
Effect of Tomato Peel Extract Grown under Drought Stress Condition in a Sarcopenia Model

 ,
, 
 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
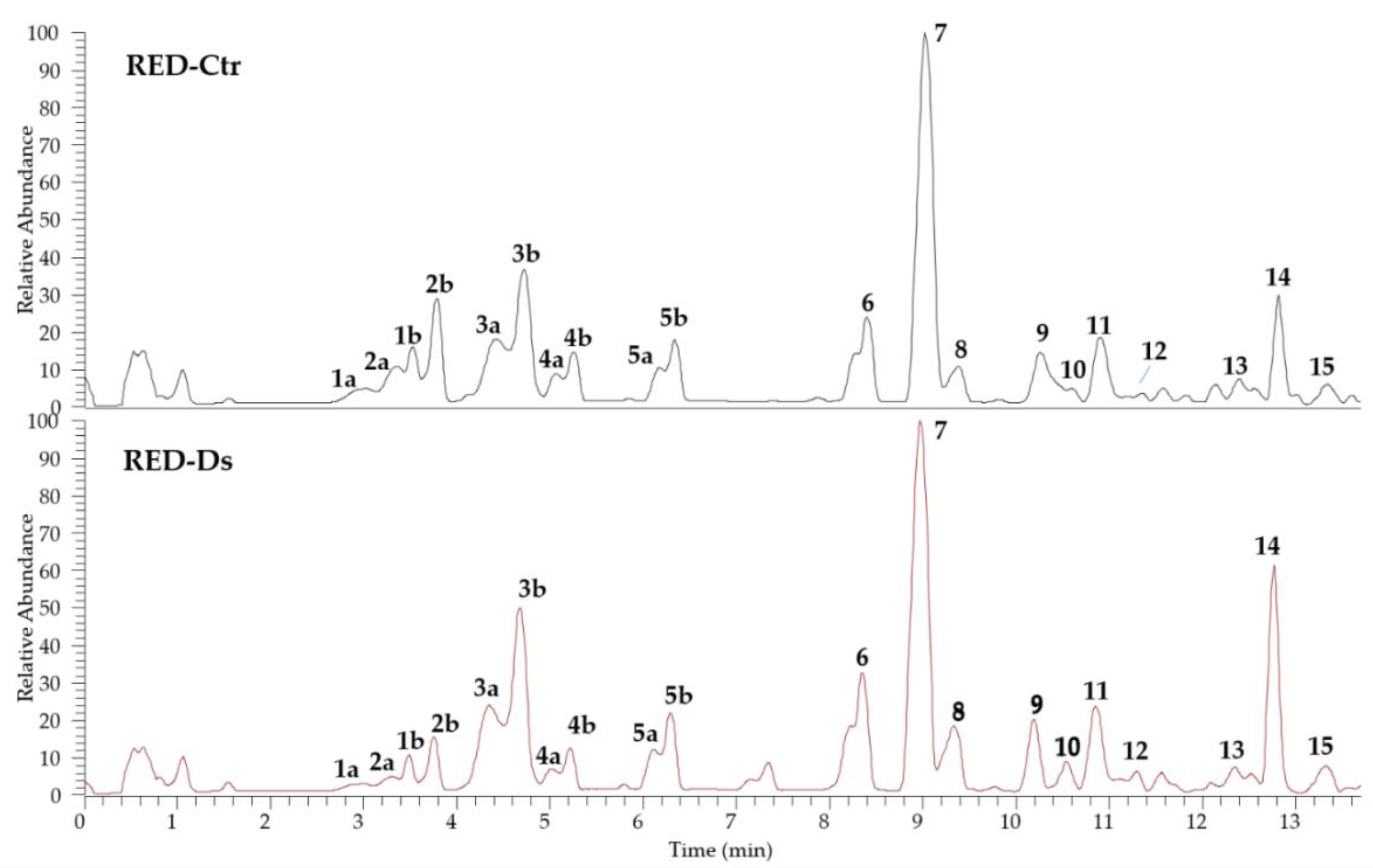
2.1. Characterization and Phenolic Profile of Tomato Peel Extracts
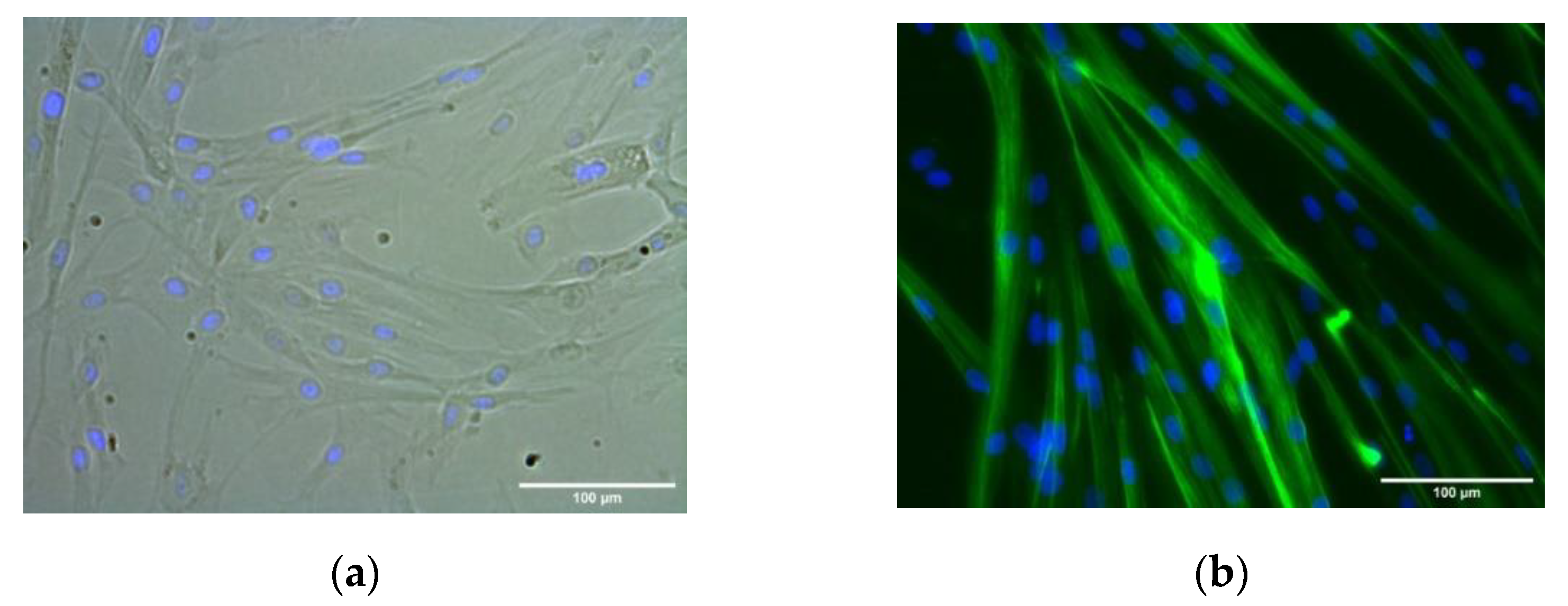
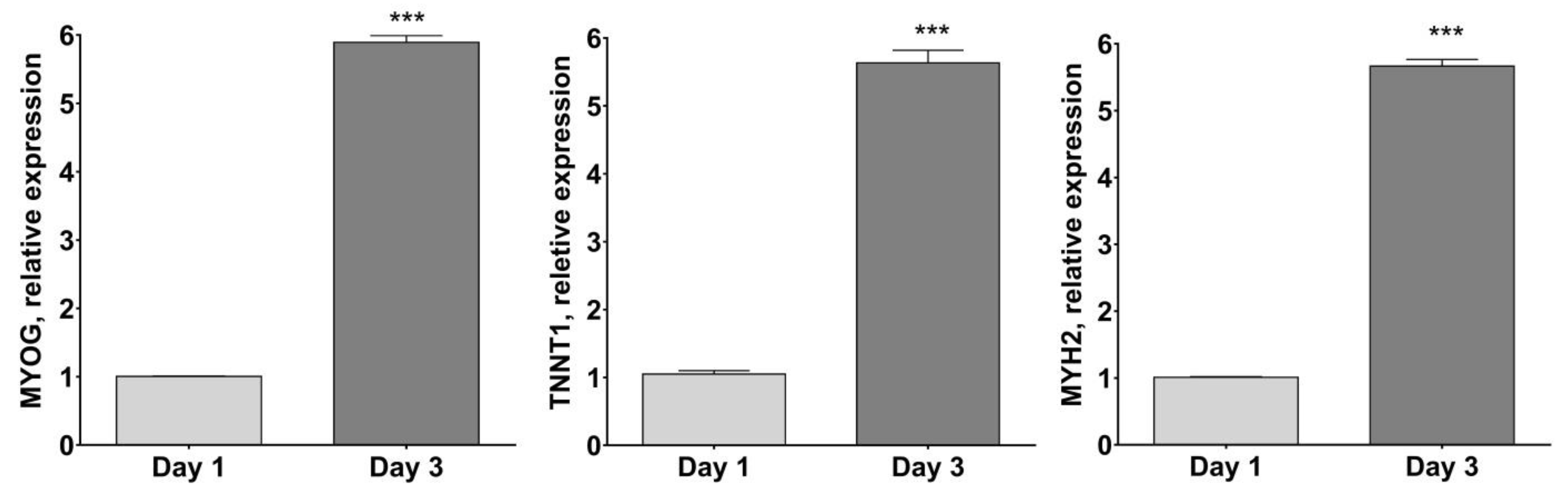
2.2. HSMM Differentiation into Multinucleated Myotubes
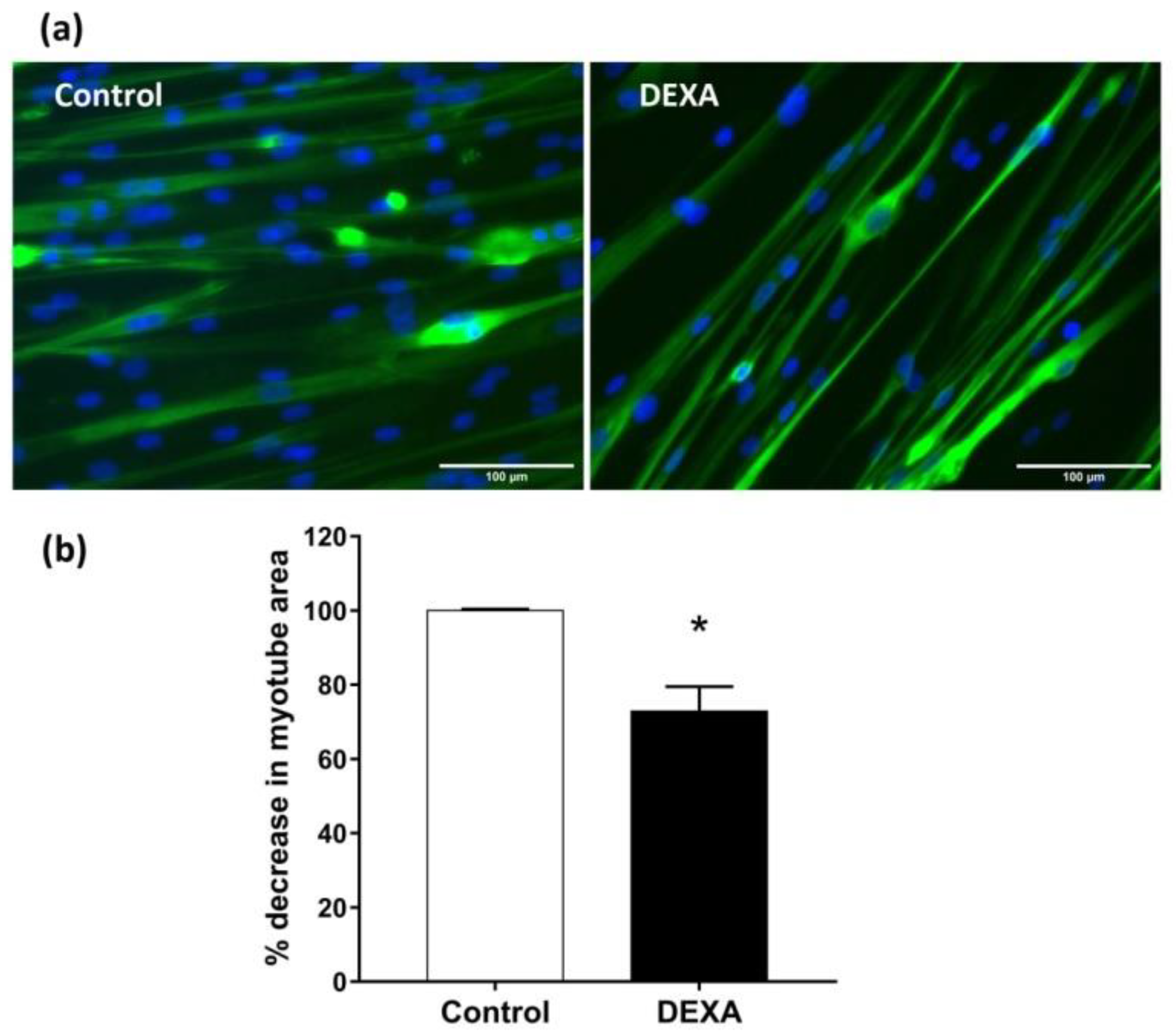
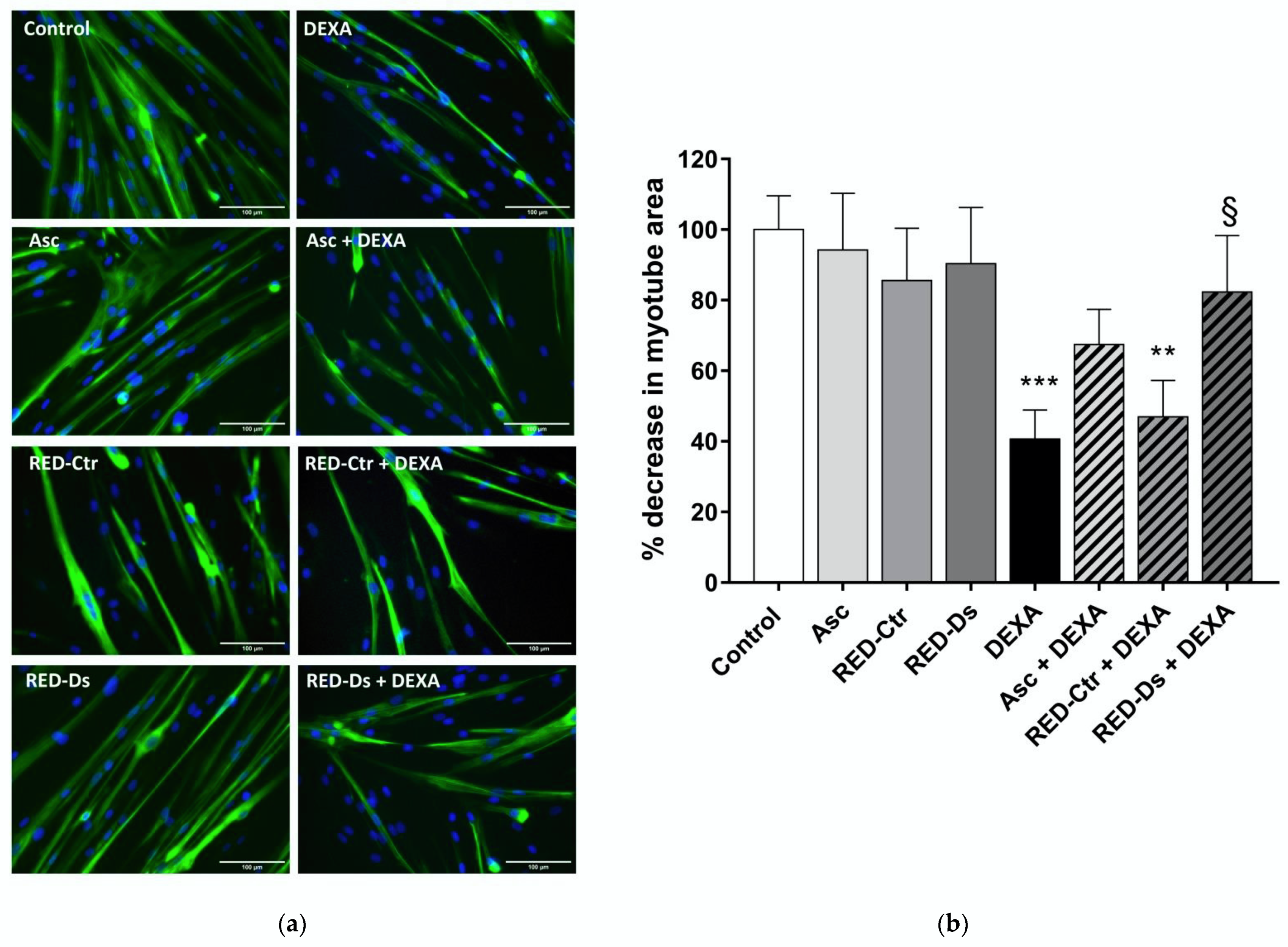
2.3. In Vitro Sarcopenia Induction
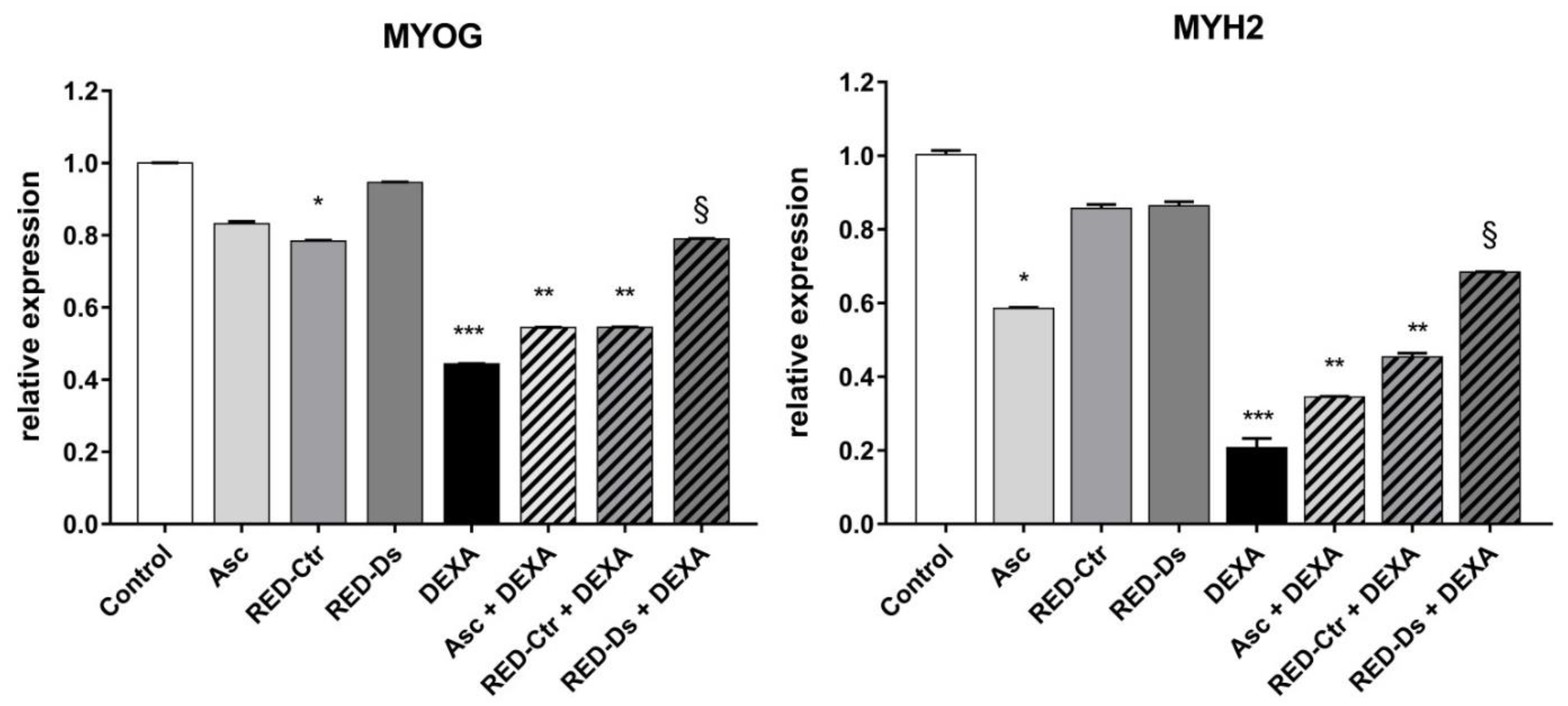
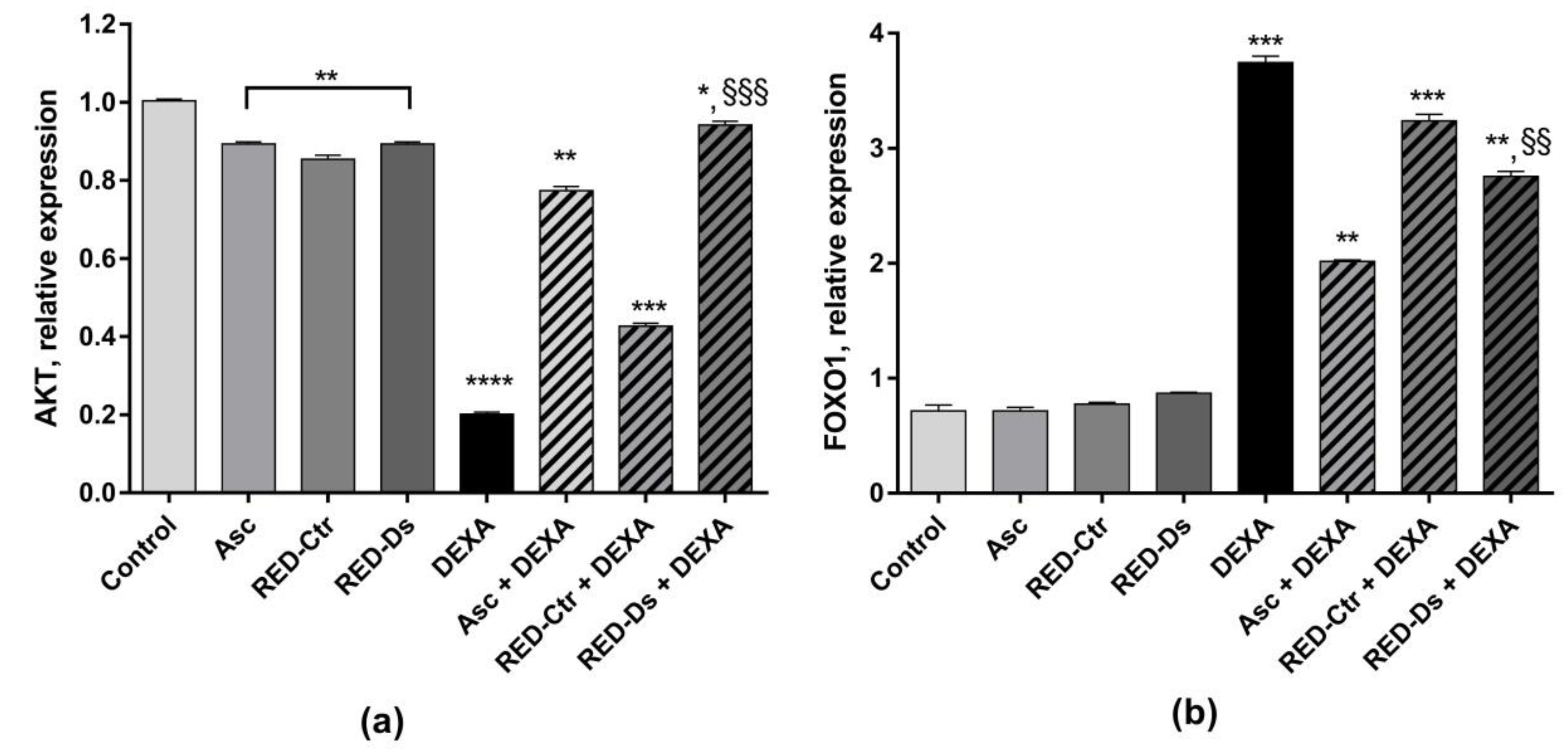
2.4. Tomato Peel Extracts Effect on Induced-Sarcopenia
3. Discussion
4. Materials and Methods
4.1. Cells, Media and Supplement
4.2. Sample Preparation
4.2.1. Fruit Harvesting and Growth Conditions
4.2.2. Tomato Peel Extracts Preparation
4.3. UHPLC-HR-ESI-MS Analyses of Tomato Peel Extracts
4.4. Cell Culture and Differentiation
4.5. Cell Treatment in Sarcopenia-Induced Model
4.6. Immunostaining and Fluorescence Analysis
4.7. RNA Extraction, Reverse Transcription and Real-Time PCR Experiments
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Marques, A.; Queiros, C. Frailty, Sarcopenia and Falls. In Fragility Fracture Nursing: Holistic Care and Management of the Orthogeriatric Patient; Hertz, K., Santy-Tomlinson, J., Eds.; Springer: Cham, Switzerland, 2018; pp. 15–26. [Google Scholar]
- Afroze, D.; Kumar, A. ER stress in skeletal muscle remodeling and myopathies. FEBS J. 2019, 286, 379–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, I.H. Sarcopenia: Origins and clinical relevance. J. Nutr. 1997, 127, 990S–991S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, J.; Morley, J.E.; Schols, A.; Ferrucci, L.; Cruz-Jentoft, A.J.; Dent, E.; Baracos, V.E.; Crawford, J.A.; Doehner, W.; Heymsfield, S.B.; et al. Sarcopenia: A time for action. An SCWD position paper. J. Cachexia Sarcopenia Muscle 2019, 10, 956–961. [Google Scholar] [CrossRef] [PubMed]
- Mastrocola, R.; Collino, M.; Nigro, D.; Chiazza, F.; D’Antona, G.; Aragno, M.; Minetto, M.A. Accumulation of advanced glycation end-products and activation of the SCAP/SREBP lipogenetic pathway occur in diet-induced obese mouse skeletal muscle. PLoS ONE 2015, 10, e0119587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auclair, D.; Garrel, D.R.; Chaouki Zerouala, A.; Ferland, L.H. Activation of the ubiquitin pathway in rat skeletal muscle by catabolic doses of glucocorticoids. Am. J. Physiol. 1997, 272, C1007–C1016. [Google Scholar] [CrossRef]
