
Forensic Discrimination of Differentially Sourced Animal Blood Using a Bottom-Up Proteomics Based MALDI MS Approach




Abstract
:
1. Introduction
2. Results and Discussion
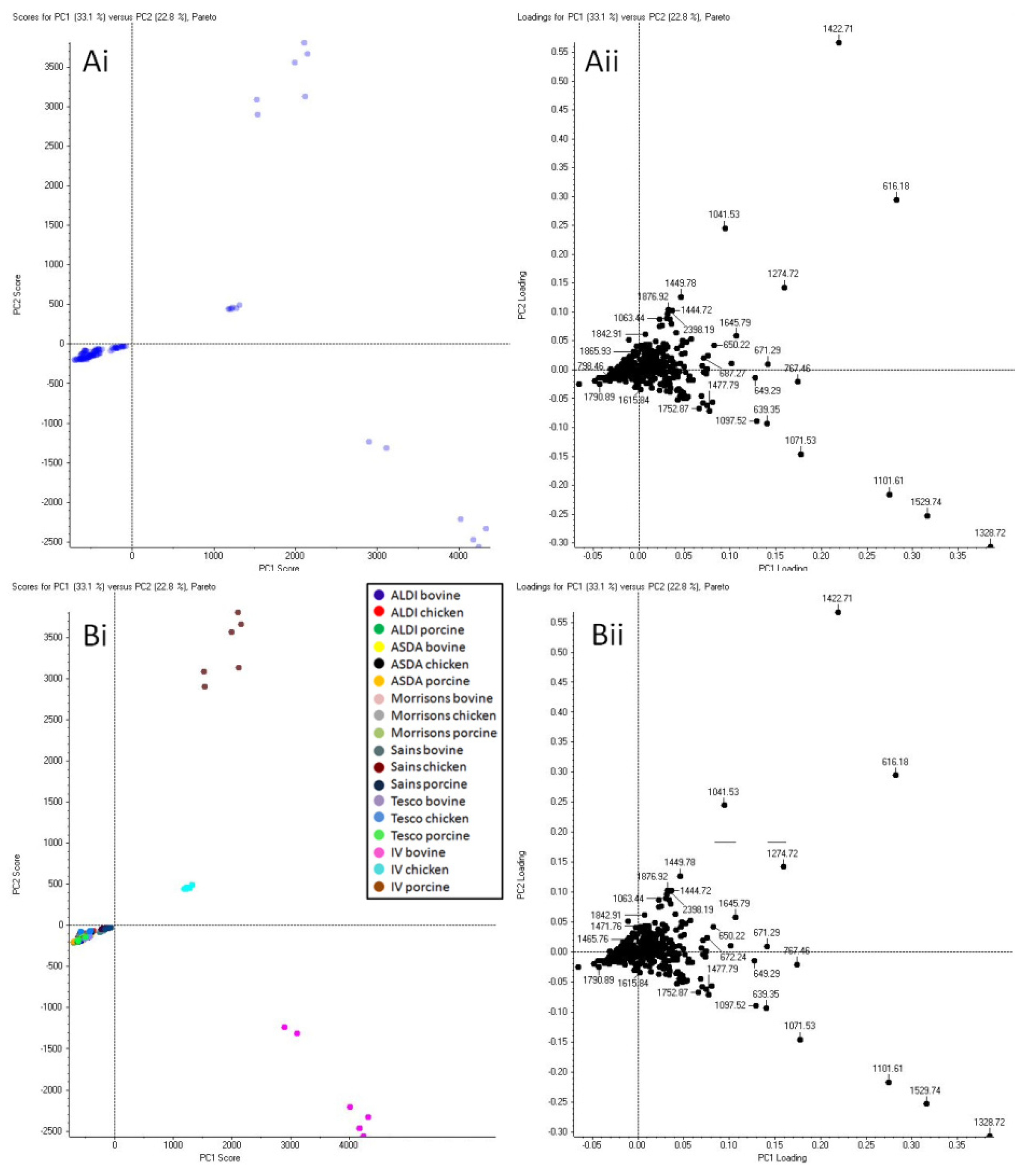
2.1. ID Level III: Differentiation between Blood Simulating a Wounded Animal (Intravenous) Versus Blood in Packaged Meat
2.1.1. Animal Species Determination from Intravenous Blood Simulating a Wounded Animal (Collected from the Jugular Vein)
ID Level III: Intravenous Bovine Blood Marker Identification
ID Level III: Intravenous Porcine Blood Marker Identification
ID Level III: Intravenous Chicken Blood Marker Identification
2.1.2. ID Level III: Statistical Analysis Discrimination between Intravenous Blood (Mimicking a Wounded Animal) Versus Blood in Packaged Raw Meat
2.2. ID Level III: Species Determination from Blood in Packaged Meat
2.2.1. ID Level III: Bovine Packaged Blood Marker Identification
2.2.2. ID Level III: Porcine Packaged Blood Marker Identification
2.2.3. ID Level III: Chicken Packaged Blood Marker Identification
2.3. ID Level IV: Assessment of the Potential to Identify the Retailer for the Animal Blood in Packaged Meat
3. Materials and Methods
3.1. Materials
3.2. Methods
Enzymatic Digestion of Blood
3.3. Instrumental Conditions
3.3.1. MALDI MS and MS/MS
3.3.2. Matrix and Application
3.3.3. Data Processing of MALDI MS Data
MALDI MS/MS Spectral Identification
Data Processing: Principal Component Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bossers, L.C.; Roux, C.; Bell, M.; McDonagh, A.M. Methods for the enhancement of fingermarks in blood. Forensic Sci. Int. 2011, 210, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, K.; Bengiat, R.; Heaton, C.; Herman, Y.; Oz, C.; Elad, M.L.; Cole, L.; Francese, S. “MALDI-CSI”: A proposed method for the tandem detection of human blood and DNA typing from enhanced fingermarks. Forensic Sci. Int. 2021, 323, 110774. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, R.; Bleay, S.; Clench, M.R.; Francese, S. Direct detection of blood in fingermarks by MALDI MS profiling and imaging. Sci. Justice 2014, 54, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Patel, E.; Cicatiello, P.; Deininger, L.; Clench, M.R.; Marino, G.; Giardina, P.; Langenburg, G.; West, A.; Marshall, P.; Sears, V.; et al. A proteomic approach for the rapid, multi-informative and reliable identification of blood. Analyst 2016, 141, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Zhou, B.; Deng, H.; Prinz, M.; Siegel, D. Body fluid identification by mass spectrometry. Int. J. Leg. Med. 2013, 127, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Sun, J.; Huang, X.; Shi, H.; Xiong, C.; Nie, Z. Direct identification of forensic body fluids by MALDI-MS. Analyst 2019, 144, 7017–7023. