
Flavonoids in Treatment of Chronic Kidney Disease


Abstract
:1. Introduction
2. Diagnosis and Staging of CKD
3. Pathophysiology of CKD
4. Flavonoids
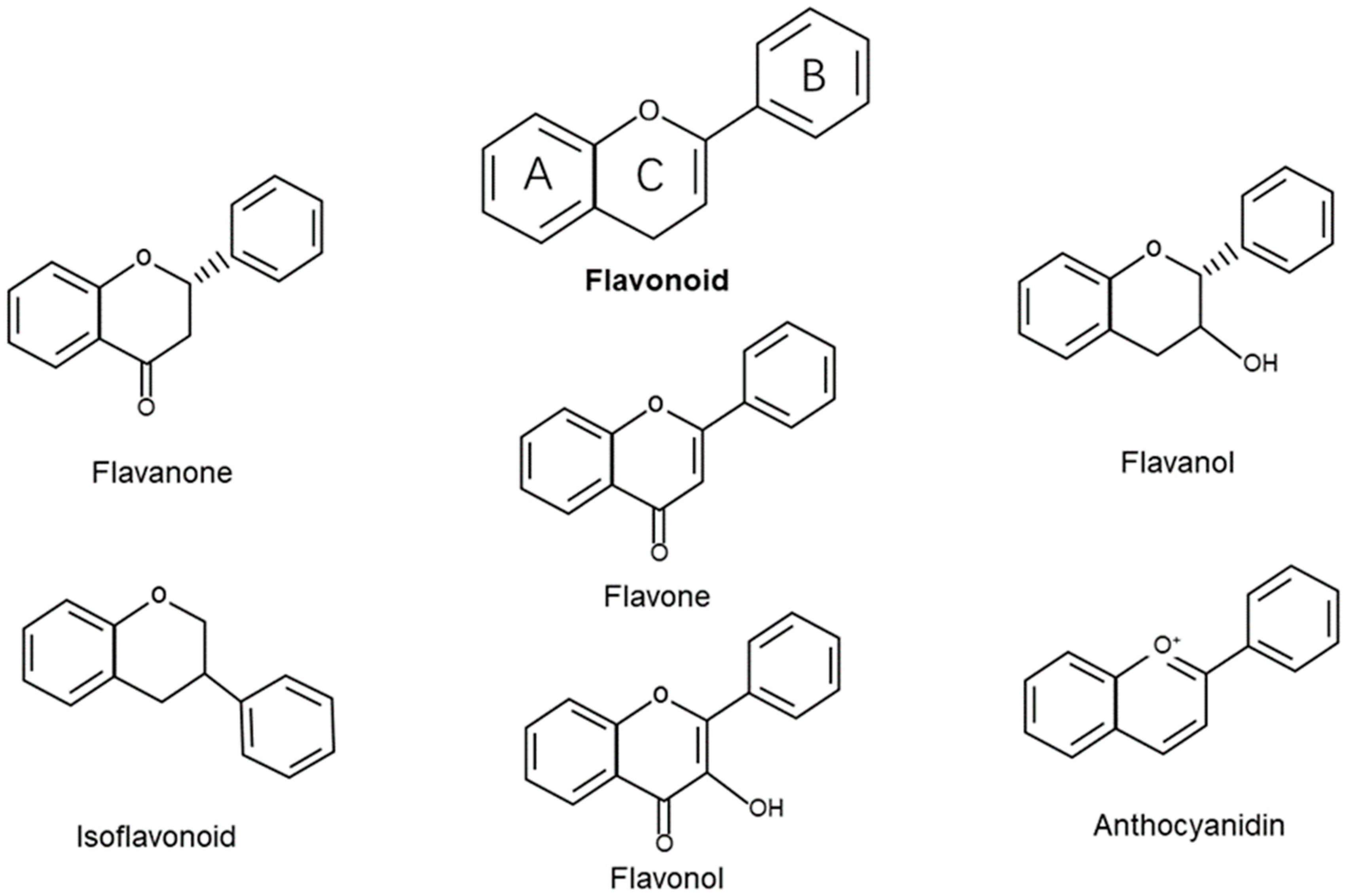
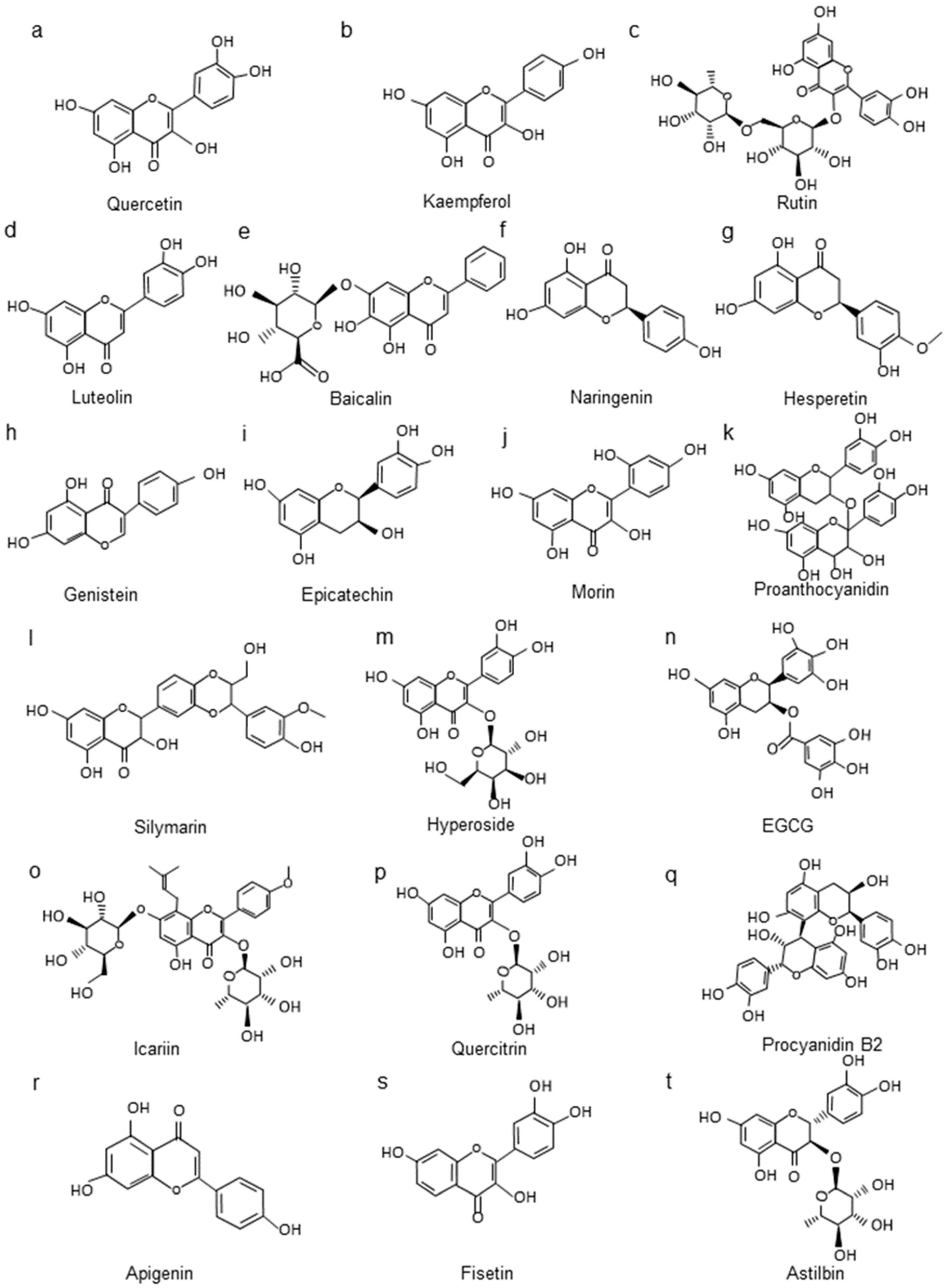
4.1. Structure and Classification of Flavonoids
4.2. Metabolism of Flavonoid
5. Bioactivities of Flavonoids in CKD
5.1. Antidiabetic Effect
5.2. Antihypertensive Effects
5.3. Anti-Inflammatory Effects
5.4. Antioxidant Effects
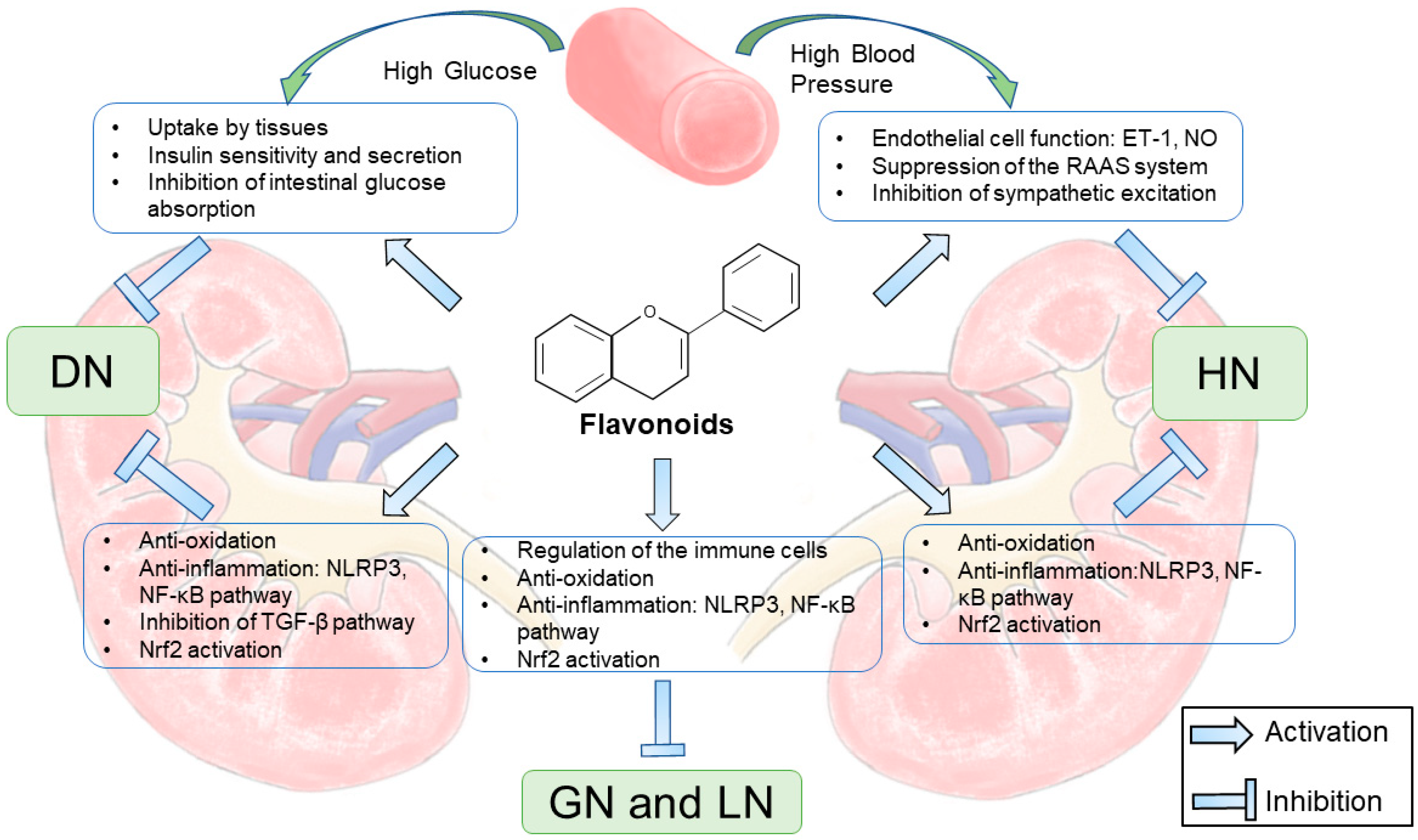
6. Flavonoids in CKD
6.1. Flavonoids in Diabetic Nephropathy

{kind=link}
{kind=link}
{kind=link}
| Animal Models | Flavonoids | Functions | References |
|---|---|---|---|
| STZ-induced DN rats | Quercetin | Increasing SOD and CAT activity; suppressing ROS and the NLRP3 inflammasome; scavenging free radicals; inhibiting TGF-β1 and CTGF | [144,145,146,147] |
| Kaempferol | Upregulating the Nrf2/HO-1 axis | [151] | |
| Baicalin | Downregulating PI3K/Akt/mTOR signaling | [159] | |
| Rutin | Inhibiting TGF-β1/Smad/ECM and TGF-β1/CTGF/ECM signaling pathways; influencing MMPs | [152,153] | |
| Luteolin | Upregulating Nphs2; increasing SOD/HO-1 and decreasing MDA | [157,158] | |
| Naringenin | Downregulating TGF-β1 and IL-1β; downregulating ER stress markers ATF4, p-PERK, p-eIF2α, and XBP1s | [160,161] | |
| Hesperidin | Restoring the α-Klotho/FGF-23 pathway; activating the Nrf2/ARE pathway | [162,163] | |
| STZ-induced DN mice | Kaempferol | Regulating TRAF6; inhibiting RhoA/Rho Kinase | [149,150] |
| Baicalin | Restoring Klotho expression and inhibiting Klotho hypermethylation | [164] | |
| Genistein | Reducing phospho-ERK/ERK ratio | [165] | |
| Alloxan-induced DN rats | Rutin | Regulating AQP2/AQP3/V2R genes; downregulating TGF-β1, GRP78, and CHOP | [154,155] |
| db/db mice | Quercetin | Reactivating the Hippo pathway | [148] |
| Baicalin | Activating Nrf2 and inhibiting the MAPK-mediated inflammatory signaling pathway | [166] | |
| Luteolin | Repressing STAT3 activation | [156] |
6.2. Flavonoids in Hypertensive Nephropathy
6.3. Flavonoids in Glomerulonephritis
| Animal Models | Flavonoids | Functions | References |
|---|---|---|---|
| Adriamycin-induced rat nephropathy | Baicalin | Suppressing the Notch1-Snail pathway | [184] |
| Total flavonoids in Astragali Radix | Regulating the immune system and RAAS | [185] | |
| Silymarin | Suppressing renal MDA and GSH depletion | [186] | |
| Adriamycin-induced mouse nephropathy | Hyperoside | Regulating mitochondrial fission by restoring the expression of Mfn-1 | [187] |
| Epigallocatechin-3- gallate | Suppressing oxidant stress and cell apoptosis; inhibiting the HIF-1α/ANGPTL4 pathway | [189] | |
| Anti-GBM-GN in 129/svJ mice | Epigallocatechin-3- gallate | Inhibiting MAPK pathways and phosphorylation of ERK1/2, JNK, and p38 | [188] |
| Bovine gamma-globulin-induced rat IgA nephropathy | Icariin | Inhibiting the NF-κB pathway and mediating NLRP3 inflammasome activation | [190] |
6.4. Flavonoids in Lupus Nephritis
7. Prospects and Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Kumari, A.; Karnatak, M.; Singh, D.; Shankar, R.; Jat, J.L.; Sharma, S.; Yadav, D.; Shrivastava, R.; Verma, V.P. Current scenario of artemisinin and its analogues for antimalarial activity. Eur. J. Med. Chem. 2019, 163, 804–829. [Google Scholar] [CrossRef] [PubMed]
- Kingston, D.G.I.; Cassera, M.B. Antimalarial Natural Products. Prog. Chem. Org. Nat. Prod. 2022, 117, 1–106. [Google Scholar] [PubMed]
- Habtemariam, S. Berberine pharmacology and the gut microbiota: A hidden therapeutic link. Pharmacol. Res. 2020, 155, 104722. [Google Scholar] [CrossRef] [PubMed]
- Jha, V.; Garcia-Garcia, G.; Iseki, K.; Li, Z.; Naicker, S.; Plattner, B.; Saran, R.; Wang, A.Y.-M.; Yang, C.-W. Chronic kidney disease: Global dimension and perspectives. Lancet 2013, 382, 260–272. [Google Scholar] [CrossRef]
- Coresh, J.; Selvin, E.; Stevens, L.A.; Manzi, J.; Kusek, J.W.; Eggers, P.; Van Lente, F.; Levey, A.S. Prevalence of Chronic Kidney Disease in the United States. JAMA: J. Am. Med Assoc. 2007, 298, 2038–2047. [Google Scholar] [CrossRef] [Green Version]
- Levin, A.; Tonelli, M.; Bonventre, J.; Coresh, J.; Donner, J.-A.; Fogo, A.B.; Fox, C.S.; Gansevoort, R.T.; Heerspink, H.J.L.; Jardine, M.; et al. Global kidney health 2017 and beyond: A roadmap for closing gaps in care, research, and policy. Lancet 2017, 390, 1888–1917. [Google Scholar] [CrossRef]
- Stevens, P.E.; Levin, A. Kidney Disease: Improving Global Outcomes Chronic Kidney Disease Guideline Development Work Group, Evaluation and management of chronic kidney disease: Synopsis of the kidney disease: Improving global outcomes 2012 clinical practice guideline. Ann. Intern. Med. 2013, 158, 825–830. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Bowe, B.; Mokdad, A.H.; Xian, H.; Yan, Y.; Li, T.; Maddukuri, G.; Tsai, C.-Y.; Floyd, T.; Al-Aly, Z. Analysis of the Global Burden of Disease study highlights the global, regional, and national trends of chronic kidney disease epidemiology from 1990 to 2016. Kidney Int. 2018, 94, 567–581. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, K.; Coresh, J.; Sang, Y.; Chalmers, J.; Fox, C.; Guallar, E.; Jafar, T.; Jassal, S.K.; Landman, G.W.D.; Muntner, P.; et al. Estimated glomerular filtration rate and albuminuria for prediction of cardiovascular outcomes: A collaborative meta-analysis of individual participant data. Lancet Diabetes Endocrinol. 2015, 3, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Gansevoort, R.T.; Matsushita, K.; van der Velde, M.; Astor, B.C.; Woodward, M.; Levey, A.S.; de Jong, P.E.; Coresh, J. Lower estimated GFR and higher albuminuria are associated with adverse kidney outcomes. A collaborative meta-analysis of general and high-risk population cohorts. Kidney Int. 2011, 80, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Vanholder, R.; Lameire, N.; Annemans, L.; Van Biesen, W. Cost of renal replacement: How to help as many as possible while keeping expenses reasonable? Nephrol. Dial. Transplant. 2015, 31, 1251–1261. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.-Q.; Hu, H.-H.; Wang, Y.-N.; Feng, Y.-L.; Cao, G.; Zhao, Y.-Y. Natural products for the prevention and treatment of kidney disease. Phytomedicine 2018, 50, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Bao, N.; Chen, F.; Dai, D. The Regulation of Host Intestinal Microbiota by Polyphenols in the Development and Prevention of Chronic Kidney Disease. Front. Immunol. 2020, 10, 2981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamsi, A.; Anwar, S.; Shahbaaz, M.; Mohammad, T.; Alajmi, M.F.; Hussain, A.; Hassan, I.; Ahmad, F.; Islam, A. Evaluation of Binding of Rosmarinic Acid with Human Transferrin and Its Impact on the Protein Structure: Targeting Polyphenolic Acid-Induced Protection of Neurodegenerative Disorders. Oxidative Med. Cell. Longev. 2020, 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Shamsi, A.; Shahbaaz, M.; Queen, A.; Khan, P.; Hasan, G.M.; Islam, A.; Alajmi, M.F.; Hussain, A.; Ahmad, F.; et al. Rosmarinic Acid Exhibits Anticancer Effects via MARK4 Inhibition. Sci. Rep. 2020, 10, 10300. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.-S. Regulatory Roles of Flavonoids on Inflammasome Activation during Inflammatory Responses. Mol. Nutr. Food Res. 2018, 62, e1800147. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croft, K.D. The chemistry and biological effects of flavonoids and phenolic acids. Ann. N. Y. Acad. Sci. 1998, 854, 435–442. [Google Scholar] [CrossRef]