- Jackman, R.W.; Kandarian, S.C. The molecular basis of skeletal muscle atrophy. Am. J. Physiol. Cell Physiol. 2004, 287, C834–C843. [Google Scholar] [CrossRef] [Green Version]
- Clarke, B.A.; Drujan, D.; Willis, M.S.; Murphy, L.O.; Corpina, R.A.; Burova, E.; Rakhilin, S.V.; Stitt, T.N.; Patterson, C.; Latres, E.; et al. The E3 ligase MuRF1 degrades myosin heavy chain protein in dexamethasone-treated skeletal muscle. Cell Metab. 2007, 6, 376–385. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Qin, W.; Pan, J.; Wu, Y.; Bauman, W.A.; Cardozo, C. Dependence of dexamethasone-induced Akt/FOXO1 signaling, upregulation of MAFbx, and protein catabolism upon the glucocorticoid receptor. Biochem. Biophys. Res. Commun. 2009, 378, 668–672. [Google Scholar] [CrossRef]
- Waddell, D.S.; Baehr, L.M.; Van Den Brandt, J.; Johnsen, S.A.; Reichardt, H.M.; Furlow, J.D.; Bodine, S.C. The glucocorticoid receptor and FOXO1 synergistically activate the skeletal muscle atrophy-associated MuRF1 gene. Am. J. Physiol.-Endocrinol. Metab. 2008, 295, E785–E797. [Google Scholar] [CrossRef]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.; Mallidis, C.; Bhasin, S.; Mahabadi, V.; Artaza, J.; Gonzalez-Cadavid, N.; Arias, J.; Salehian, B. Glucocorticoid-induced skeletal muscle atrophy is associated with upregulation of myostatin gene expression. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E363–E371. [Google Scholar] [CrossRef]
- Chen, C.; Yang, J.S.; Lu, C.C.; Chiu, Y.J.; Chen, H.C.; Chung, M.I.; Wu, Y.T.; Chen, F.A. Effect of quercetin on dexamethasone-induced C2C12 skeletal muscle cell injury. Molecules 2020, 25, 3267. [Google Scholar] [CrossRef] [PubMed]
- Fougere, B.; van Kan, G.A.; Vellas, B.; Cesari, M. Redox systems, antioxidants and sarcopenia. Curr. Protein Pept. Sci. 2018, 19, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, Y.; Egawa, K.; Kanzaki, N.; Izumo, T.; Rogi, T.; Shibata, H. Quercetin glycosides prevent dexamethasone-induced muscle atrophy in mice. Biochem. Biophys. Rep. 2019, 18, 100618. [Google Scholar] [CrossRef] [PubMed]