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, K.; Heaton, C.; Langenburg, G.; Cole, L.; Clark, T.; Clench, M.R.; Sears, V.; Sealey, M.; McColm, R.; Francese, S. Pre-validation of a MALDI MS proteomics-based method for the reliable detection of blood and blood provenance. Sci. Rep. 2020, 10, 17087. [Google Scholar] [CrossRef] [PubMed]
- Witt, M.; Kennedy, K.; Heaton, C.; Langenburg, G.; Francese, S. Forensic Visualisation of Blood and Blood Provenance in Old Fingermarks by MALDI MS Imaging. Available online: www.bruker.com/en/applications/academia-life-science/imaging/maldi-imaging/_jcr_content/root/contentpar/search.download-asset.pdf/144229c8-ea3b-4d82-8060-fc4d795b270f/1885564-msi-22-imaging-fingerprints-ebook-rev-01.pdf (accessed on 14 January 2022).
- Kamanna, S.; Henry, J.; Voelcker, N.; Linacre, A.; Kirkbride, K.P. “Bottom-up” in situ proteomic differentiation of human and non-human haemoglobins for forensic purposes by matrix-assisted laser desorption/ionization time-of-flight tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2017, 31, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Heaton, C.; Witt, M.; Cole, L.; Eyre, J.; Tazzyman, S.; McColm, R.; Francese, S. Detection and mapping of haemoglobin variants in blood fingermarks by MALDI MS for suspect “profiling”. Analyst 2021, 146, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Deininger, L.; Patel, E.; Clench, M.R.; Sears, V.; Sammon, C.; Francese, S. Proteomics goes forensic: Detection and mapping of blood signatures in fingermarks. Proteomics 2016, 16, 1707–1717. [Google Scholar] [CrossRef] [PubMed]
- Lecrenier, M.C.; Marbaix, H.; Dieu, M.; Veys, P.; Saegerman, C.; Raes, M.; Baeten, V. Identification of specific bovine blood biomarkers with a non-targeted approach using HPLC ESI tandem mass spectrometry. Food Chem. 2016, 213, 417–424. [Google Scholar] [CrossRef]
- TV Series Forensic Files, Chief Evidence, Season 7, Episode 17 Broadcast 24 July 2002. Available online: https://www.imdb.com/title/tt2610674/ (accessed on 20 January 2022).