- Abdou, H.M.; Elkader, H.-T.A.E.A. The potential therapeutic effects of Trifolium alexandrinum extract, hesperetin and quercetin against diabetic nephropathy via attenuation of oxidative stress, inflammation, GSK-3β and apoptosis in male rats. Chem. Interact. 2021, 352, 109781. [Google Scholar] [CrossRef]
- Jin, Y.; Zhai, Z.; Jia, H.; Lai, J.; Si, X.; Wu, Z. Kaempferol attenuates diquat-induced oxidative damage and apoptosis in intestinal porcine epithelial cells. Food Funct. 2021, 12, 6889–6899. [Google Scholar] [CrossRef]
- Mhatre, S.; Srivastava, T.; Naik, S.; Patravale, V. Antiviral activity of green tea and black tea polyphenols in prophylaxis and treatment of COVID-19: A review. Phytomedicine 2021, 85, 153286. [Google Scholar] [CrossRef] [PubMed]
- Jing, S.; Kong, X.; Wang, L.; Wang, H.; Feng, J.; Wei, L.; Meng, Y.; Liu, C.; Chang, X.; Qu, Y.; et al. Quercetin Reduces the Virulence of S. aureus by Targeting Clp, P to Protect Mice from MRSA-Induced Lethal Pneumonia. Microbiol. Spectr. 2022, e02340-21. [Google Scholar] [CrossRef] [PubMed]
- Scicchitano, M.; Carresi, C.; Nucera, S.; Ruga, S.; Maiuolo, J.; Macrì, R.; Scarano, F.; Bosco, F.; Mollace, R.; Cardamone, A.; et al. Icariin Protects H9c2 Rat Cardiomyoblasts from Doxorubicin-Induced Cardiotoxicity: Role of Caveolin-1 Upregulation and Enhanced Autophagic Response. Nutrients 2021, 13, 4070. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, X.; He, Q.; Zhang, M.; Chu, L.; Zhao, Y.; Wu, Y.; Zhang, J.; Han, X.; Chu, X.; et al. Investigation of the ameliorative effects of baicalin against arsenic trioxide-induced cardiac toxicity in mice. Int. Immunopharmacol. 2021, 99, 108024. [Google Scholar] [CrossRef]
- Alam, M.; Ali, S.; Ashraf, G.M.; Bilgrami, A.L.; Yadav, D.K.; Hassan, I. Epigallocatechin 3-gallate: From green tea to cancer therapeutics. Food Chem. 2022, 379, 132135. [Google Scholar] [CrossRef]
- Xiao, X.; Hu, Q.; Deng, X.; Shi, K.; Zhang, W.; Jiang, Y.; Ma, X.; Zeng, J.; Wang, X. Old wine in new bottles: Kaempferol is a promising agent for treating the trilogy of liver diseases. Pharmacol. Res. 2021, 175, 106005. [Google Scholar] [CrossRef]
- Suarez, N.G.; Fernandez-Marrero, Y.; Torabidastgerdooei, S.; Annabi, B. EGCG Prevents the Onset of an Inflam-matory and Cancer-Associated Adipocyte-like Phenotype in Adipose-Derived Mesenchymal Stem/Stromal Cells in Response to the Triple-Negative Breast Cancer Secretome. Nutrients 2022, 14, 1099. [Google Scholar] [CrossRef]
- Vargas, F.; Romecín, P.; Guillen, A.I.G.; Wangesteen, R.; Vargas-Tendero, P.; Paredes, M.D.; Atucha, N.M.; García-Estañ, J. Flavonoids in Kidney Health and Disease. Front. Physiol. 2018, 9, 394. [Google Scholar] [CrossRef] [Green Version]
- Letourneau, P.; Bataille, S.; Chauveau, P.; Fouque, D.; Koppe, L. Source and Composition in Amino Acid of Dietary Proteins in the Primary Prevention and Treatment of CKD. Nutrients 2020, 12, 3892. [Google Scholar] [CrossRef]
- Vanholder, R.; Baurmeister, U.; Brunet, P.; Cohen, G.; Glorieux, G.; Jankowski, J. A Bench to Bedside View of Uremic Toxins. J. Am. Soc. Nephrol. 2008, 19, 863–870. [Google Scholar] [CrossRef] [Green Version]
- Levey, A.S.; Becker, C.; Inker, L.A. Glomerular filtration rate and albuminuria for detection and staging of acute and chronic kidney disease in adults: A systematic review. JAMA 2015, 313, 837–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.K.; Knicely, D.H.; Grams, M.E. Chronic Kidney Disease Diagnosis and Management. JAMA J. Am. Med. Assoc. 2019, 322, 1294–1304. [Google Scholar] [CrossRef] [PubMed]
- Charles, C.; Ferris, A.H. Chronic Kidney Disease. Prim. Care 2020, 47, 585–595. [Google Scholar] [CrossRef]
- Webster, A.C.; Nagler, E.V.; Morton, R.L.; Masson, P. Chronic Kidney Disease. Lancet 2017, 389, 1238–1252. [Google Scholar] [CrossRef]
- Nagata, M. Podocyte injury and its consequences. Kidney Int. 2016, 89, 1221–1230. [Google Scholar] [CrossRef]
- Zhao, X.-C.; Livingston, M.J.; Liang, X.-L.; Dong, Z. Cell Apoptosis and Autophagy in Renal Fibrosis. Adv. Pain Res. Mech. Modul. Chronic Pain 2019, 1165, 557–584. [Google Scholar]
- Sol, M.; Kamps, J.A.A.M.; Born, J.V.D.; Heuvel, M.C.V.D.; Van Der Vlag, J.; Krenning, G.; Hillebrands, J.-L. Glomerular Endothelial Cells as Instigators of Glomerular Sclerotic Diseases. Front. Pharmacol. 2020, 11, 573557. [Google Scholar] [CrossRef]
- Schöcklmann, H.O.; Lang, S.; Sterzel, R.B. Regulation of mesangial cell proliferation. Kidney Int. 1999, 56, 1199–1207. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.H. Mesangial Cells and Renal Fibrosis. Adv. Exp. Med. Biol. 2019, 1165, 165–194. [Google Scholar]
- Shankland, S. The podocyte’s response to injury: Role in proteinuria and glomerulosclerosis. Kidney Int. 2006, 69, 2131–2147. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.-C.; Wang, G.-H.; Lu, J.; Chen, P.-P.; Zhang, Y.; Hu, Z.-B.; Ma, K.-L. Role of Podocyte Injury in Glomerulosclerosis. Adv. Exp. Med. Biol. 2019, 1165, 195–232. [Google Scholar] [PubMed]
- Schelling, J.R. Tubular atrophy in the pathogenesis of chronic kidney disease progression. Pediatr. Nephrol. 2016, 31, 693–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.-C.; Tang, T.-T.; Lv, L.-L. How Tubular Epithelial Cell Injury Contributes to Renal Fibrosis. Sunlight Vitam. D Skin Cancer 2019, 1165, 233–252. [Google Scholar]
- Tucker, P.S.; Scanlan, A.; Dalbo, V.J. Chronic Kidney Disease Influences Multiple Systems: Describing the Relationship between Oxidative Stress, Inflammation, Kidney Damage, and Concomitant Disease. Oxidative Med. Cell. Longev. 2015, 2015, 806358. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Luo, K.; Liu, H.; Huang, T.; Fang, X.; Tu, W. The progress of inflammation and oxidative stress in patients with chronic kidney disease. Ren. Fail. 2014, 37, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.; Booz, G.W.; Wang, Y.; Fan, F.; Roman, R.J. Inflammation and renal fibrosis: Recent developments on key signaling molecules as potential therapeutic targets. Eur. J. Pharmacol. 2018, 820, 65–76. [Google Scholar] [CrossRef]
- Duffield, J.S. Cellular and molecular mechanisms in kidney fibrosis. J. Clin. Investig. 2014, 124, 2299–2306. [Google Scholar] [CrossRef] [Green Version]
- Djudjaj, S.; Boor, P. Cellular and molecular mechanisms of kidney fibrosis. Mol. Asp. Med. 2019, 65, 16–36. [Google Scholar] [CrossRef]
- Kozłowska, A.; Szostak-Wegierek, D. Flavonoids--food sources and health benefits. Rocz. Państwowego Zakładu Hig. 2014, 65, 79–85. [Google Scholar]
