
In Vitro Antioxidant and Anticancer Properties of Various E. senegalensis Extracts

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Extracts Preparation and Their Chemical Class Composition
2.2. In Vitro Growth Inhibitory Effects of the Extracts Assessed by MTT Assay
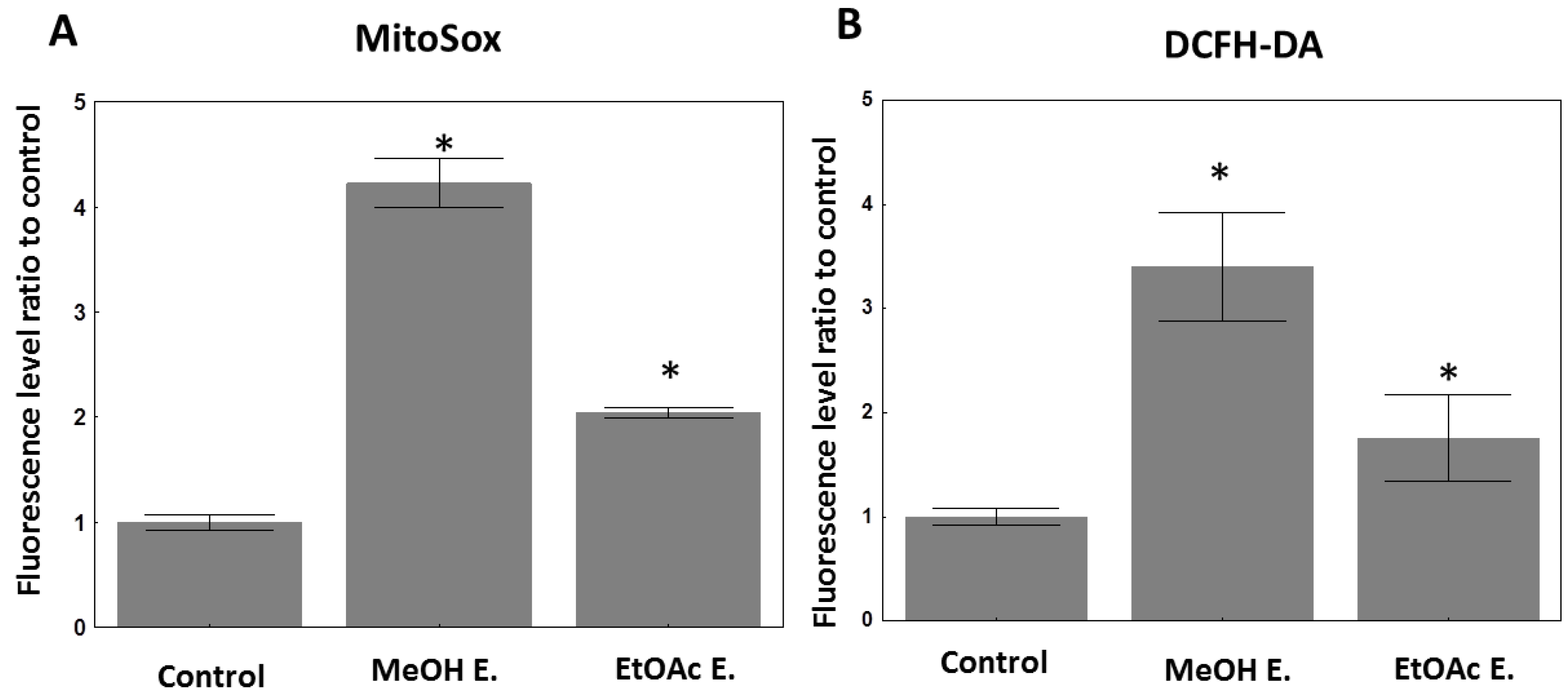
2.3. Antioxidant Effects of the Extracts from E. senegalensis
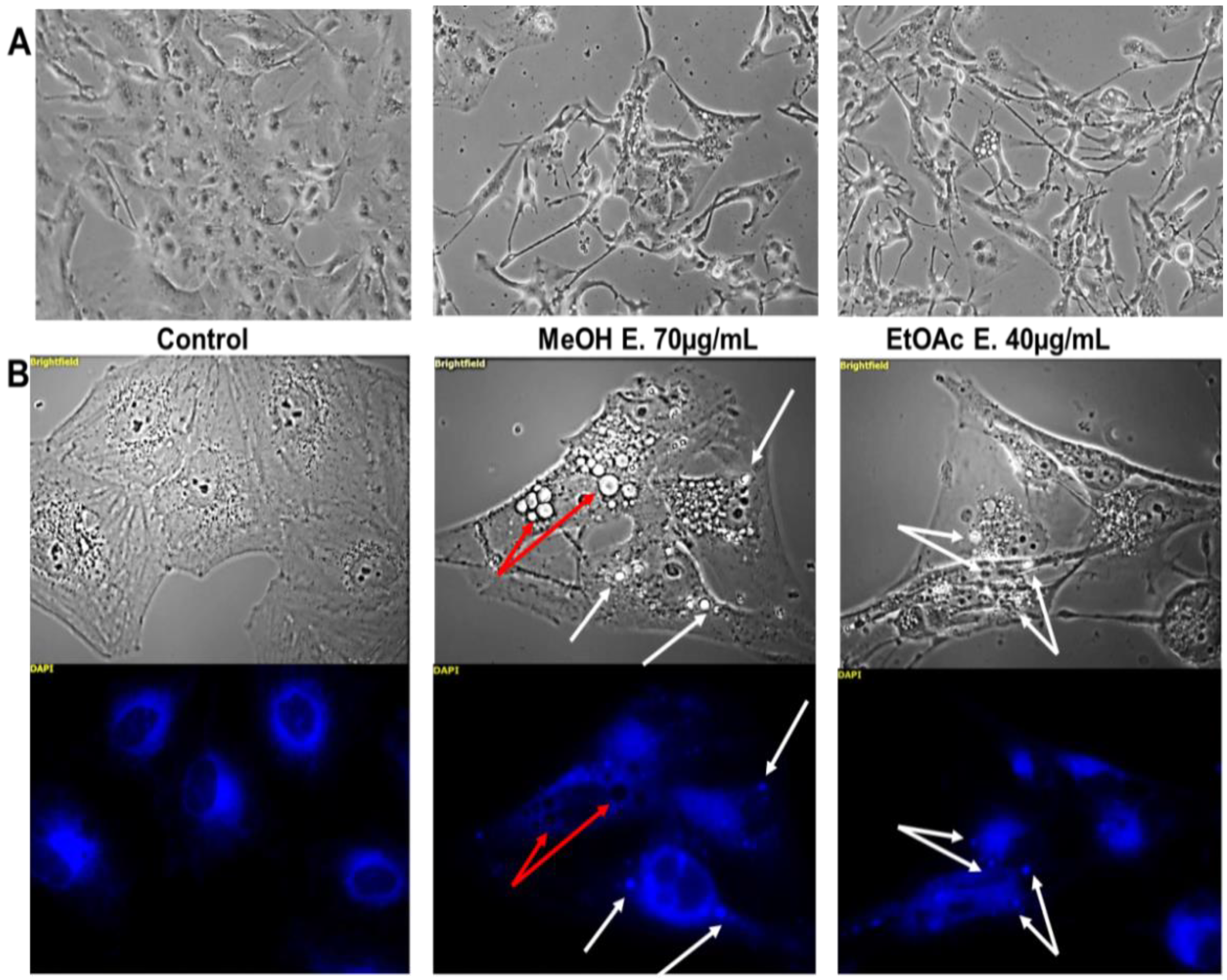
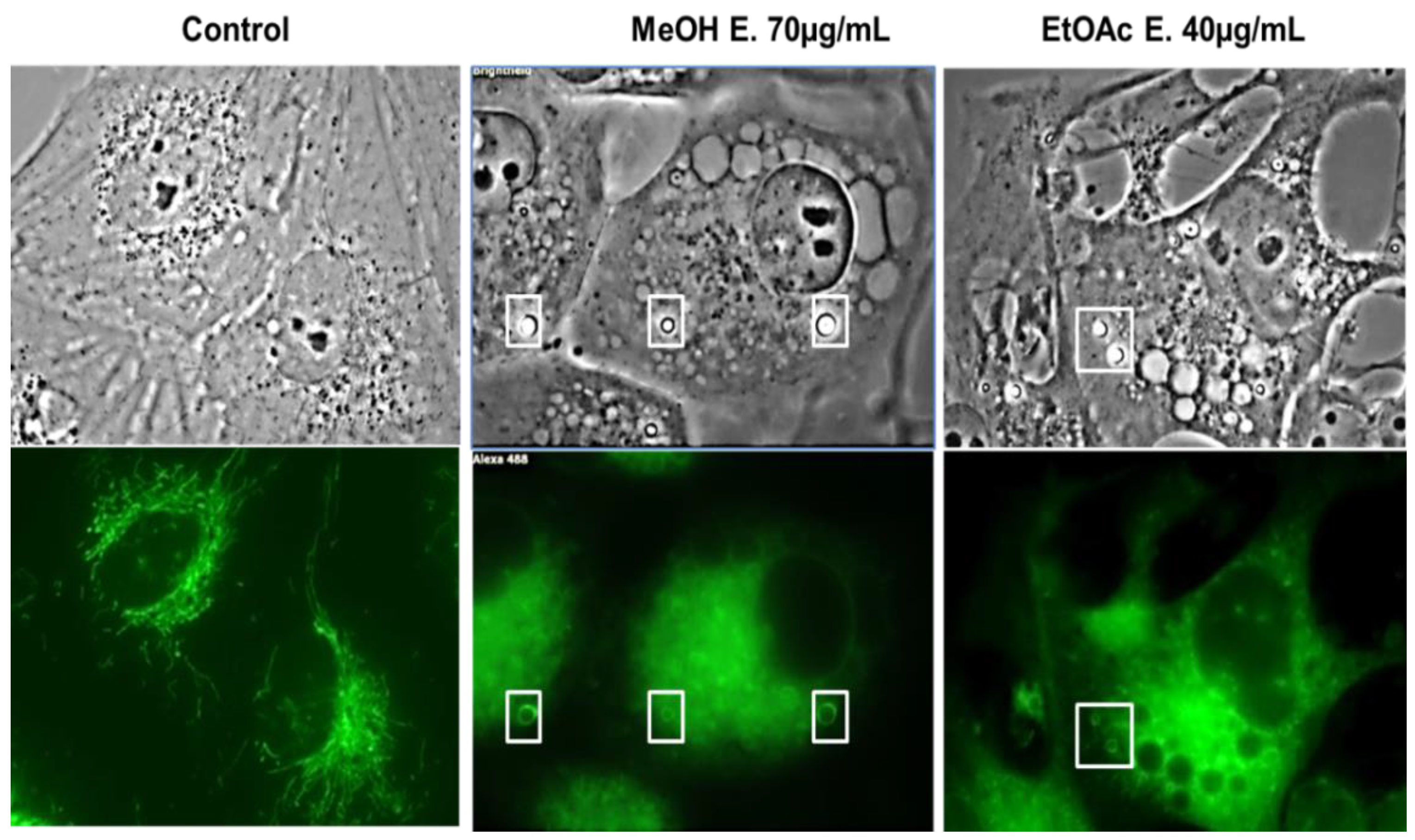
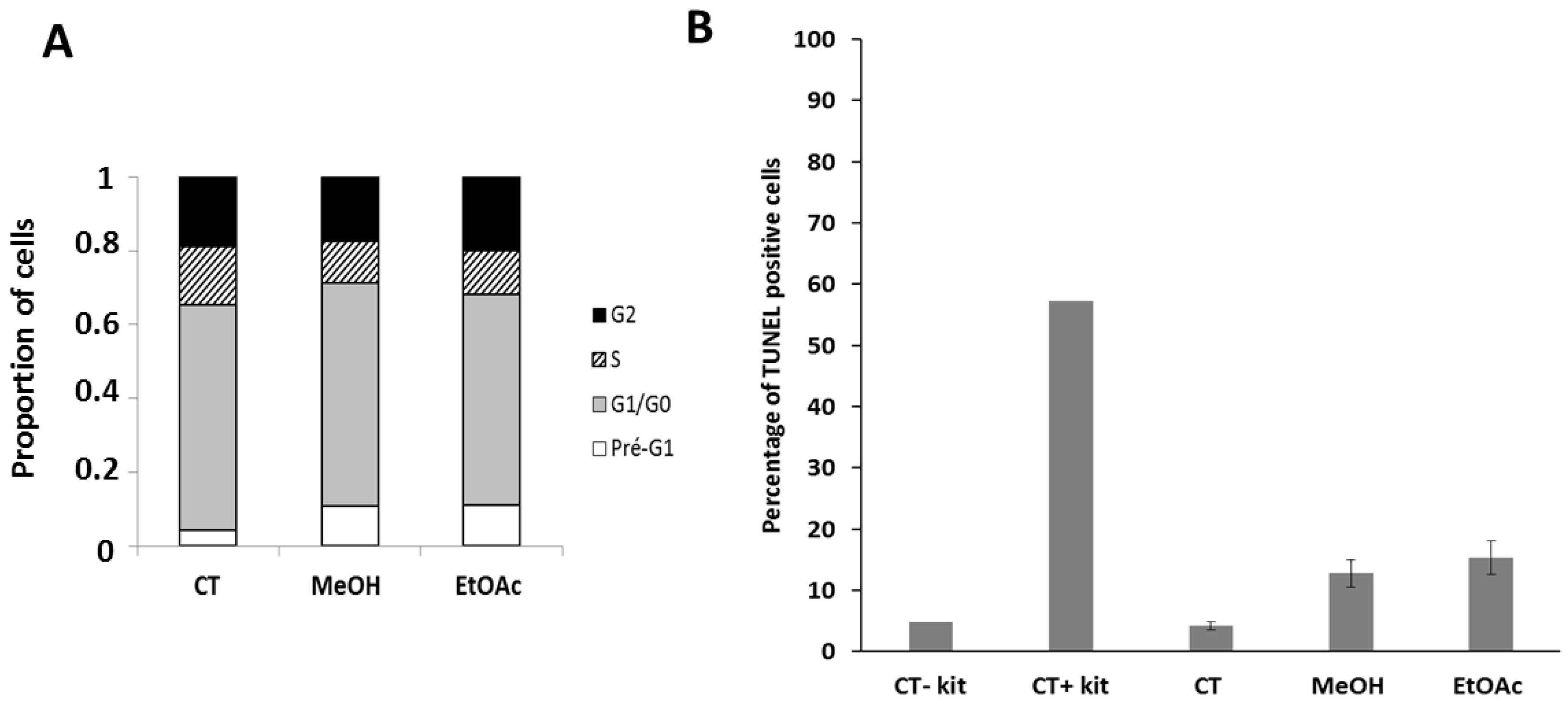
2.4. E. senegalensis Extracts Induce Morphological Changes including Vacuolization of Cancer Cells