- Monti, D.M.; De Simone, G.; Langella, E.; Supuran, C.T.; Di Fiore, A.; Monti, S.M. Insights into the role of reactive sulfhydryl groups of carbonic anhydrase III and VII during oxidative damage. J. Enzym. Inhib. Med. Chem. 2017, 32, 5–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strohalm, M.; Hassman, M.; Košata, B.; Kodíček, M. mMass data miner: An open source alternative for mass spectrometric data analysis. Rapid Commun. Mass Spectrom. 2008, 22, 905–908. [Google Scholar] [CrossRef]
- Strohalm, M.; Kavan, D.; Novak, P.; Volny, M.; Havlicek, V. mMass 3: A cross-platform software environment for precise analysis of mass spectrometric data. Anal. Chem. 2010, 82, 4648–4651. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.W.; Cagney, G.; Cartwright, H.M. SpecAlign—Processing and alignment of mass spectra datasets. Bioinformatics 2005, 21, 2088–2090. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
|
Experimental m/z |
Theoretical m/z | Putative Peptide Match | Mass Accuracy (ppm) | Proteotypic | Protein |
|---|---|---|---|---|---|
| 639.394 | 639.394 | VKAHGK | 0 | NO | βHb |
| 767.487 | 767.489 | VKAHGKK | 2.6 | NO | βHb |
| 950.506 | 950.509 | AAVTAFWGK | 3.2 | YES | βHb |
| 1071.554 | 1071.554 | MFLSFPTTK | 0 | NO | αHb |
| 1101.627 | 1101.629 | VLSAADKGNVK | −1.8 | YES | αHb |
| 1225.626 | 1225.625 | KVLDSFSNGMK | −0.8 | YES | βHb |
| 1274.724 | 1274.726 | LLVVYPWTQR | −1.6 | NO | βHb |
| 1328.715 | 1328.717 | VKVDEVGGEALGR | 1.5 | YES | βHb |
| 1477.795 | 1477.802 | VVAGVANALAHRYH | 4.7 | NO | βHb |
| 1529.733 | 1529.734 | VGGHAAEYGAEALER | 0.7 | NO | αHb |
| 1752.900 | 1752.899 | MLTAEEKAAVTAFWGK | −0.6 | YES | βHb |
| 1833.890 | 1833.891 | TYFPHFDLSHGSAQVK | 0.5 | NO | αHb |
| 1868.961 | 1868.954 | NFGKEFTPVLQADFQK | −3.7 | YES | βHb |
| 2089.953 | 2089.953 | FFESFGDLSTADAVMNNPK | 0 | YES | βHb |
| 2284.129 | 2284.126 | TYFPHFDLSHGSAQVKGHGAK | −1.3 | YES | αHb |
| Experimental m/z | Theoretical m/z | Mass Accuracy (ppm) | Peptide Sequence | Proteotypic | Protein |
|---|---|---|---|---|---|
| 767.487 | 767.487 | 0 | VKAHGKK | NO | βHb |
| 1041.542 | 1041.544 | 1.9 | MFLGFPTTK | YES | αHb |
| 1115. 643 | 1115.642 | −0.9 | VLSAADKANVK | YES | αHb |
| 1238.680 | 1238.685 | 4.0 | AHGQKVADALTK | YES | αHb |
| 1243.679 | 1243.679 | 0 | YELDKAFSDR | YES | αHb |
| 1265.826 | 1265.830 | 3.2 | LLGNVIVVVLAR | NO | αHb |
| 1274.724 | 1274.726 | 1.6 | LLVVYPWTQR | NO | βHb |
| 1314.670 | 1314.665 | −3.8 | VNVDEVGGEALGR | NO | βHb |
| 1422.700 | 1422.708 | 5.6 | VGGQAGAHGAEALER | YES | αHb |
| 1449.784 | 1449.796 | 8.3 | VVAGVANALAHKYH | NO | βHb |
| 1628.908 | 1628.912 | 2.5 | VLSAADKANVKAAWGK | YES | αHb |
| 1813.977 | 1813.981 | 2.2 | VLQSFSDGLKHLDNLK | YES | βHb |
| 1876.897 | 1876.898 | 0.5 | TYFPHFNLSHGSDQVK | YES | βHb |
| 1935.967 | 1935.978 | 5.7 | AAWGKVGGQAGAHGAEALER | YES | βHb |
| 2045.920 | 2045.927 | 3.4 | FFESFGDLSNADAVMGNPK | YES | βHb |
| 2237.156 | 2237.167 | 4.9 | AVGHLDDLPGALSALSDLHAHK | YES | βHb |
| 2318.244 | 2318.250 | 2.6 | VLQSFSDGLKHLDNLKGTFA K | YES | βHb |
| 2398.167 | 2398.167 | 0 | TYFPHFNLSHGSDQVKAHGQK | YES | αHb |
