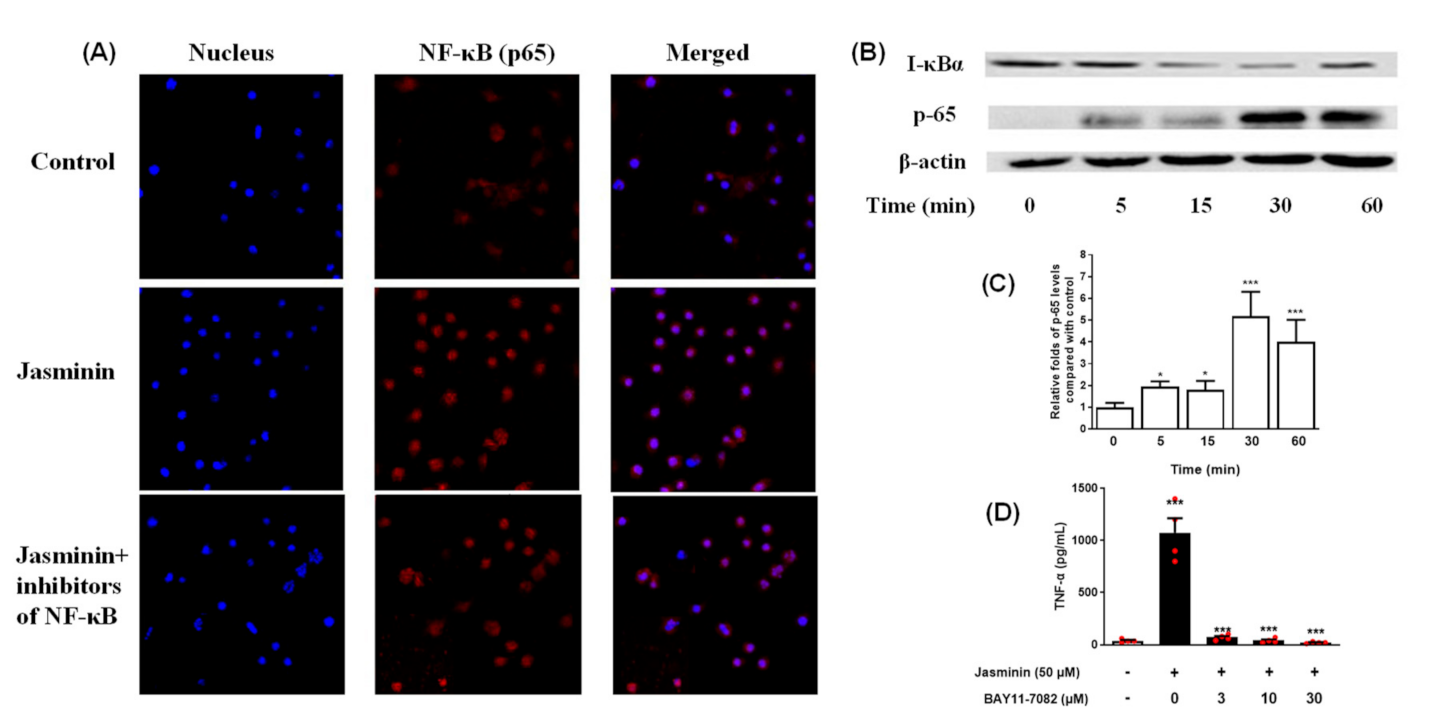
2.5. NF–κB Signal Transduction Induced by Jasminin in RAW 264.7 Cells
3. Discussion
4. Materials and Methods
4.1. Virus Strains
4.2. Cell Culture and Reagents
4.3. Cell Proliferation
4.4. In Vitro Stimulation for Cytokines Production
4.5. Plaque Reduction Assay
4.6. Western Blotting Analysis
4.7. Immunofluorescence Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Johnston, C.; Gottlieb, S.L.; Wald, A. Status of vaccine research and development of vaccines for herpes simplex virus. Vaccine 2016, 34, 2948–2952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Herpes Simplex Virus; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- Tian, T.; Wang, M.; Ma, D. TNF-α, a good or bad factor in hematological diseases? Stem Cell Investig. 2014, 1, 12. [Google Scholar] [PubMed]
- Matikainen, S.; Siren, J.; Tissari, J.; Veckman, V.; Pirhonen, J.; Severa, M.; Sun, Q.; Lin, R.T.; Meri, S.; Uze, G.; et al. Tumor necrosis factor alpha enhances influenza A virus-induced expression of antiviral cytokines by activating RIG-I gene expression. J. Virol. 2006, 80, 3515–3522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruby, J.; Bluethmann, H.; Peschon, J.J. Antiviral activity of tumor necrosis factor (TNF) is mediated via p55 and p75 TNF receptors. J. Exp. Med. 1997, 186, 1591–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minami, M.; Kita, M.; Yan, X.Q.; Yamamoto, T.; Iida, T.; Sekikawa, K.; Iwakura, Y.; Imanishi, J. Role of IFN-gamma and tumor necrosis factor-alpha in herpes simplex virus type 1 infection. J. Interf. Cytokine Res. 2002, 22, 671–676. [Google Scholar] [CrossRef]
- Lundberg, P.; Welander, P.V.; Edwards III, C.K.; van Rooijen, N.; Cantin, E. Tumor necrosis factor (TNF) protects resistant C57BL/6 mice against herpes simplex virus-induced encephalitis independently of signaling via TNF receptor 1 or 2. J. Virol. 2007, 81, 1451–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, H.; Saito, K.; Kanda, T.; Kobayashi, I.; Fujii, H.; Fujigaki, S.; Maekawa, N.; Takatsu, H.; Fujiwara, H.; Sekikawa, K.; et al. Tumor necrosis factor-alpha (TNF-alpha) plays a protective role in acute viral myocarditis in mice—A study using mice lacking TNF-alpha. Circulation 2001, 103, 743–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zidek, Z.; Anzenbacher, P.; Kmonickova, E. Current status and challenges of cytokine pharmacology. Br. J. Pharmacol. 2009, 157, 342–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Xiao, H.-T.; Liao, C.-H.; Li, L.; Kang, Q.-R.; Jiang, Y.-C.; Hu, X.-P.; Zheng, K.; Fan, L.; He, Z.-D. Natural Products: The Master Regulators of Antiviral Cytokines. Curr. Org. Chem. 2017, 21, 1805–1823. [Google Scholar] [CrossRef]