- Pinto, C.; Cidade, H.; Pinto, M.; Tiritan, M.E. Chiral Flavonoids as Antitumor Agents. Pharmaceuticals 2021, 14, 1267. [Google Scholar] [CrossRef]
- Ren, F.; Nian, Y.; Perussello, C.A. Effect of storage, food processing and novel extraction technologies on onions flavonoid content: A review. Food Res. Int. 2020, 132, 108953. [Google Scholar] [CrossRef] [PubMed]
- Janabi, A.H.W.; Kamboh, A.A.; Saeed, M.; Xiaoyu, L.; Bi, J.; Majeed, F.; Naveed, M.; Mughal, M.J.; Korejo, N.A.; Kamboh, R.; et al. Flavonoid-rich foods (FRF): A promising nutraceutical approach against lifespan-shortening diseases. Iran J. Basic Med. Sci. 2020, 23, 140–153. [Google Scholar]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.; et al. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnol. Adv. 2020, 38, 107316. [Google Scholar] [CrossRef]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Siah, M.; Farzaei, M.H.; Ashrafi-Kooshk, M.R.; Adibi, H.; Arab, S.S.; Rashidi, M.R.; Khodarahmi, R. Inhibition of guinea pig aldehyde oxidase activity by different flavonoid compounds: An in vitro study. Bioorg. Chem. 2016, 64, 74–84. [Google Scholar] [CrossRef]
- Brunetti, C.; Di Ferdinando, M.; Fini, A.; Pollastri, S.; Tattini, M. Flavonoids as Antioxidants and Developmental Regulators: Relative Significance in Plants and Humans. Int. J. Mol. Sci. 2013, 14, 3540–3555. [Google Scholar] [CrossRef] [Green Version]
- Ciumărnean, L.; Milaciu, M.V.; Runcan, O.; Vesa, S.C.; Răchișan, A.L.; Negrean, V.; Perné, M.-G.; Donca, V.I.; Alexescu, T.-G.; Para, I.; et al. The effects of favonoids in cardiovascular diseases. Molecules 2020, 25, 4320. [Google Scholar] [CrossRef]
- Pan, K.S.; Cho, I.; Yi, C.H.; To, D.F.; Chen, C.W. Do, Baicalein, Baicalin, and Wogonin: Protective Effects against Ische-mia-Induced Neurodegeneration in the Brain and Retina. Oxid Med. Cell Longev 2021, 2021, 8377362. [Google Scholar] [CrossRef]
- Sun, X.Y.; Li, L.J.; Dong, Q.X.; Zhu, J.; Huang, Y.R.; Hou, S.J.; Yu, X.L.; Liu, R.T. Rutin prevents tau pathology and neuroin-flammation in a mouse model of Alzheimer’s disease. J. Neuroinflamm. 2021, 18, 131. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as anticancer agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousuf, M.; Shamsi, A.; Queen, A.; Shahbaaz, M.; Khan, P.; Hussain, A.; Alajmi, M.F.; Haque, Q.M.R.; Hassan, I. Targeting cyclin-dependent kinase 6 by vanillin inhibits proliferation of breast and lung cancer cells: Combined computational and biochemical studies. J. Cell. Biochem. 2021, 122, 897–910. [Google Scholar] [CrossRef]
- Jiang, Z.-B.; Wang, W.-J.; Xu, C.; Xie, Y.-J.; Wang, X.-R.; Zhang, Y.-Z.; Huang, J.-M.; Huang, M.; Xie, C.; Liu, P.; et al. Luteolin and its derivative apigenin suppress the inducible PD-L1 expression to improve anti-tumor immunity in KRAS-mutant lung cancer. Cancer Lett. 2021, 515, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Minihane, A.M. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzales, G.; Smagghe, C.; Grootaert, M.; Zotti, K.; Raes, J. Van Camp, Flavonoid interactions during digestion, ab-sorption, distribution and metabolism: A sequential structure-activity/property relationship-based approach in the study of bioavailability and bioactivity. Drug Metab. Rev. 2015, 47, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Jakobek, L. Interactions of polyphenols with carbohydrates, lipids and proteins. Food Chem. 2015, 175, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yang, J.; Xie, Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. Int. J. Pharm. 2019, 570, 118642. [Google Scholar] [CrossRef]
- Akkati, D.; Sam, K.G.; Tungha, G. Emergence of promising therapies in diabetes mellitus. J. Clin. Pharmacol. 2011, 51, 796–804. [Google Scholar] [CrossRef]
- Babu, P.V.A.; Liu, D.; Gilbert, E.R. Recent advances in understanding the anti-diabetic actions of dietary flavonoids. J. Nutr. Biochem. 2013, 24, 1777–1789. [Google Scholar] [CrossRef] [Green Version]
- Shahwan, M.; Alhumaydhi, F.; Ashraf, G.M.; Hasan, P.M.; Shamsi, A. Role of polyphenols in combating Type 2 Diabetes and insulin resistance. Int. J. Biol. Macromol. 2022, 206, 567–579. [Google Scholar] [CrossRef]
- Haddad, P.S.; Eid, H.M. The Antidiabetic Potential of Quercetin: Underlying Mechanisms. Curr. Med. Chem. 2017, 24, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Gu, Y.; Zhang, Q.; Liu, L.; Meng, G.; Wu, H.; Xia, Y.; Bao, X.; Shi, H.; Sun, S.; et al. Estimated daily quercetin intake and association with the prevalence of type 2 diabetes mellitus in Chinese adults. Z. für Ernähr. 2018, 58, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Hu, J.; Zhao, W.; Gao, X.; Jiang, C.; Liu, K.; Liu, B.; Huang, F. Quercetin differently regulates insulin-mediated glucose transporter 4 translocation under basal and inflammatory conditions in adipocytes. Mol. Nutr. Food Res. 2013, 58, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Eid, H.M.; Martineau, L.C.; Saleem, A.; Muhammad, A.; Vallerand, D.; Benhaddou-Andaloussi, A.; Nistor, L.; Afshar, A.; Arnason, J.T.; Haddad, P.S. Stimulation of AMP-activated protein kinase and enhancement of basal glucose uptake in muscle cells by quercetin and quercetin glycosides, active principles of the antidiabetic medicinal plant Vaccinium vitis-idaea. Mol. Nutr. Food Res. 2010, 54, 991–1003. [Google Scholar] [CrossRef]
- He, L.; Zhang, N.; Wang, L.; Du, L.; Li, C.; Li, Y.; Li, X.; Zhu, X.; Lu, Q.; Yin, X. Quercetin Inhibits AQP1 Translocation in High-Glucose-Cultured SRA01/04 Cells Through PI3K/Akt/m, TOR Pathway. Curr. Mol. Pharmacol. 2021, 14, 587–596. [Google Scholar] [CrossRef]
- Li, X.; Wang, R.; Zhou, N.; Wang, X.; Liu, Q.; Bai, Y.; Bai, Y.; Liu, Z.; Yang, H.; Zou, J.; et al. Quercetin improves insulin resistance and hepatic lipid accumulation in vitro in a NAFLD cell model. Biomed. Rep. 2013, 1, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Tzeng, Y.-M.; Chen, K.; Rao, Y.K.; Lee, M.-J. Kaempferitrin activates the insulin signaling pathway and stimulates secretion of adiponectin in 3T3-L1 adipocytes. Eur. J. Pharmacol. 2009, 607, 27–34. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, D. Flavonol kaempferol improves chronic hyperglycemia-impaired pancreatic beta-cell viability and insulin secretory function. Eur. J. Pharmacol. 2011, 670, 325–332. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Q.; Liang, J.; Dai, X.; Ding, Y.; Wang, J.; Li, Y. Epigallocatechin-3-O-gallate (EGCG) protects the insulin sensitivity in rat L6 muscle cells exposed to dexamethasone condition. Phytomedicine 2010, 17, 14–18. [Google Scholar] [CrossRef]
- Ha, B.G.; Nagaoka, M.; Yonezawa, T.; Tanabe, R.; Woo, J.T.; Kato, H.; Chung, U.-I.; Yagasaki, K. Regulatory mechanism for the stimulatory action of genistein on glucose uptake in vitro and in vivo. J. Nutr. Biochem. 2012, 23, 501–509. [Google Scholar] [CrossRef]