- Salucci, S.; Falcieri, E. Polyphenols and their potential role in preventing skeletal muscle atrophy. Nutr. Res. 2020, 74, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.H.; Jang, E.J.; Kim, Y.W.; Lee, J.-H. Sulforaphane prevents dexamethasone-induced muscle atrophy via regulation of the Akt/Foxo1 axis in C2C12 myotubes. Biomed. Pharmacother. 2017, 95, 1486–1492. [Google Scholar] [CrossRef]
- Pellicanò, T.M.; Sicari, V.; Loizzo, M.R.; Leporini, M.; Falco, T.; Poiana, M. Optimizing the supercritical fluid extraction process of bioactive compounds from processed tomato skin by-products. Food Sci. Technol. 2020, 40, 692–697. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M. Anticarcinogenic, cardioprotective, and other health benefits of tomato compounds lycopene, alpha-tomatine, and tomatidine in pure form and in fresh and processed tomatoes. J. Agric. Food Chem. 2013, 61, 9534–9550. [Google Scholar] [CrossRef]
- Cesare, M.M.; Felice, F.; Conti, V.; Cerri, L.; Zambito, Y.; Romi, M.; Cai, G.; Cantini, C.; Di Stefano, R. Impact of peels extracts from an Italian ancient tomato variety grown under drought stress conditions on vascular related dysfunction. Molecules 2021, 26, 4289. [Google Scholar] [CrossRef]
- Barros, L.; Duenas, M.; Pinela, J.; Carvalho, A.M.; Buelga, C.S.; Ferreira, I.C. Characterization and quantification of phenolic compounds in four tomato (Lycopersicon esculentum L.) farmers’ varieties in northeastern Portugal homegardens. Plant Foods Hum. Nutr. 2012, 67, 229–234. [Google Scholar] [CrossRef]
- Slimestad, R.; Verheul, M. Review of flavonoids and other phenolics from fruits of different tomato (Lycopersicon esculentum Mill.) cultivars. J. Sci. Food Agric. 2009, 89, 1255–1270. [Google Scholar] [CrossRef]
- Moco, S.; Bino, R.J.; Vorst, O.; Verhoeven, H.A.; de Groot, J.; van Beek, T.A.; Vervoort, J.; de Vos, C.H. A liquid chromatography-mass spectrometry-based metabolome database for tomato. Plant Physiol. 2006, 141, 1205–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, J.; Moreira, K.; Bain, G. Characterization of primary human skeletal muscle cells from multiple commercial sources. Vitr. Cell Dev. Biol. Anim. 2013, 49, 695–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burattini, S.; Ferri, P.; Battistelli, M.; Curci, R.; Luchetti, F.; Falcieri, E. C2C12 murine myoblasts as a model of skeletal muscle development: Morpho-functional characterization. Eur. J. Histochem. 2004, 48, 223–233. [Google Scholar]