- Hur, S.J.; Kang, S.H.; Jung, H.S.; Kim, S.C.; Jeon, H.S.; Kim, I.H.; Lee, J.D. Review of natural products actions on cytokines in inflammatory bowel disease. Nutr Res. 2012, 32, 801–816. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.T.; Zeng, W.F.; Zhang, F.Y.; Zhang, C.L.; Liang, W. Naringenin ameliorates acute inflammation by regulating intracellular cytokine degradation. J. Immunol. 2017, 199, 3466–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; He, J.; Xu, H.; Hu, X.P.; Wu, X.L.; Wu, H.Q.; Liu, L.Z.; Liao, C.H.; Zeng, Y.; Li, Y.; et al. The antiviral effects of acteoside and the underlying IFN-γ-inducing action. Food Funct. 2016, 7, 3017–3030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- College, J.N.M. Jiangsu New Medical College, Dictionary of Chinese Materia Medica; Shanghai People’s Press: Shanghai, China, 1986. [Google Scholar]
- Duque, G.A.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar]
- Smith, M.V.; Lee, M.J.; Islam, A.S.; Rohrer, J.L.; Goldberg, V.M.; Beidelschies, M.A.; Greenfield, E.M. Inhibition of the PI3K-Akt signaling pathway reduces tumor necrosis factor-alpha production in response to titanium particles in vitro. J. Bone Jt. Surg. Am. 2007, 89A, 1019–1027. [Google Scholar]
- Akhtar, M.S.; Swamy, M.K.; Umar, A.; Al Sahli, A.A. Biosynthesis and Characterization of Silver Nanoparticles from Methanol Leaf Extract of Cassia didymobotyra and Assessment of Their Antioxidant and Antibacterial Activities. J. Nanosci. Nanotechnol. 2015, 15, 9818–9823. [Google Scholar] [CrossRef] [PubMed]
- Carswell, E.A.; Old, L.J.; Kassel, R.L.; Green, S.; Fiore, N.; Williamson, B. Endotoxin-Induced Serum Factor That Causes Necrosis of Tumors. Proc. Natl. Acad. Sci. USA 1975, 72, 3666–3670. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Solomon, D.H. Tumor necrosis factor blockade and the risk of viral infection. Nat. Rev. Rheumatol. 2010, 6, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Sedger, L.M.; McDermott, M.F. TNF and TNF-receptors: From mediators of cell death and inflammation to therapeutic giants—Past, present and future. Cytokine Growth Factor Rev. 2014, 25, 453–472. [Google Scholar] [CrossRef] [Green Version]
- Ahrens, P.B.; Besancon, F.; Memet, S.; Ankel, H. Tumor-Necrosis-Factor Enhances Induction by Beta-Interferon of a Ubiquitin Cross-Reactive Protein. J. Gen. Virol. 1990, 71, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.K.; Kaspar, T.A.; Coppenhaver, D.H. Synergy of Antiviral Actions of Tnf and Ifn-Gamma—Evidence for a Major Role of Tnf-Induced Ifn-Beta. Antivir. Res. 1988, 10, 1–9. [Google Scholar] [CrossRef]
- Wang, W.S.; Xu, L.; Brandsma, J.H.; Wang, Y.J.; Hakim, M.S.; Zhou, X.Y.; Yin, Y.B.; Fuhler, G.M.; van der Laan, L.J.W.; van der Woude, C.J.; et al. Convergent Transcription of Interferon-stimulated Genes by TNF-alpha and IFN-alpha Augments Antiviral Activity against HCV and HEV. Sci. Rep. 2016, 6, 25482. [Google Scholar] [CrossRef] [Green Version]