- Tzeng, T.F.; Liou, S.S.; Liu, I.M. Myricetin Ameliorates Defective Post-Receptor Insulin Signaling via beta-Endorphin Signaling in the Skeletal Muscles of Fructose-Fed Rats. Evid-Based Compl. Alt. 2011, 2011, 150752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, I.-M.; Tzeng, T.-F.; Liou, S.-S.; Lan, T.-W. Myricetin, a naturally occurring flavonol, ameliorates insulin resistance induced by a high-fructose diet in rats. Life Sci. 2007, 81, 1479–1488. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Zhou, F.C.; Gao, F.; Bian, J.S.; Shan, F. Comparative Evaluation of Quercetin, Isoquercetin and Rutin as Inhibitors of α-Glucosidase. J. Agric. Food Chem. 2009, 57, 11463–11468. [Google Scholar] [CrossRef] [PubMed]
- Kappel, V.D.; Cazarolli, L.H.; Pereira, D.F.; Postal, B.G.; Zamoner, A.; Reginatto, F.H.; Silva, F.R.M.B. Involvement of GLUT-4 in the stimulatory effect of rutin on glucose uptake in rat soleus muscle. J. Pharm. Pharmacol. 2013, 65, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Ola, M.S.; Ahmed, M.M.; Ahmad, R.; Abuohashish, H.M.; Al-Rejaie, S.S.; Alhomida, A.S. Neuroprotective Effects of Rutin in Streptozotocin-Induced Diabetic Rat Retina. J. Mol. Neurosci. 2015, 56, 440–448. [Google Scholar] [CrossRef]
- Houston, M.; Hays, L. Acute Effects of an Oral Nitric Oxide Supplement on Blood Pressure, Endothelial Function, and Vascular Compliance in Hypertensive Patients. J. Clin. Hypertens. 2014, 16, 524–529. [Google Scholar] [CrossRef] [Green Version]
- Puzserova, A.; Bernatova, I. Blood Pressure Regulation in Stress: Focus on Nitric Oxide-Dependent Mechanisms. Physiol. Res. 2016, 65, S309–S342. [Google Scholar] [CrossRef]
- Gao, Y.; Chen, T.; Raj, J.U. Endothelial and Smooth Muscle Cell Interactions in the Pathobiology of Pulmonary Hypertension. Am. J. Respir. Cell. Mol. Biol. 2016, 54, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Thamcharoen, N.; Susantitaphong, P.; Wongrakpanich, S.; Chongsathidkiet, P.; Tantrachoti, P.; Pitukweerakul, S.; Avihingsanon, Y.; Praditpornsilpa, K.; Jaber, B.L.; Eiam-Ong, S. Effect of N- and T-type calcium channel blocker on proteinuria, blood pressure and kidney function in hypertensive patients: A meta-analysis. Hypertens. Res. 2015, 38, 847–855. [Google Scholar] [CrossRef]
- Lin, X.; Han, T.; Fan, Y.; Wu, S.; Wang, F.; Wang, C. Quercetin improves vascular endothelial function through promotion of autophagy in hypertensive rats. Life Sci. 2020, 258, 118106. [Google Scholar] [CrossRef]
- Yamamoto, M.; Suzuki, A.; Hase, T. Short-term effects of glucosyl hesperidin and hesperetin on blood pressure and vascular endothelial function in spontaneously hypertensive rats. J. Nutr. Sci. Vitaminol. 2008, 54, 95–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, A.J.; Symons, J.D.; Jalili, T. Therapeutic Potential of Quercetin to Decrease Blood Pressure: Review of Efficacy and Mechanisms. Adv. Nutr. Int. Rev. J. 2012, 3, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunbupha, S.; Apaijit, K.; Potue, P.; Maneesai, P.; Pakdeechote, P. Hesperidin inhibits L-NAME-induced vascular and renal alterations in rats by suppressing the renin-angiotensin system, transforming growth factor-beta1, and oxidative stress. Clin. Exp. Pharmacol. Physiol. 2021, 48, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Oyagbemi, A.A.; Omobowale, T.O.; Ola-Davies, O.E.; Asenuga, E.R.; Ajibade, T.O.; Adejumobi, O.A.; Arojojoye, O.A.; Afolabi, J.M.; Ogunpolu, B.S.; Falayi, O.O.; et al. Quercetin attenuates hypertension induced by sodium fluoride via reduction in oxidative stress and modulation of HSP 70/ERK/PPARgamma signaling pathways. Biofactors 2018, 44, 465–479. [Google Scholar] [CrossRef]
- Olaleye, M.T.; Crown, O.; Akinmoladun, A.; Akindahunsi, A.A. Rutin and quercetin show greater efficacy than nifedipin in ameliorating hemodynamic, redox, and metabolite imbalances in sodium chloride-induced hypertensive rats. Hum. Exp. Toxicol. 2013, 33, 602–608. [Google Scholar] [CrossRef]
- Bruno, R.M.; Ghiadoni, L. Polyphenols, Antioxidants and the Sympathetic Nervous System. Curr. Pharm. Des. 2018, 24, 130–139. [Google Scholar] [CrossRef]
- Wunpathe, C.; Potue, P.; Maneesai, P.; Bunbupha, S.; Prachaney, P.; Kukongviriyapan, U.; Kukongviriyapan, V.; Pakdeechote, P. Hesperidin Suppresses Renin-Angiotensin System Mediated NOX2 Over-Expression and Sympathoexcitation in 2K-1C Hypertensive Rats. Am. J. Chin. Med. 2018, 46, 751–767. [Google Scholar] [CrossRef]
- Leeya, Y.; Mulvany, M.J.; Queiroz, E.F.; Marston, A.; Hostettmann, K.; Jansakul, C. Hypotensive activity of an n-butanol extract and their purified compounds from leaves of Phyllanthus acidus (L.) Skeels in rats. Eur. J. Pharmacol. 2010, 649, 301–313. [Google Scholar] [CrossRef]
- Rivera, L.; Morón, R.; Sánchez, M.; Zarzuelo, A.; Galisteo, M. Quercetin Ameliorates Metabolic Syndrome and Improves the Inflammatory Status in Obese Zucker Rats. Obesity 2008, 16, 2081–2087. [Google Scholar] [CrossRef]
- Campos-Toimil, M.; Orallo, F. Effects of (−)-epigallocatechin-3-gallate in Ca2+-permeable non-selective cation channels and voltage-operated Ca2+ channels in vascular smooth muscle cells. Life Sci. 2007, 80, 2147–2153. [Google Scholar] [CrossRef]
- Liu, P.L.; Liu, J.T.; Kuo, H.F.; Chong, I.W.; Hsieh, C.C. Epigallocatechin gallate attenuates proliferation and oxidative stress in human vascular smooth muscle cells induced by interleukin-1beta via heme oxygenase-1. Mediat. Inflamm. 2014, 2014, 523684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Niu, L.; Cui, L.; Hou, X.; Li, J.; Zhang, X.; Zhang, M. Hesperetin inhibits rat coronary constriction by inhibiting Ca2+ influx and enhancing voltage-gated K+ channel currents of the myocytes. Eur. J. Pharmacol. 2014, 735, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Jokura, H.; Hashizume, K.; Ominami, H.; Shibuya, Y.; Suzuki, A.; Hase, T.; Shimotoyodome, A. Hesperidin metabolite hesperetin-7-O-glucuronide, but not hesperetin-3′-O-glucuronide, exerts hypotensive, vasodilatory, and anti-inflammatory activities. Food Funct. 2013, 4, 1346–1351. [Google Scholar] [CrossRef] [PubMed]
- Mita, M.; Tanaka, H.; Yanagihara, H.; Nakagawa, J.; Hishinuma, S.; Sutherland, C.; Walsh, M.P.; Shoji, M. Membrane depo-larization-induced Rho, A/Rho-associated kinase activation and sustained contraction of rat caudal arterial smooth muscle involves genistein-sensitive tyrosine phosphorylation. J. Smooth Muscle Res. 2013, 49, 26–45. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Xu, H.-T.; Yu, J.-J.; Gao, J.; Lei, J.; Yin, Q.-S.; Li, B.; Pang, M.-X.; Su, M.-X.; Mi, W.-J.; et al. Luteolin Ameliorates Hypertensive Vascular Remodeling through Inhibiting the Proliferation and Migration of Vascular Smooth Muscle Cells. Evid.-Based Complement. Altern. Med. 2015, 2015, 364876. [Google Scholar] [CrossRef] [Green Version]
- Saponara, S.; Testai, L.; Iozzi, D.; Martinotti, E.; Martelli, A.; Chericoni, S.; Sgaragli, G.; Fusi, F.; Calderone, V. (+/−)-Naringenin as large conductance Ca(2+)-activated K+ (BKCa) channel opener in vascular smooth muscle cells. Br. J. Pharmacol. 2006, 149, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Zhou, Z.; Zhang, Q.; Cai, W.; Zhou, Y.; Sun, H.; Qiu, L. Vaccarin administration ameliorates hypertension and cardiovascular remodeling in renovascular hypertensive rats. J. Cell. Biochem. 2018, 119, 926–937. [Google Scholar] [CrossRef]