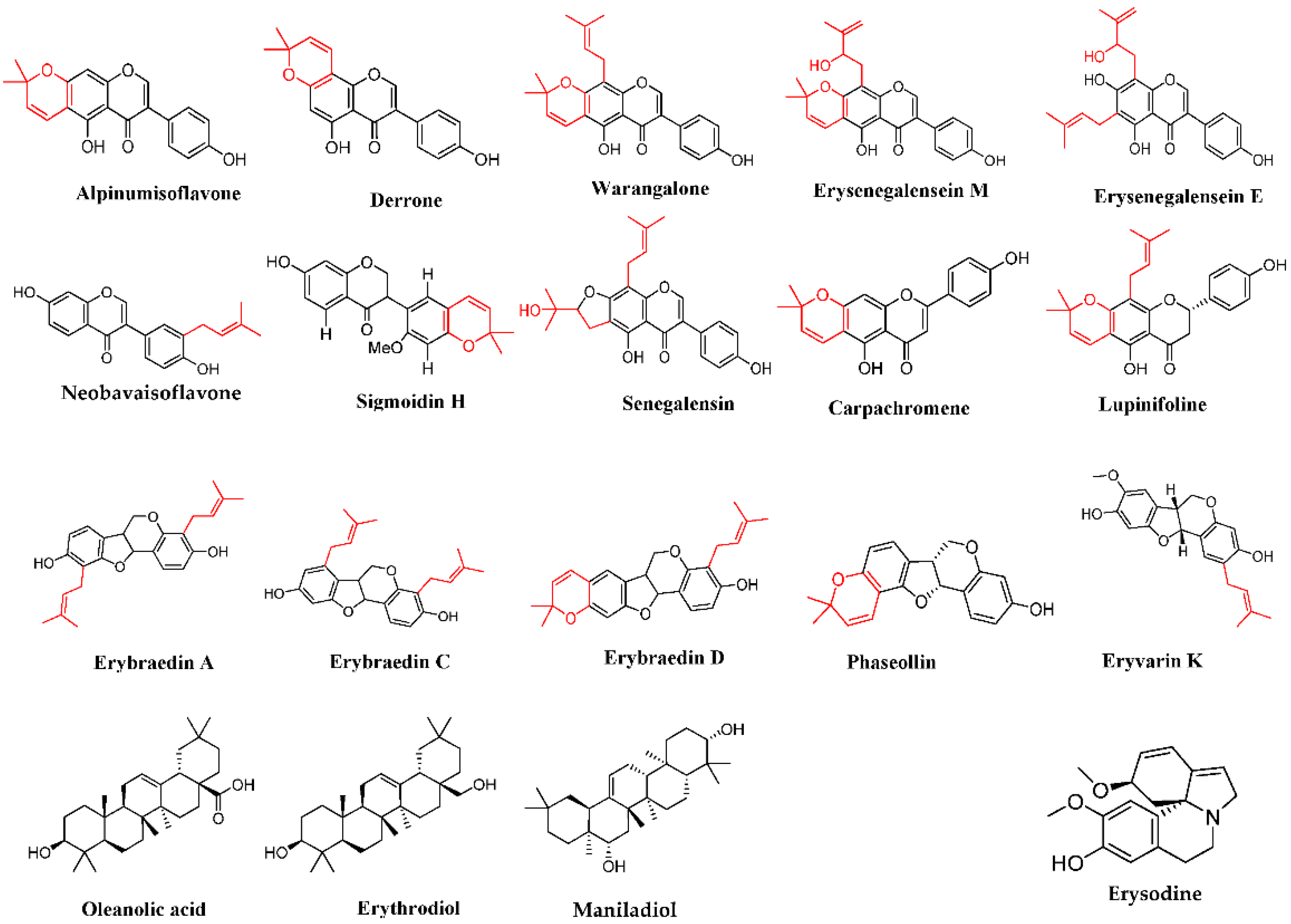
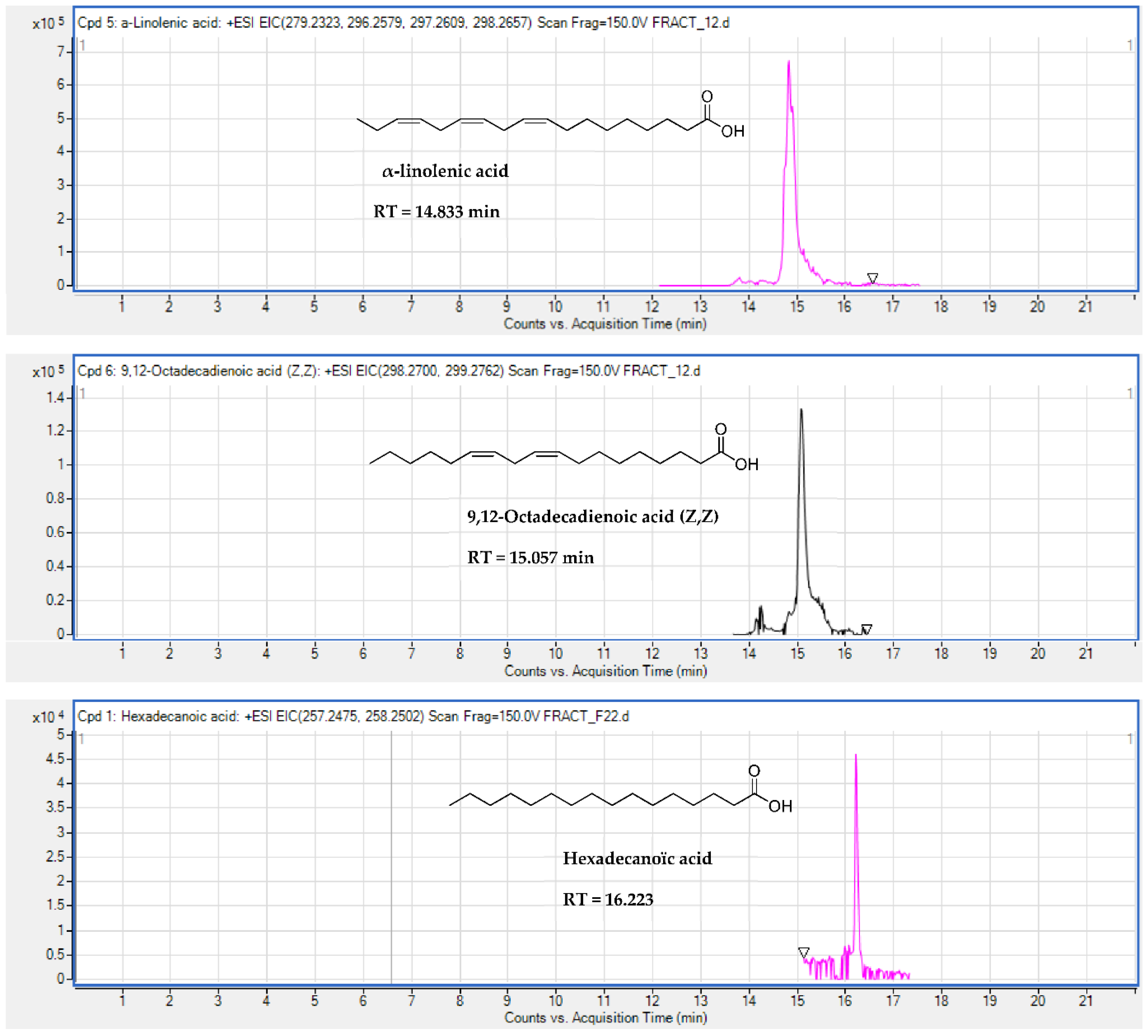
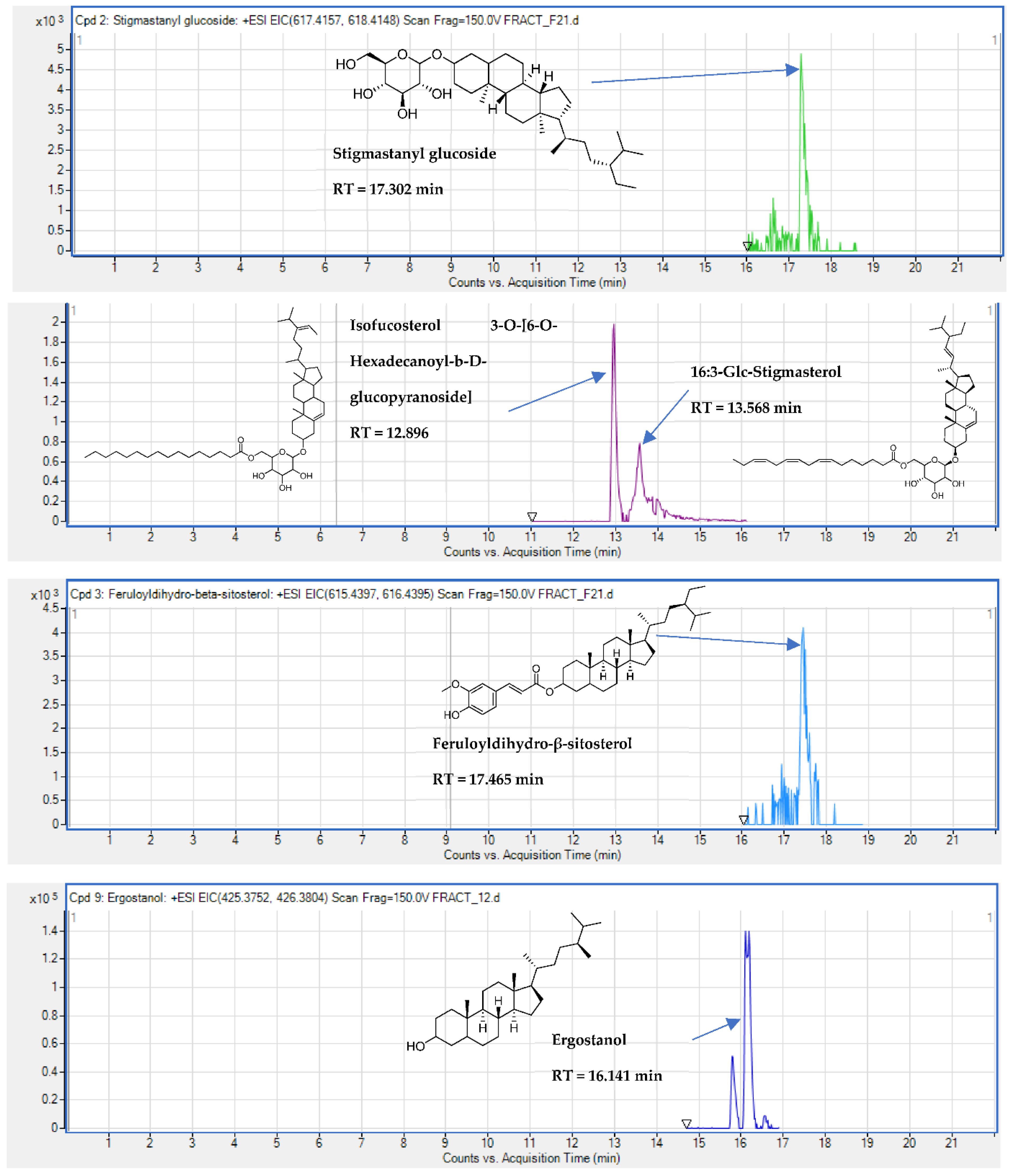
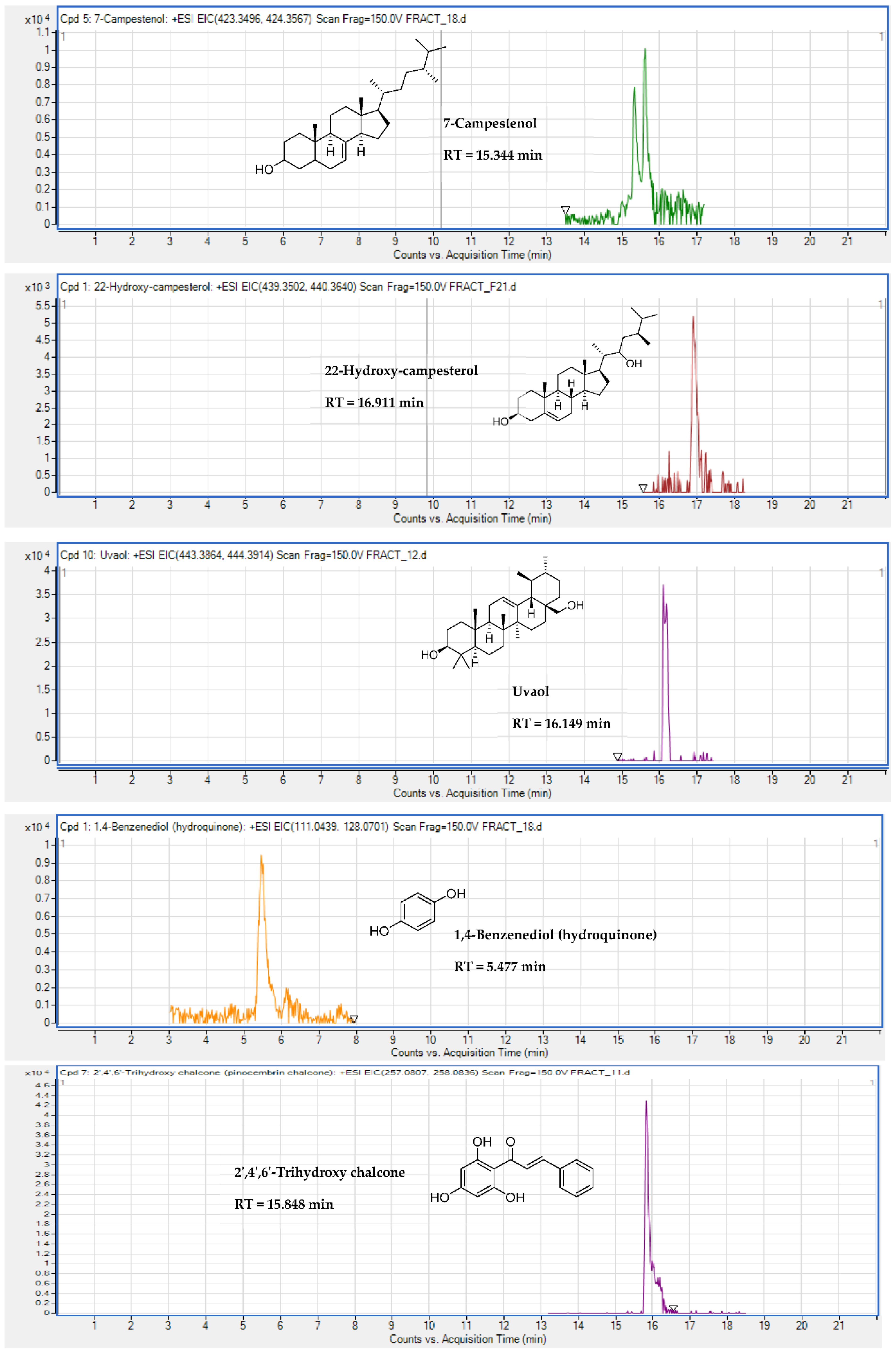
2.5. Identification of Substances of E. senegalensis Extracts by LC-HRMS
3. Discussion
4. Materials and Methods
4.1. Chemistry
4.1.1. Plant Material
4.1.2. Reagents and Solvents
4.1.3. Extraction Method
4.1.4. Phytochemicals Characterization
4.1.5. Fractionation Method
4.1.6. LC-MS Process, Data Acquisition and Analysis
4.2. Activity Assays
4.2.1. Antioxidant Activity Evaluation
4.2.2. Culture Media and Cancer Cell Lines