- Fink, K.; Martin, L.; Mukawera, E.; Chartier, S.; De Deken, X.; Brochiero, E.; Miot, F.; Grandvaux, N. IFNβ/TNFα synergism induces a non-canonical STAT2/IRF9-dependent pathway triggering a novel DUOX2 NADPH oxidase-mediated airway antiviral response. Cell Res. 2013, 23, 673–690. [Google Scholar] [CrossRef] [Green Version]
- Bartee, E.; Mohamed, M.R.; Lopez, M.C.; Baker, H.V.; McFadden, G. The Addition of Tumor Necrosis Factor plus Beta Interferon Induces a Novel Synergistic Antiviral State against Poxviruses in Primary Human Fibroblasts. J. Virol. 2009, 83, 498–511. [Google Scholar] [CrossRef] [Green Version]
- Lucin, P.; Jonjic, S.; Messerle, M.; Polic, B.; Hengel, H.; Koszinowski, U.H. Late-Phase Inhibition of Murine Cytomegalovirus Replication by Synergistic Action of Interferon-Gamma and Tumor-Necrosis-Factor. J. Gen. Virol. 1994, 75, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaiboullina, S.F.; Netski, D.M.; Krumpe, P.; St Jeor, S.C. Effects of tumor necrosis factor alpha on Sin Nombre virus infection in vitro. J. Virol. 2000, 74, 11966–11971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanza, I.; Brown, I.H.; Paton, D.J. Pathogenicity of Concurrent Infection of Pigs with Porcine Respiratory Coronavirus and Swine Influenza-Virus. Res. Vet. Sci. 1992, 53, 309–314. [Google Scholar] [CrossRef]
- Mestan, J.; Digel, W.; Mittnacht, S.; Hillen, H.; Blohm, D.; Möller, A.; Jacobsen, H.; Kirchner, H. Antiviral effects of recombinant tumour necrosis factor in vitro. Nature 1986, 323, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.H.; Goeddel, D.V. Tumour necrosis factors alpha and beta inhibit virus replication and synergize with interferons. Nature 1986, 323, 819–822. [Google Scholar] [CrossRef]
- Lane, B.R.; Markovitz, D.M.; Woodford, N.L.; Rochford, R.; Strieter, R.M.; Coffey, M.J. TNF-alpha inhibits HIV-1 replication in peripheral blood monocytes and alveolar macrophages by inducing the production of RANTES and decreasing C-C chemokine receptor 5 (CCR5) expression. J. Immunol. 1999, 163, 3653–3661. [Google Scholar] [PubMed]
- Van Campen, H. Influenza A virus replication is inhibited by tumor necrosis factor-alpha in vitro. Arch. Virol. 1994, 136, 439–446. [Google Scholar] [CrossRef]
- Lejeune, F.J.; Rüegg, C.; Liénard, D. Clinical applications of TNF-α in cancer. Curr. Opin. Immunol. 1998, 10, 573–580. [Google Scholar] [CrossRef]