- Pan, M.-H.; Lai, C.-S.; Ho, C.-T. Anti-inflammatory activity of natural dietary flavonoids. Food Funct. 2010, 1, 15–31. [Google Scholar] [CrossRef]
- Shen, H.; Kreisel, D.; Goldstein, D.R. Processes of Sterile Inflammation. J. Immunol. 2013, 191, 2857–2863. [Google Scholar] [CrossRef]
- Lin, W.; Wang, W.; Wang, D.; Ling, W. Quercetin protects against atherosclerosis by inhibiting dendritic cell activation. Mol. Nutr. Food Res. 2017, 61, 1700031. [Google Scholar] [CrossRef]
- Weng, Z.; Zhang, B.; Asadi, S.; Sismanopoulos, N.; Butcher, A.; Fu, X.; Katsarou-Katsari, A.; Antoniou, C.; Theoharides, T.C. Quercetin Is More Effective than Cromolyn in Blocking Human Mast Cell Cytokine Release and Inhibits Contact Dermatitis and Photosensitivity in Humans. PLoS ONE 2012, 7, e33805. [Google Scholar] [CrossRef] [PubMed]
- Weng, Z.; Patel, A.B.; Panagiotidou, S.; Theoharides, T.C. The novel flavone tetramethoxyluteolin is a potent inhibitor of human mast cells. J. Allergy Clin. Immunol. 2015, 135, 1044–1052.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.H.; Shin, D.; Han, S.Y.; Kim, J.L.; Kang, Y.H. Kaempferol suppresses eosionphil infiltration and airway inflam-mation in airway epithelial cells and in mice with allergic asthma. J. Nutr. 2012, 142, 47–56. [Google Scholar] [CrossRef] [PubMed]
- del Bo, C.; Roursgaard, M.; Porrini, M.; Loft, S.; Moller, P.; Riso, P. Different effects of anthocyanins and phenolic acids from wild blueberry (Vaccinium angustifolium) on monocytes adhesion to endothelial cells in a TNF-alpha stimulated proin-flammatory environment. Mol. Nutr. Food Res. 2016, 60, 2355–2366. [Google Scholar] [CrossRef] [PubMed]
- Lolli, G.; Cozza, G.; Mazzorana, M.; Tibaldi, E.; Cesaro, L.; Donella-Deana, A.; Meggio, F.; Venerando, A.; Franchin, C.; Sarno, S.; et al. Inhibition of Protein Kinase CK2 by Flavonoids and Tyrphostins. A Structural Insight. Biochemistry 2012, 51, 6097–6107. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Kosaka, Y.; Mizuguchi, M. Structural Insight into the Interactions between Death-Associated Protein Kinase 1 and Natural Flavonoids. J. Med. Chem. 2015, 58, 7400–7408. [Google Scholar] [CrossRef]
- Parate, S.; Kumar, V.; Hong, J.; Lee, K. Identification of Flavonoids as Putative ROS-1 Kinase Inhibitors Using Pharmacophore Modeling for NSCLC Therapeutics. Molecules 2021, 26, 2114. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Belchor, M.N.; Gaeta, H.H.; Rodrigues, C.F.B.; Costa, C.R.D.C.; Toyama, D.D.O.; Passero, L.F.D.; Laurenti, M.D.; Toyama, M.H. Evaluation of Rhamnetin as an Inhibitor of the Pharmacological Effect of Secretory Phospholipase A. Molecules 2017, 22, 1441. [Google Scholar] [CrossRef] [Green Version]
- Hanáková, Z.; Hošek, J.; Kutil, Z.; Temml, V.; Landa, P.; Vaněk, T.; Schuster, D.; Dall’acqua, S.; Cvačka, J.; Polanský, O.; et al. Anti-inflammatory Activity of Natural Geranylated Flavonoids: Cyclooxygenase and Lipoxygenase Inhibitory Properties and Proteomic Analysis. J. Nat. Prod. 2017, 80, 999–1006. [Google Scholar] [CrossRef]
- Mosquera, D.M.G.; Ortega, Y.H.; Fernandez, P.L.; Gonzalez, Y.; Doens, D.; Heyden, Y.V.; Foubert, K.; Pieters, L. Flavonoids from Boldoa purpurascens inhibit proinflammatory cytokines (TNF-alpha and IL-6) and the expression of COX-2. Phytother. Res. 2018, 32, 1750–1754. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Men, W.; Shan, X.; Zhai, H.; Qiao, X.; Geng, L.; Li, C. Baicalein exerts neuroprotective effect against ischaem-ic/reperfusion injury via alteration of NF-k, B and LOX and AMPK/Nrf2 pathway. Inflammopharmacology 2020, 28, 1327–1341. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.L.; Huang, W.C.; Cheng, S.C.; Liou, C.J. Fisetin inhibits the generation of inflammatory mediators in interleu-kin-1beta-induced human lung epithelial cells by suppressing the NF-kappa, B and ERK1/2 pathways. Int. Immunopharmacol. 2018, 60, 202–210. [Google Scholar] [CrossRef]
- Ren, Q.; Guo, F.; Tao, S.; Huang, R.; Ma, L.; Fu, P. Flavonoid fisetin alleviates kidney inflammation and apoptosis via inhibiting Src-mediated NF-kappa, B p65 and MAPK signaling pathways in septic AKI mice. Biomed. Pharmacother. 2020, 122, 109772. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arts, I.C.W.; Sesink, A.L.A.; Faassen-Peters, M.; Hollman, P.C.H. The type of sugar moiety is a major determinant of the small intestinal uptake and subsequent biliary excretion of dietary quercetin glycosides. Br. J. Nutr. 2004, 91, 841–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, M.; Romero, M.; Gomez-Guzman, M.; Tamargo, J.; Perez-Vizcaino, F.; Duarte, J. Cardiovascular Effects of Flavonoids. Curr. Med. Chem. 2019, 26, 6991–7034. [Google Scholar] [CrossRef]
- Hamid, A.A.; Aminuddin, A.; Yunus, M.H.M.; Murthy, J.K.; Hui, C.K.; Ugusman, A. Antioxidative and anti-inflammatory activities of Polygonum minus: A review of literature. Rev. Cardiovasc. Med. 2020, 21, 275–287. [Google Scholar]
- Akhlaghi, M.; Bandy, B. Mechanisms of flavonoid protection against myocardial ischemia–reperfusion injury. J. Mol. Cell. Cardiol. 2009, 46, 309–317. [Google Scholar] [CrossRef]
- Kulczynski, B.; Kobus-Cisowska, J.; Kmiecik, D.; Gramza-Michalowska, A.; Golczak, D.; Korczak, J. Antiradical capacity and polyphenol composition of asparagus spears varieties cultivated under different sunlight conditions. Acta Sci. Pol. Technol. Aliment 2016, 15, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Kejík, Z.; Kaplánek, R.; Masařík, M.; Babula, P.; Matkowski, A.; Filipenský, P.; Veselá, K.; Gburek, J.; Sýkora, D.; Martásek, P.; et al. Iron Complexes of Flavonoids-Antioxidant Capacity and Beyond. Int. J. Mol. Sci. 2021, 22, 646. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Liu, L.; Guo, X.; Zhang, S.; Wang, J.; Zhou, F.; Tang, Y.; Yao, P. Quercetin attenuates high fat diet-induced atherosclerosis in apolipoprotein E knockout mice: A critical role of NADPH oxidase. Food Chem. Toxicol. 2017, 105, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Song, L. Quercetin protects human liver cells from o,p’-DDT-induced toxicity by suppressing Nrf2 and NADPH oxidase-regulated ROS production. Food Chem. Toxicol. 2022, 116, 112849. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Tian, R.; Yang, Z.; Peng, Y.-Y.; Lu, N. Quercetin suppressed NADPH oxidase-derived oxidative stress via heme oxygenase-1 induction in macrophages. Arch. Biochem. Biophys. 2019, 671, 69–76. [Google Scholar] [CrossRef]
- Shi, J.; Wu, G.; Zou, X.; Jiang, K. Enteral Baicalin, a Flavone Glycoside, Reduces Indicators of Cardiac Surgery-Associated Acute Kidney Injury in Rats. Cardiorenal Med. 2018, 9, 31–40. [Google Scholar] [CrossRef]
- Satta, S.; Mahmoud, A.M.; Wilkinson, F.L.; Yvonne Alexander, M.; White, S.J. The role of Nrf2 in cardiovascular function and disease. Oxid. Med. Cell. Longev. 2017, 2017, 9237263. [Google Scholar] [CrossRef]