| 2445.216 | 2445.234 | 7.4 | VGGQAGAHGAEALERMFLGFPTTK | YES | αHb |
| Experimental m/z |
Theoretical m/z | Putative Peptide Match | Mass Accuracy (ppm) | Proteotypic | Protein |
|---|---|---|---|---|---|
| 920.489 | 920.495 | LSDLHAHK | 6.5 | YES | αHb |
| 1036.560 | 1036.567 | VLTSFGDAVK | 6.8 | YES | βHb |
| 1085.533 | 1085.534 | MFTTYPPTK | 0.9 | YES | βHb |
| 1288.728 | 1288.741 | LLIVYPWTQR | 10.1 | YES | βHb |
| 1302.635 | 1302.647 | VNVAECGAEALAR | 9.2 | YES | βHb |
| 1645.778 | 1645.782 | IAGHAEEYGAETLER | 2.4 | YES | αHb |
| 1704.959 | 1704.964 | VLSAADKNNVKGIFTK | 2.9 | YES | αHb |
| 2121.124 | 2121.155 | VVAALIEAANHIDDIAGTLSK | 14.6 | YES | αHb |
| 2226.142 | 2226.138 | FFASFGNLSSPTAILGNPMVR | −1.8 | YES | βHb |
| Species | Experimental m/z (Th) | Putative ID and UniProt Accession No. | Mass Accuracy (ppm) | MS/MS ID (and Accession No.) | Peptide Sequence |
|---|---|---|---|---|---|
| Bovine | 1198.718 | Actin (P62739) | −10.4 | Actin (P62739) | AVFPSIVGRPR |
| Bovine | 1348.768 | Carbonic anhydrase 3 (Q3SZX4) | −6.2 | Carbonic anhydrase 3 (Q3SZX4) | NWRPPQPIKGR |
| Bovine | 1669.851 | Myoglobin (P02192) | −8.6 | Myoglobin (P02192) | ALELFRNDMAAQYK |
| Bovine | 1771.932 | NI | −4.3 | Carbonic anhydrase 3 (Q3SZX4) | TLYSSAENEPPVPLVR |
| Bovine | 1790.916 | Actin (P62739) | −13.5 | Actin (P62739) | SYELPDGQVITIGNER |
| Bovine | 2280.178 | Myoglobin (P02192) | −8.2 | Myoglobin (P02192) | ALELFRNDMAAQYKVLGFHG |
| Porcine | 758.576 | NI | (Lipid) | NA | |
| Porcine | 796.533 | NI | (Lipid) | NA | |
| Porcine | 1536.823 | NI | NI | NA | |
| Porcine | 1541.766 | Beta-Enolase (Q1KYT0) | −1.2 | Beta-Enolase (Q1KYT0) | LAQSNGWGVMVSHR |
| Porcine | 2458.314 | NI | NI | NA | |
| Porcine | 2463.248 | NI | −0.3 | Beta-Enolase (Q1KYT0) | AAVPSGASTGIYEALELRDG DKSR |
| Porcine | 2123.125 | NI | Fructose-bisphosphate aldolase (Q6UV40) | IGEHTPSSLAIMENANVLAR | |
| Chicken | 1314.710 | NI | NI | NA | |
| Chicken | 1749.799 | GAPDH (P00356) | −7.0 | GAPDH (P00356) | LVSWYDNEFGYSNR |
| Chicken | 1936.042 | NI | NI | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kennedy, K.; Cole, L.; Witt, M.; Sealey, M.; Francese, S. Forensic Discrimination of Differentially Sourced Animal Blood Using a Bottom-Up Proteomics Based MALDI MS Approach. Molecules 2022, 27, 2039. https://doi.org/10.3390/molecules27072039
Kennedy K, Cole L, Witt M, Sealey M, Francese S. Forensic Discrimination of Differentially Sourced Animal Blood Using a Bottom-Up Proteomics Based MALDI MS Approach. Molecules. 2022; 27(7):2039. https://doi.org/10.3390/molecules27072039
Chicago/Turabian StyleKennedy, Katie, Laura Cole, Matthias Witt, Mark Sealey, and Simona Francese. 2022. "Forensic Discrimination of Differentially Sourced Animal Blood Using a Bottom-Up Proteomics Based MALDI MS Approach" Molecules 27, no. 7: 2039. https://doi.org/10.3390/molecules27072039
APA StyleKennedy, K., Cole, L., Witt, M., Sealey, M., & Francese, S. (2022). Forensic Discrimination of Differentially Sourced Animal Blood Using a Bottom-Up Proteomics Based MALDI MS Approach. Molecules, 27(7), 2039. https://doi.org/10.3390/molecules27072039