- Lecomte, V.; Meugnier, E.; Euthine, V.; Durand, C.; Freyssenet, D.; Nemoz, G.; Rome, S.; Vidal, H.; Lefai, E. A new role for sterol regulatory element binding protein 1 transcription factors in the regulation of muscle mass and muscle cell differentiation. Mol. Cell Biol. 2010, 30, 1182–1198. [Google Scholar] [CrossRef] [Green Version]
- Shima, A.; Pham, J.; Blanco, E.; Barton, E.R.; Sweeney, H.L.; Matsuda, R. IGF-I and vitamin C promote myogenic differentiation of mouse and human skeletal muscle cells at low temperatures. Exp. Cell Res. 2011, 317, 356–366. [Google Scholar] [CrossRef]
- Viuda-Martos, M.; Sanchez-Zapata, E.; Sayas-Barbera, E.; Sendra, E.; Perez-Alvarez, J.A.; Fernandez-Lopez, J. Tomato and tomato byproducts. Human health benefits of lycopene and its application to meat products: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1032–1049. [Google Scholar] [CrossRef]
- Qin, J.; Du, R.; Yang, Y.-Q.; Zhang, H.-Q.; Li, Q.; Liu, L.; Guan, H.; Hou, J.; An, X.-R. Dexamethasone-induced skeletal muscle atrophy was associated with upregulation of myostatin promoter activity. Res. Vet. Sci. 2013, 94, 84–89. [Google Scholar] [CrossRef]
- Di Cesare Mannelli, L.; Micheli, L.; Lucarini, E.; Parisio, C.; Toti, A.; Tenci, B.; Zanardelli, M.; Branca, J.J.V.; Pacini, A.; Ghelardini, C. Effects of the combination of β-hydroxy-β-methyl butyrate and R(+) lipoic acid in a cellular model of sarcopenia. Molecules 2020, 25, 2117. [Google Scholar] [CrossRef]
- Nikawa, T.; Ulla, A.; Sakakibara, I. Polyphenols and their effects on muscle atrophy and muscle health. Molecules 2021, 26, 4887. [Google Scholar] [CrossRef]
- Upadhyay, R.; Mohan Rao, L.J. An outlook on chlorogenic acids-occurrence, chemistry, technology, and biological activities. Crit. Rev. Food Sci. Nutr. 2013, 53, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Zhou, C.Y.; Qiu, C.H.; Lu, X.M.; Wang, Y.T. Chlorogenic acid induced apoptosis and inhibition of proliferation in human acute promyelocytic leukemia HL60 cells. Mol. Med. Rep. 2013, 8, 1106–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, M.S.; Jiao, D.; Chan, B.C.; Hon, K.L.; Leung, P.C.; Lau, C.B.; Wong, E.C.; Cheng, L.; Chan, C.K.; Lam, C.W.; et al. Anti-inflammatory