- Qin, S.; Deng, F.; Wu, W.; Jiang, L.; Yamashiro, T.; Yano, S.; Hou, D.X. Baicalein modulates Nrf2/Keap1 system in both Keap1-dependent and Keap1-independent mechanisms. Arch. Biochem. Biophys. 2014, 559, 53–61. [Google Scholar] [CrossRef]
- Yuan-Jing, F.; Wei, W.; Jian-Ping, L.; Yu-Xia, J.; Zi-Ling, D. Genistein promotes the metabolic transformation of aceta-minophen to glucuronic acid in human L-O2, Hep, G2 and Hep3b cells via the Nrf2/Keap1 pathway. Food Funct. 2016, 7, 4683–4692. [Google Scholar] [CrossRef]
- Kapoor, R.; Sirohi, V.K.; Gupta, K.; Dwivedi, A. Naringenin ameliorates progression of endometriosis by modulating Nrf2/Keap1/HO1 axis and inducing apoptosis in rats. J. Nutr. Biochem. 2019, 70, 215–226. [Google Scholar] [CrossRef]
- Darband, S.G.; Sadighparvar, S.; Yousefi, B.; Kaviani, M.; Ghaderi-Pakdel, F.; Mihanfar, A.; Rahimi, Y.; Mobaraki, K.; Majidinia, M. Quercetin attenuated oxidative DNA damage through NRF2 signaling pathway in rats with DMH induced colon carcinogenesis. Life Sci. 2020, 253, 117584. [Google Scholar] [CrossRef]
- Tian, R.; Yang, W.; Xue, Q.; Gao, L.; Huo, J.; Ren, D.; Chen, X. Rutin ameliorates diabetic neuropathy by lowering plasma glucose and decreasing oxidative stress via Nrf2 signaling pathway in rats. Eur. J. Pharmacol. 2016, 771, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Suraweera, T.L.; Rupasinghe, H.P.V.; Dellaire, G.; Xu, Z. Regulation of Nrf2/ARE Pathway by Dietary Flavonoids: A Friend or Foe for Cancer Management? Antioxidants 2020, 9, 973. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Zhou, L. The Signaling of Cellular Senescence in Diabetic Nephropathy. Oxidative Med. Cell. Longev. 2019, 2019, 7495629-16. [Google Scholar] [CrossRef] [PubMed]
- Elbe, H.; Vardi, N.; Esrefoglu, M.; Ates, B.; Yologlu, S.; Taskapan, C. Amelioration of streptozotocin-induced diabetic nephropathy by melatonin, quercetin, and resveratrol in rats. Hum. Exp. Toxicol. 2014, 34, 100–113. [Google Scholar] [CrossRef]
- Ding, T.; Wang, S.; Zhang, X.; Zai, W.; Fan, J.; Chen, W.; Bian, Q.; Luan, J.; Shen, Y.; Zhang, Y.; et al. Kidney protection effects of dihydroquercetin on diabetic nephropathy through suppressing ROS and NLRP3 inflammasome. Phytomedicine 2018, 41, 45–53. [Google Scholar] [CrossRef]
- Tang, L.; Li, K.; Zhang, Y.; Li, H.; Li, A.; Xu, Y.; Wei, B. Quercetin liposomes ameliorate streptozotocin-induced diabetic nephropathy in diabetic rats. Sci. Rep. 2020, 10, 2440. [Google Scholar] [CrossRef] [Green Version]
- Lai, P.B.; Zhang, L.; Yang, L.Y. Quercetin ameliorates diabetic nephropathy by reducing the expressions of transforming growth factor-beta1 and connective tissue growth factor in streptozotocin-induced diabetic rats. Ren. Fail. 2012, 34, 83–87. [Google Scholar] [CrossRef]
- Lei, D.; Chengcheng, L.; Xuan, Q.; Yibing, C.; Lei, W.; Hao, Y.; Xizhi, L.; Yuan, L.; Xiaoxing, Y.; Qian, L. Quercetin inhibited mesangial cell proliferation of early diabetic nephropathy through the Hippo pathway. Pharmacol. Res. 2019, 146, 104320. [Google Scholar] [CrossRef]
- Luo, W.; Chen, X.; Ye, L.; Chen, X.; Jia, W.; Zhao, Y.; Samorodov, A.V.; Zhang, Y.; Hu, X.; Zhuang, F.; et al. Kaempferol attenuates streptozotocin-induced diabetic nephropathy by downregulating TRAF6 expression: The role of TRAF6 in diabetic nephropathy. J. Ethnopharmacol. 2020, 268, 113553. [Google Scholar] [CrossRef]
- Sharma, D.; Tekade, R.K.; Kalia, K. Kaempferol in ameliorating diabetes-induced fibrosis and renal damage: An in vitro and in vivo study in diabetic nephropathy mice model. Phytomedicine 2020, 76, 153235. [Google Scholar] [CrossRef]
- Alshehri, A.S. Kaempferol attenuates diabetic nephropathy in streptozotocin-induced diabetic rats by a hypoglycaemic effect and concomitant activation of the Nrf-2/Ho-1/antioxidants axis. Arch. Physiol. Biochem. 2021, 24, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.-H.; Shao, Z.-M.; Tang, D.-Q.; Lu, Q.; Chen, X.; Yin, X.-X.; Wu, J.; Chen, H. Preventive effects of rutin on the development of experimental diabetic nephropathy in rats. Life Sci. 2012, 91, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Kamalakkannan, N.; Prince, P.S.M. The influence of rutin on the extracellular matrix in streptozoto-cin-induced diabetic rat kidney. J. Pharm. Pharmacol. 2006, 58, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, D.; Albert, A.; Paul, E.; Ananthapadmanabhan, K.; Andiappan, R.; Sadasivam, S.G. Rutin ameliorates metabolic acidosis and fibrosis in alloxan induced diabetic nephropathy and cardiomyopathy in experimental rats. Mol. Cell. Biochem. 2020, 471, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, D.; Holkar, A.; Albert, A.; Paul, E.; Mariakuttikan, J.; Selvam, G.S. Combination of ramipril and rutin alleviate alloxan induced diabetic nephropathy targeting multiple stress pathways in vivo. Biomed. Pharmacother. 2018, 108, 1338–1346. [Google Scholar] [CrossRef]
- Zhang, M.; He, L.; Liu, J.; Zhou, L. Luteolin Attenuates Diabetic Nephropathy through Suppressing Inflammatory Response and Oxidative Stress by Inhibiting STAT3 Pathway. Exp. Clin. Endocrinol. Diabetes 2020, 129, 729–739. [Google Scholar] [CrossRef]
- Xiong, C.; Wu, Q.; Fang, M.; Li, H.; Chen, B.; Chi, T. Protective effects of luteolin on nephrotoxicity induced by long-term hyperglycaemia in rats. J. Int. Med Res. 2020, 48. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.G.; Lu, X.H.; Li, W.; Zhao, X.; Zhang, C. Protective Effects of Luteolin on Diabetic Nephropathy in STZ-Induced Diabetic Rats. Evid.-Based Complement. Altern. Med. 2011, 2011, 323171. [Google Scholar] [CrossRef] [Green Version]
- Ou, Y.; Zhang, W.J.; Chen, S.P.; Deng, H.H. Baicalin improves podocyte injury in rats with diabetic nephropathy by inhibiting PI3K/Akt/ m, TOR signaling pathway. Open Med. (Wars.) 2021, 16, 1286–1298. [Google Scholar] [CrossRef]
- Roy, S.; Ahmed, F.; Banerjee, S.; Saha, U. Naringenin ameliorates streptozotocin-induced diabetic rat renal impairment by downregulation of TGF-beta1 and IL-1 via modulation of oxidative stress correlates with decreased apoptotic events. Pharm. Biol. 2016, 54, 1616–1627. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.F.; Mathur, A.; Pandey, V.K.; Kakkar, P. Naringenin alleviates hyperglycemia-induced renal toxicity by regu-lating activating transcription factor 4-C/EBP homologous protein mediated apoptosis. J. Cell Commun. Signal 2021, 16, 271–291. [Google Scholar] [CrossRef] [PubMed]
- Dokumacioglu, E.; Iskender, H.; Musmul, A. Effect of hesperidin treatment on α-Klotho/FGF-23 pathway in rats with experimentally-induced diabetes. Biomed. Pharmacother. 2019, 109, 1206–1210. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Kong, L.; Tang, Z.Z.; Zhang, Y.M.; Liu, Y.; Wang, T.Y.; Liu, Y.W. Hesperetin ameliorates diabetic nephropathy in rats by activating Nrf2/ARE/glyoxalase 1 pathway. Biomed. Pharmacother. 2019, 111, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.T.; Wang, G.; Ye, L.F.; Pu, Y.; Li, R.T.; Liang, J.X.; Wang, L.J.; Lee, K.K.H.; Yang, X.S. Baicalin reversal of DNA hy-permethylation-associated Klotho suppression ameliorates renal injury in type 1 diabetic mouse model. Cell Cycle 2020, 19, 3329–3347. [Google Scholar] [CrossRef] [PubMed]