4.2.3. MTT Colorimetric Assay
4.2.4. Phase Contrast Microscopy
4.2.5. Fluorescent Microscopy for Vacuole Characterization
4.2.6. Effects of the Extracts on the Cell Cycle and Apoptosis
4.2.7. Evaluation of Cellular ROS Species under E. senegalensis Treatment
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Appendix A



Appendix B


References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the Global Cancer Incidence and Mortality in 2018: GLOBOCAN Sources and Methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, R.M.; Muqbil, I.; Lowe, L.; Yedjou, C.; Hsu, H.-Y.; Lin, L.-T.; Siegelin, M.D.; Fimognari, C.; Kumar, N.B.; Dou, Q.P.; et al. Broad Targeting of Resistance to Apoptosis in Cancer. Semin. Cancer Biol. 2015, 35, S78–S103. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Tavakol, S.; Zarrabi, A.; Ashrafizadeh, M. Dual Role of Quercetin in Enhancing the Efficacy of Cisplatin in Chemotherapy and Protection against Its Side Effects: A Review. Arch. Physiol. Biochem. 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.-F.; Han, X.-Y.; Sun, Y.-S.; Zhang, L.-X.; Liu, W.; Liu, X.-X.; Li, W.; Liu, Y.-Y. Kidney Protection Effect of Ginsenoside Re and Its Underlying Mechanisms on Cisplatin-Induced Kidney Injury. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 48, 2219–2229. [Google Scholar] [CrossRef] [PubMed]
- Bayala, B.; Nadembega, C.; Guenné, S.; Buñay, J.; Mahoukèdè Zohoncon, T.; Wendkuuni Djigma, F.; Yonli, A.; Baron, S.; Figueredo, G.; A Lobaccaro, J.-M.; et al. Chemical Composition, Antioxidant and Cytotoxic Activities of Hyptis Suaveolens (L.) Poit. Essential Oil on Prostate and Cervical Cancers Cells. Pak. J. Biol. Sci. 2020, 23, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Sawadogo, W.R.; Luo, Y.; Elkington, B.; He, T.-C.; Wang, C.-Z.; Yuan, C.-S. Cytotoxicity and Preliminary Analysis of the Pro-Apoptotic and Cell Cycle Arrest Effects of Lantana Ukambensis Against Colorectal Cancer Cells. Int. J. Appl. Biol. Pharm. Technol. 2020, 11, 170–187. [Google Scholar]
- Sawadogo, W.R.; Schumacher, M.; Teiten, M.-H.; Dicato, M.; Diederich, M. Traditional West African Pharmacopeia, Plants and Derived Compounds for Cancer Therapy. Biochem. Pharmacol. 2012, 84, 1225–1240. [Google Scholar] [CrossRef]
- Togola, A.; Austarheim, I.; Theïs, A.; Diallo, D.; Paulsen, B.S. Ethnopharmacological Uses of Erythrina Senegalensis: A Comparison of Three Areas in