activities of pentaherbs formula, berberine, gallic acid and chlorogenic acid in atopic dermatitis-like skin inflammation. Molecules 2016, 21, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, H.Y.; Li, Z.Y.; Zheng, Y.; Chen, Y.; Zhou, Z.H.; Jin, J. The attenuation of chlorogenic acid on oxidative stress for renal injury in streptozotocin-induced diabetic nephropathy rats. Arch. Pharm. Res. 2016, 39, 989–997. [Google Scholar] [CrossRef]
- Ommati, M.M.; Farshad, O.; Mousavi, K.; Khalili, M.; Jamshidzadeh, A.; Heidari, R. Chlorogenic acid supplementation improves skeletal muscle mitochondrial function in a rat model of resistance training. Biologia 2020, 75, 1221–1230. [Google Scholar] [CrossRef]
- Chang, W.T.; Huang, S.C.; Cheng, H.L.; Chen, S.C.; Hsu, C.L. Rutin and gallic acid regulates mitochondrial functions via the SIRT1 pathway in C2C12 myotubes. Antioxidants 2021, 10, 286. [Google Scholar] [CrossRef]
- Ilavenil, S.; Kim da, H.; Srigopalram, S.; Arasu, M.V.; Lee, K.D.; Lee, J.C.; Lee, J.S.; Renganathan, S.; Choi, K.C. Potential application of p-coumaric acid on differentiation of C2C12 skeletal muscle and 3T3-L1 preadipocytes-an in vitro and in silico approach. Molecules 2016, 21, 997. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, M.; Bulzomi, P.; Galluzzo, P.; Lecis, M.; Leone, S.; Pallottini, V.; Marino, M. Naringenin modulates skeletal muscle differentiation via estrogen receptor α and β signal pathway regulation. Genes Nutr. 2014, 9, 425. [Google Scholar] [CrossRef]
- Butow, R.A.; Avadhani, N.G. Mitochondrial signaling: The retrograde response. Mol. Cell 2004, 14, 1–15. [Google Scholar] [CrossRef]
- Furuyama, T.; Kitayama, K.; Yamashita, H.; Mori, N. Forkhead transcription factor FOXO1 (FKHR)-dependent induction of PDK4 gene expression in skeletal muscle during energy deprivation. Biochem. J. 2003, 375, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Lecker, S.H.; Jagoe, R.T.; Gilbert, A.; Gomes, M.; Baracos, V.; Bailey, J.; Price, S.R.; Mitch, W.E.; Goldberg, A.L. Multiple types of skeletal muscle atrophy involve a common program of changes in gene expression. FASEB J. 2004, 18, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Conti, V.; Romi, M.; Guarnieri, M.; Cantini, C.; Cai, G. Italian tomato cultivars under drought stress show different content of bioactives in pulp and peel of fruits. Foods 2022, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Santos Sánchez, N.