- Elmarakby, A.A.; Ibrahim, A.S.; Faulkner, J.; Mozaffari, M.S.; Liou, G.I.; Abdelsayed, R. Tyrosine kinase inhibitor, genistein, reduces renal inflammation and injury in streptozotocin-induced diabetic mice. Vasc. Pharmacol. 2011, 55, 149–156. [Google Scholar] [CrossRef]
- Ma, L.; Wu, F.; Shao, Q.; Chen, G.; Xu, L.; Lu, F. Baicalin Alleviates Oxidative Stress and Inflammation in Diabetic Nephropathy via Nrf2 and MAPK Signaling Pathway. Drug Des. Dev. Ther. 2021, 15, 3207–3221. [Google Scholar] [CrossRef]
- Kao, T.-W.; Huang, C.-C. Blood pressure management and renal protection: Revisiting hypertensive nephropathy. J. Chin. Med Assoc. 2021, 84, 911–916. [Google Scholar] [CrossRef]
- Seccia, T.M.; Caroccia, B.; Calo, L.A. Hypertensive nephropathy. Moving from classic to emerging pathogenetic mechanisms. J. Hypertens. 2017, 35, 205–212. [Google Scholar] [CrossRef]
- Satou, R.; Shao, W.; Navar, L.G. Role of stimulated intrarenal angiotensinogen in hypertension. Ther. Adv. Cardiovasc. Dis. 2015, 9, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Brown, I.A.; Diederich, L.; Good, M.; De Lalio, L.; Murphy, S.; Cortese-Krott, M.M.; Hall, J.L.; Le, T.H.; Isakson, B.E. Vascular Smooth Muscle Remodeling in Conductive and Resistance Arteries in Hypertension. Arter. Thromb. Vasc. Biol. 2018, 38, 1969–1985. [Google Scholar] [CrossRef]
- Hill, G.S. Hypertensive nephrosclerosis. Curr. Opin. Nephrol. Hypertens. 2008, 17, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.; Jimenez, R.; O’Valle, F.; Galisteo, M.; Pérez-Palencia, R.; Vargas, F.; Perez-Vizcaino, F.; Zarzuelo, A.; Tamargo, J. Protective effects of the flavonoid quercetin in chronic nitric oxide deficient rats. J. Hypertens. 2002, 20, 1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Galisteo, M.; García-Saura, M.F.; Jimenez, R.; Villar, I.C.; Zarzuelo, A.; Vargas, F.; Duarte, J. Effects of chronic quercetin treatment on antioxidant defence system and oxidative status of deoxycorticosterone acetate-salt-hypertensive rats. Mol. Cell. Biochem. 2004, 259, 91–99. [Google Scholar] [CrossRef]
- Galisteo, M.; García-Saura, M.F.; Jiménez, R.; Villar, I.C.; Wangensteen, R.; Zarzuelo, A.; Vargas, F.; Duarte, J. Effects of Quercetin Treatment on Vascular Function in Deoxycorticosterone Acetate-Salt Hypertensive Rats. Comparative Study with Verapamil. Planta Medica 2004, 70, 334–341. [Google Scholar]
- García-Saura, M.F.; Galisteo, M.; Villar, I.C.; Bermejo, A.; Zarzuelo, A.; Vargas, F.; Duarte, J. Effects of chronic quercetin treatment in experimental renovascular hypertension. Mol. Cell. Biochem. 2005, 270, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Guzmán, M.; Jiménez, R.; Sánchez, M.; Zarzuelo, M.J.; Galindo, P.; Quintela, A.M.; López-Sepúlveda, R.; Romero, M.; Tamargo, J.; Vargas, F.; et al. Epicatechin lowers blood pressure, restores endothelial function, and decreases oxidative stress and endothelin-1 and NADPH oxidase activity in DOCA-salt hypertension. Free Radic. Biol. Med. 2012, 52, 70–79. [Google Scholar] [CrossRef]
- Jiménez, R.; López-Sepúlveda, R.; Kadmiri, M.; Romero, M.; Vera, R.; Sánchez, M.; Vargas, F.; O’Valle, F.; Zarzuelo, A.; Dueñas, M. Polyphenols restore endothelial function in DOCA-salt hypertension: Role of endothelin-1 and NADPH oxidase. Free Radic. Biol. Med. 2007, 43, 462–473. [Google Scholar] [CrossRef]
- Prahalathan, P.; Kumar, S.; Raja, B. Effect of morin, a flavonoid against DOCA-salt hypertensive rats: A dose dependent study. Asian Pac. J. Trop. Biomed. 2012, 2, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Muthuraman, A. Therapeutic evaluation of rutin in two-kidney one-clip model of renovascular hypertension in rat. Life Sci. 2016, 150, 89–94. [Google Scholar] [CrossRef]
- Lan, C.-Z.; Ding, L.; Su, Y.-L.; Guo, K.; Wang, L.; Kan, H.-W.; Ou, Y.-R.; Gao, S. Grape seed proanthocyanidins prevent DOCA-salt hypertension-induced renal injury and its mechanisms in rats. Food Funct. 2015, 6, 2179–2186. [Google Scholar] [CrossRef]
- Wang, Q.Z.; Gao, H.Q.; Liang, Y.; Zhang, J.; Wang, J.; Qiu, J. Cofilin1 is involved in hypertension-induced renal damage via the regulation of NF-kappa, B in renal tubular epithelial cells. J. Transl. Med. 2015, 13, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanisamy, N.; Venkataraman, A.C. Beneficial effect of genistein on lowering blood pressure and kidney toxicity in fructose-fed hypertensive rats. Br. J. Nutr. 2012, 109, 1806–1812. [Google Scholar] [CrossRef] [Green Version]
- Floege, J.; Amann, K. Primary glomerulonephritides. Lancet 2016, 387, 2036–2048. [Google Scholar] [CrossRef]
- Dou, Y.; Shang, Y.; Shen, Y.; Qu, J.; Liu, C.; Cao, J. Baicalin alleviates adriamycin-induced focal segmental glomerulosclerosis and proteinuria by inhibiting the Notch1-Snail axis mediated podocyte EMT. Life Sci. 2020, 257, 118010. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-N.; Li, A.-P.; Qi, Y.-S.; Qin, X.-M.; Li, Z.-Y. Metabolomics coupled with system pharmacology reveal the protective effect of total flavonoids of Astragali Radix against adriamycin-induced rat nephropathy model. J. Pharm. Biomed. Anal. 2018, 158, 128–136. [Google Scholar] [CrossRef]
- El-Shitany, N.A.; El-Haggar, S.; El-Desoky, K. Silymarin prevents adriamycin-induced cardiotoxicity and nephrotoxicity in rats. Food Chem. Toxicol. 2008, 46, 2422–2428. [Google Scholar] [CrossRef]
- Chen, Z.; An, X.; Liu, X.; Qi, J.; Ding, D.; Zhao, M.; Duan, S.; Huang, Z.; Zhang, C.; Wu, L.; et al. Hyperoside alleviates adriamycin-induced podocyte injury via inhibiting mitochondrial fission. Oncotarget 2017, 8, 88792–88803. [Google Scholar] [CrossRef] [Green Version]
- Peng, A.; Ye, T.; Rakheja, D.; Tu, Y.; Wang, T.; Du, Y.; Zhou, J.K.; Vaziri, N.D.; Hu, Z.; Mohan, C.; et al. The green tea polyphenol (−)-epigallocatechin-3-gallate ameliorates experimental immune-mediated glomerulonephritis. Kidney Int. 2011, 80, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; He, L. Epigallocatechin-3-Gallate Attenuates Adriamycin-Induced Focal Segmental Glomerulosclerosis via Suppression of Oxidant Stress and Apoptosis by Targeting Hypoxia-Inducible Factor-1alpha/ Angiopoietin-Like 4 Pathway. Pharmacology 2019, 103, 303–314. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.-Z.; Li, Y.-S.; Hao, L.-R. Icariin ameliorates Ig, A nephropathy by inhibition of nuclear factor kappa b/Nlrp3 pathway. FEBS Open Bio 2016, 7, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Anders, H.J.; Saxena, R.; Zhao, M.H.; Parodis, I.; Salmon, J.E.; Mohan, C. Lupus nephritis. Nat. Rev. Dis. Primers 2020, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Bagavant, H.; Fu, S.M. Pathogenesis of kidney disease in systemic lupus erythematosus. Curr. Opin. Rheumatol. 