- Riemer, A.B.; Hantusch, B.; Sponer, B.; Kraml, G.; Hafner, C.; Zielinski, C.C.; Scheiner, O.; Pehamberger, H.; Jensen-Jarolim, E. High-molecular weight melanoma-associated antigen mimotope immunizations induce antibodies recognizing melanoma cells. Cancer Immunol. Immunother. 2005, 54, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, P.; Yang, H.J.; Jung, S.J.; Behlke, M.A.; Rose, S.D.; Cantin, E.M. Protection against TNFalpha-dependent liver toxicity by intraperitoneal liposome delivered DsiRNA targeting TNFalpha in vivo. J. Control. Release 2012, 160, 194–199. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zhao, S.J.; Shen, S.T.; Fang, S.X.; Ye, Z.L.; Shi, Z.; Hong, A. A novel recombinant slow-release TNF α-derived peptide effectively inhibits tumor growth and angiogensis. Sci. Rep. 2015, 5, 13595. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, Y.; Sternberg, P.; Cai, J. Essential roles of the PI3 kinase/Akt pathway in regulating Nrf2-dependent antioxidant functions in the RPE. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1671–1678. [Google Scholar] [CrossRef] [Green Version]
- Means, T.K.; Pavlovich, R.P.; Roca, D.; Vermeulen, M.W.; Fenton, M.J. Activation of TNF-alpha transcription utilizes distinct MAP kinase pathways in different macrophage populations. J. Leukoc. Biol. 2000, 67, 885–893. [Google Scholar] [CrossRef]
- Rao, K.M.K. MAP kinase activation in macrophages. J. Leukoc. Biol. 2001, 69, 3–10. [Google Scholar] [PubMed]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, A.S. The NF-kappa B and I kappa B proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–683. [Google Scholar] [CrossRef] [Green Version]
- Lei, C.F.; Yang, J.; Hu, J.; Sun, X.L. On the Calculation of TCID50 for Quantitation of Virus Infectivity. Virol. Sin. 2021, 36, 141–144. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Hu, Z.; Yu, T.; Hu, H.; Zhao, Y.; Li, C.; Zhu, Q.; Wang, M.; Zhai, P.; He, L.; et al. The Antiviral Effects of Jasminin via Endogenous TNF-α and the Underlying TNF-α-Inducing Action. Molecules 2022, 27, 1598. https://doi.org/10.3390/molecules27051598
Zhu X, Hu Z, Yu T, Hu H, Zhao Y, Li C, Zhu Q, Wang M, Zhai P, He L, et al. The Antiviral Effects of Jasminin via Endogenous TNF-α and the Underlying TNF-α-Inducing Action. Molecules. 2022; 27(5):1598. https://doi.org/10.3390/molecules27051598
Chicago/Turabian StyleZhu, Xiaohong, Ziwei Hu, Tian Yu, Hao Hu, Yunshi Zhao, Chenyang Li, Qinchang Zhu, Mingzhong Wang, Peng Zhai, Longxia He, and et al. 2022. "The Antiviral Effects of Jasminin via Endogenous TNF-α and the Underlying TNF-α-Inducing Action" Molecules 27, no. 5: 1598. https://doi.org/10.3390/molecules27051598
APA StyleZhu, X., Hu, Z., Yu, T., Hu, H., Zhao, Y., Li, C., Zhu, Q., Wang, M., Zhai, P., He, L., Riaz Rajoka, M. S., Song, X., & He, Z. (2022). The Antiviral Effects of Jasminin via Endogenous TNF-α and the Underlying TNF-α-Inducing Action. Molecules, 27(5), 1598. https://doi.org/10.3390/molecules27051598