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva, C. Shikimic acid pathway in biosynthesis of phenolic compounds. In Plant Physiological Aspects of Phenolic Compounds; Intechopen: London, UK, 2019; Volume 1, pp. 1–15. [Google Scholar]
- Macaluso, M.; Bianchi, A.; Sanmartin, C.; Taglieri, I.; Venturi, F.; Testai, L.; Flori, L.; Calderone, V.; De Leo, M.; Braca, A.; et al. By-products from winemaking and olive mill value chains for the enrichment of refined olive oil: Technological challenges and nutraceutical features. Foods 2020, 9, 1390. [Google Scholar] [CrossRef] [PubMed]
- Semsarian, C.; Sutrave, P.; Richmond, D.R.; Graham, R.M. Insulin-like growth factor (IGF-I) induces myotube hypertrophy associated with an increase in anaerobic glycolysis in a clonal skeletal-muscle cell model. Biochem. J. 1999, 339, 443–451. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak a | Compound b | tR (min) | HR-[M−H]− (m/z) | HR-MS/MS Product Ions (m/z) c | Molecular Formula | Error (ppm) | RED Extract |
|---|---|---|---|---|---|---|---|
| Phenolic acids | |||||||
| 1a | Caffeic acid glucoside isomer I | 3.0 | 341.0875 | 179.03, 145.03, 161.02, 135.04 | C15H18O9 | −0.73 | Ctr, Ds |
| 2a | p-Coumaric acid glucoside isomer I | 3.3 | 325.0927 | 163.04, 119.05, 93.03 | C15H18O8 | −0.31 | Ctr, Ds |
| 1b | Caffeic acid glucoside isomer II | 3.5 | 341.0875 | 179.03, 145.03, 161.02, 135.04 | C15H18O9 | −0.73 | Ctr, Ds |
| 2b | p-Coumaric acid glucoside isomer II | 3.8 | 325.0927 | 163.04, 145.03, 119.05, 93.03 | C15H18O8 | −0.31 | Ctr, Ds |
| 3a | Chlorogenic acid isomer I d (3-O-caffeoylquinic acid) | 4.4 | 353.0878 | 191.06, 179.03, 173.04, 135.04 | C16H18O9 | 0.00 | Ctr, Ds |
| 3b | Chlorogenic acid isomer II d | 4.7 | 353.0878 | 191.06, 179.03, 173.04, 135.04 | C16H18O9 | 0.00 | Ctr, Ds |
| 4a | p-Coumaric acid glucoside isomer III | 5.1 | 325.0927 | 163.04, 145.03, 119.05, 93.03 | C15H18O9 | −0.31 | Ctr, Ds |
| 4b | p-Coumaric acid glucoside isomer IV | 5.2 | 325.0927 | 163.04, 145.03, 119.05, 93.03 | C15H18O9 | −0.31 | Ctr, Ds |
| 5a | Caffeoylquinic acid isomer I | 6.2 | 353.0878 | 191.06, 179.03, 135.04 | C16H18O9 | 0.00 | Ctr, Ds |