2009, 21, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, R.; Guo, C.; Wang, S.; Huang, Y.; Jin, O.; Hu, H.; Chen, J.; Xu, B.; Zhou, M.; Zhao, J.; et al. Podocyte Activation of NLRP3 Inflammasomes Contributes to the Development of Proteinuria in Lupus Nephritis. Arthritis Rheumatol. 2017, 69, 1636–1646. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yu, C.; Ji, K.; Wang, X.; Li, X.; Xie, H.; Wang, Y.; Huang, Y.; Qi, D.; Fan, H. Quercetin reduces TNF-alpha-induced mesangial cell proliferation and inhibits PTX3 production: Involvement of NF-kappa, B signaling pathway. Phytother. Res. 2019, 33, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Shi, G.P.; Wang, J.L.; Zhang, D.Y.; Pan, Y.C.; Dou, H.; Hou, Y.Y. Baicalein ameliorates pristane-induced lupus nephritis via activating Nrf2/HO-1 in myeloid-derived suppressor cells. Arthritis Res. Ther. 2019, 21, 105. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yang, X.; Li, M. Baicalin ameliorates lupus autoimmunity by inhibiting differentiation of Tfh cells and inducing expansion of Tfr cells. Cell Death Dis. 2019, 10, 140. [Google Scholar] [CrossRef]
- Abrego-Peredo, A.; Romero-Ramírez, H.; Espinosa, E.; López-Herrera, G.; García-García, F.; Flores-Muñoz, M.; Sandoval-Montes, C.; Rodríguez-Alba, J.C. Naringenin mitigates autoimmune features in lupus-prone mice by modulation of T-cell subsets and cytokines profile. PLoS ONE 2020, 15, e0233138. [Google Scholar] [CrossRef]
- Su, B.; Ye, H.; You, X.; Ni, H.; Chen, X.; Li, L. Icariin alleviates murine lupus nephritis via inhibiting NF-kappa, B activation pathway and NLRP3 inflammasome. Life Sci. 2018, 208, 26–32. [Google Scholar] [CrossRef]
- dos Santos, M.; Poletti, P.T.; Favero, G.; Stacchiotti, A.; Bonomini, F.; Montanari, C.C.; Bona, S.R.; Marroni, N.P.; Rezzani, R.; Veronese, F.V. Protective effects of quercetin treatment in a pristane-induced mouse model of lupus nephritis. Autoimmunology 2018, 51, 69–80. [Google Scholar] [CrossRef]
- Li, W.; Li, H.; Zhang, M.; Wang, M.Q.; Zhong, Y.X.; Wu, H.Z.; Yang, Y.F.; Morel, L.; Wei, Q. Quercitrin ameliorates the de-velopment of systemic lupus erythematosus-like disease in a chronic graft-versus-host murine model. Am. J. Physiol. Renal. 2016, 311, F217–F226. [Google Scholar] [CrossRef] [Green Version]
- He, J.H.; Sun, M.C.; Tian, S.J. Procyanidin B2 prevents lupus nephritis development in mice by inhibiting NLRP3 in-flammasome activation. Innate. Immun. 2018, 24, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-K.; Ecklund, D.; Liu, M.; Datta, S.K. Apigenin, a non-mutagenic dietary flavonoid, suppresses lupus by inhibiting autoantigen presentation for expansion of autoreactive Th1 and Th17 cells. Arthritis Res. Ther. 2009, 11, R59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, P.-Y.; Ka, S.-M.; Chang, J.-M.; Chen, H.-C.; Shui, H.-A.; Li, C.-Y.; Hua, K.-F.; Chang, W.-L.; Huang, J.-J.; Yang, S.-S.; et al. Epigallocatechin-3-gallate prevents lupus nephritis development in mice via enhancing the Nrf2 antioxidant pathway and inhibiting NLRP3 inflammasome activation. Free Radic. Biol. Med. 2011, 51, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, Y. Fisetin inhibits pristine-induced systemic lupus erythematosus in a murine model through CXCLs regulation. Int. J. Mol. Med. 2018, 42, 3220–3230. [Google Scholar] [CrossRef]
- Guo, L.; Liu, W.; Lu, T.; Guo, W.; Gao, J.; Luo, Q.; Wu, X.; Sun, Y.; Wu, X.; Shen, Y.; et al. Decrease of Functional Activated T and B Cells and Treatment of Glomerulonephitis in Lupus-Prone Mice Using a Natural Flavonoid Astilbin. PLoS ONE 2015, 10, e0124002. [Google Scholar] [CrossRef]



| GFR Category | |
|---|---|
| G1 | ≥90 mL/min/1.73 m2 |
| G2 | 60–89 mL/min/1.73 m2 |
| G3a | 45–59 mL/min/1.73 m2 |
| G3b | 30–44 mL/min/1.73 m2 |
| G4 | 15–29 mL/min/1.73 m2 |
| G5 | <15 mL/min/1.73 m2 |
| Albuminuria Category | |
| A1 | ACR < 30 mg/g |
| A2 | ACR 30–300 mg/g |
| A3 | ACR > 300 mg/g |
| Animal Models | Flavonoids | Functions | References |
|---|---|---|---|
| DOCA-salt rats | Quercetin | Restoring total GSH levels and reducing TBARS level; restoring MDA content and SOD expression and improving potassium depletion | [173,174] |
| Epicatechin | Reducing NADPH oxidase activity and ET-1 levels; increasing Nrf2 | [176] | |
| Red wine polyphenols | Reducing NADPH oxidase activity and ET-1 levels | [177] | |
| Grape seed proanthocyanidins | Suppressing the JNK/p38 kinase pathway | [180] | |
| Morin | Reducing plasma urea, uric acid, and creatinine levels | [178] | |
| 2K1C rats | Quercetin | Restoring total GSH content and reducing the vasoconstrictor TXA2 | [175] |
| Rutin | Decreasing tissue TBARS and increasing GSH levels | [179] | |
| L-NAME rats | Quercetin | Reducing renal hypertrophy, proteinuria, renal parenchyma, and vascular lesions | [172] |
| SHRs | Grape seed proanthocyanidins | Upregulating cofilin1 and inhibiting the NF-κB pathway | [181] |
| Fructose-fed hypertensive rats | Genistein | Inhibiting ACE and PKC-βII and activating eNOS and NO synthesis | [182] |
| Animal Models | Flavonoids | Functions | References |
|---|---|---|---|
| Pristane-induced lupus mice | Baicalin | Inducing Nrf2/HO-1 signal and NLRP3 expression | [195] |
| Fisetin | Reducing Th17 cells; inhibiting the CXCL signaling pathway | [204] | |
| Quercetin | Increasing CAT and SOD1 expressions; lowering IL-6, TNF-α, TGF-β1, Bax, and TBARS | [199] | |
| Lupus-prone MRL/lpr mice | Baicalin | Inhibiting mTOR activation; reducing mTOR-mediated Tfh cell expansion; increasing Tfr cells | [196] |
| Naringenin | Decreasing anti-nuclear and anti-dsDNA autoantibodies; increasing the percentage of Treg cells | [197] | |
| Icariin | Suppressing the NLRP3 inflammasome and the NF-κB signaling pathway | [198] | |
| Procyanidin B2 | Inhibiting IL-1β, IL-18, and NLRP3 inflammasome | [201] | |
| Astilbin | Decreasing functional activated T and B cells | [205] | |
| Lupus-prone SNF1 mice | Epigallocatechin-3- gallate | Enhancing the Nrf2 antioxidant pathway and inhibiting the NLRP3 inflammasome | [203] |
| Apigenin | Inhibiting autoantigen-presenting and stimulatory functions of APCs; causing apoptosis of hyperactive lupus APCs and T and B cells | [202] | |
| Chronic GVHD mouse model | Quercitrin | Inhibiting CD4 + T cell activation | [200] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.-L.; Lin, J.-H.; Hammes, H.-P.; Zhang, C. Flavonoids in Treatment of Chronic Kidney Disease. Molecules 2022, 27, 2365. https://doi.org/10.3390/molecules27072365
Cao Y-L, Lin J-H, Hammes H-P, Zhang C. Flavonoids in Treatment of Chronic Kidney Disease. Molecules. 2022; 27(7):2365. https://doi.org/10.3390/molecules27072365
Chicago/Turabian StyleCao, Yi-Ling, Ji-Hong Lin, Hans-Peter Hammes, and Chun Zhang. 2022. "Flavonoids in Treatment of Chronic Kidney Disease" Molecules 27, no. 7: 2365. https://doi.org/10.3390/molecules27072365
APA StyleCao, Y. -L., Lin, J. -H., Hammes, H. -P., & Zhang, C. (2022). Flavonoids in Treatment of Chronic Kidney Disease. Molecules, 27(7), 2365. https://doi.org/10.3390/molecules27072365