| 5b | Caffeoylquinic acid isomer II | 6.3 | 353.0878 | 191.06, 179.03, 135.04 | C16H18O9 | 0.00 | Ctr, Ds |
| 11 | Dicaffeoylquinic acid isomer I | 10.8 | 515.1191 | 353.09, 191.06, 179.03, 173.04, 135.04 | C25H24O12 | −0.78 | Ctr, Ds |
| 13 | Dicaffeoylquinic acid isomer II | 12.4 | 515.1191 | 353.09, 191.06, 179.03, 173.04, 135.04 | C25H24O12 | −0.78 | Ctr, Ds |
| Flavonoids | |||||||
| 6 | Quercetin 3-O-(2″-O-apiofuranosyl-6″-O-rhamno pyranosyl- glucopyranoside) | 7.3 | 741.1885 | 300.03, 301.02, 271.03 | C32H40O21 | +0.13 | Ctr, Ds |
| 7 | Rutin d | 9.0 | 609.1461 | 300.03, 301.02, 271.03 | C27H30O16 | 0.00 | Ctr, Ds |
| 8 | Kaempferol rutinoside-pentoside | 9.3 | 725.1932 | 284.03, 285.04, 255.03 | C32H38O19 | −0.28 | Ctr, Ds |
| 9 | Kaempferol 3-O-rutinoside | 10.2 | 593.1509 | 284.03, 285.04, 255.03 | C27H30O15 | −0.34 | Ctr, Ds |
| 10 | Naringenin 7-O-glucoside | 10.5 | 433.1138 | 271.06, 151.00, 119.05 | C21H22O10 | −0.46 | Ctr, Ds |
| 12 | Naringenin chalcone glucoside | 11.3 | 433.1138 | 271.06, 151.00, 119.05 | C21H22O10 | −0.46 | Ctr, Ds |
| 14 | Naringenin d | 12.8 | 271.0611 | 151.00, 119.05 | C15H12O5 | −0.37 | Ctr, Ds |
| 15 | Naringenin calchone | 13.4 | 271.0611 | 151.00, 119.05 | C15H12O5 | −0.37 | Ctr, Ds |
| Peak a | Compound | RED-Ctr (µg/100 g FW ± SD) | RED-Ds (µg/100 g FW ± SD) |
|---|---|---|---|
| Phenolic acids | |||
| 1a + 1b | Caffeic acid glucoside (isomers I and II) | 8.18 ± 0.1 | 18.6 ± 0.1 * |
| 2a + 2b | p-Coumaric acid glucoside (isomers I and II) | 12.8 ± 0.1 | 37.8 ± 0.1 * |
| 3a + 3b | Chlorogenic acid (isomers I and II) | 46.9 ± 0.5 | 51.9 ± 0.9 * |
| 4a + 4b | p-Coumaric acid glucoside (isomers III and IV) | 9.13 ± 0.1 | 16.3 ± 0.2 * |
| 5a + 5b | Caffeoylquinic acid (isomers I and II) | 15.0 ± 0.2 | 18.3 ± 0.2 * |
| 11 | Dicaffeoylquinic acid (isomer I) | 16.5 ± 0.4 | 20.5 ± 0.4 * |
| 13 | Dicaffeoylquinic acid (isomer II) | 3.47 ± 0.06 | 5.03 ± 0.1 * |
| Flavonoids | |||
| 6 | Quercetin 3-O-(2″-O-apiofuranosyl-6″-O-rhamnopyranosyl-glucopyranoside) | 16.1 ± 0.4 | 16.7 ± 0.4 |
| 7 | Rutin | 45.5 ± 0.5 | 62.3 ± 0.9 * |
| 8 | Kaempferol rutinoside-pentoside | 4.88 ± 0.3 | 5.04 ± 0.2 |
| 9 | Kaempferol 3-O-rutinoside | 7.23 ± 0.3 | 8.17 ± 0.2 * |
| 10 | Naringenin 7-O-glucoside | 103 ± 4 * | 90.0 ± 2 |
| 12 | Naringenin chalcone glucoside | 119 ± 1 * | 109 ± 1 |
| 14 | Naringenin | 793 ± 19 * | 556 ± 2 |
| 15 | Naringenin chalcone | 77.4 ± 2 * | 58.0 ± 0.6 |
| Total phenolic acids | 112 ± 1 | 168 ± 2 * | |
| Total flavonoids | 1166 ± 27 * | 905 ± 7 | |
| Total phenols | 1278 ± 28 * | 1073 ± 9 |
| Gene | Sequence | GenBank, Accession | Length (bp) | Temperature (°C) | |
|---|---|---|---|---|---|
| AKT | Forward | CTGCACAAACGAGGGGAGTA | NM_001014431.2 | 142 | 60 |
| Reverse | GCGCCACAGAGAAGTTGTTG | ||||
| B2M | Forward | CACTGAATTCACCCCCACTGA | NM_004048.4 | 102 | 60 |
| Reverse | GCTTACATGTCTCGATCCCAC | ||||
| eEF1A | Forward | CTTTGGGTCGCTTTGCTGTT | NM_001402 | 183 | 60 |
| Reverse | CCGTTCTTCCACCACTGATT | ||||
| FOXO1 | Forward | GGGTTAGTGAGCAGGTTACAC | NM_002015.4 | 170 | 60 |
| Reverse | CTTTGCTGCCAAGTCTGACG | ||||
| MYH2 | Forward | CTCAAAGCTCTCTGCTACCCC | NM_017534.6 | 88 | 60 |
| Reverse | CTACTGCGTTGGACACCTGTTCT | ||||
| MYOG | Forward | AGATTGTCTTCCAAGCCGGG | NM_002479.6 | 112 | 60 |
| Reverse | CTGGCTTCCTAGCATCAGGG | ||||
| RPL13A | Forward | CGCCCTACGACAAGAAAAAG | NM_012423 | 206 | 60 |
| Reverse | CCGTAGCCTCATGAGCTGTT | ||||
| TNNT1 | Forward | GTCAGAGAGAGCCGAGCAAC | NM_001126133.3 | 197 | 60 |
| Reverse | CACGCTTCTGTTCTGCCTTG | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Felice, F.; Cesare, M.M.; Fredianelli, L.; De Leo, M.; Conti, V.; Braca, A.; Di Stefano, R. Effect of Tomato Peel Extract Grown under Drought Stress Condition in a Sarcopenia Model. Molecules 2022, 27, 2563. https://doi.org/10.3390/molecules27082563
Felice F, Cesare MM, Fredianelli L, De Leo M, Conti V, Braca A, Di Stefano R. Effect of Tomato Peel Extract Grown under Drought Stress Condition in a Sarcopenia Model. Molecules. 2022; 27(8):2563. https://doi.org/10.3390/molecules27082563
Chicago/Turabian StyleFelice, Francesca, Maria Michela Cesare, Luca Fredianelli, Marinella De Leo, Veronica Conti, Alessandra Braca, and Rossella Di Stefano. 2022. "Effect of Tomato Peel Extract Grown under Drought Stress Condition in a Sarcopenia Model" Molecules 27, no. 8: 2563. https://doi.org/10.3390/molecules27082563
APA StyleFelice, F., Cesare, M. M., Fredianelli, L., De Leo, M., Conti, V., Braca, A., & Di Stefano, R. (2022). Effect of Tomato Peel Extract Grown under Drought Stress Condition in a Sarcopenia Model. Molecules, 27(8), 2563. https://doi.org/10.3390/molecules27082563