Mali, and a Link between Traditional Knowledge and Modern Biological Science. J. Ethnobiol. Ethnomed. 2008, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Kuete, V.; Sandjo, L.P.; Kwamou, G.M.N.; Wiench, B.; Nkengfack, A.E.; Efferth, T. Activity of Three Cytotoxic Isoflavonoids from Erythrina Excelsa and Erythrina Senegalensis (Neobavaisoflavone, Sigmoidin H and Isoneorautenol) toward Multi-Factorial Drug Resistant Cancer Cells. Phytomed. Int. J. Phytother. Phytopharm. 2014, 21, 682–688. [Google Scholar] [CrossRef]
- Bilanda, D.C.; Bidingha, R.À.G.; Dzeufiet, P.D.D.; Fouda, Y.B.; Ngapout, R.F.; Tcheutchoua, Y.; Owona, P.E.; Wouamba, S.C.N.; Tatchou, L.T.; Dimo, T.; et al. Antihypertensive and Antidiabetic Activities of Erythrina Senegalensis DC (Fabaceae) Stem Bark Aqueous Extract on Diabetic Hypertensive Rats. J. Ethnopharmacol. 2020, 246, 112200. [Google Scholar] [CrossRef] [PubMed]
- Saidu, K.; Onah, J.; Orisadipe, A.; Olusola, A.; Wambebe, C.; Gamaniel, K. Antiplasmodial, Analgesic, and Anti-Inflammatory Activities of the Aqueous Extract of the Stem Bark of Erythrina Senegalensis. J. Ethnopharmacol. 2000, 71, 275–280. [Google Scholar] [CrossRef]
- Wanjala, C.C.; Juma, B.F.; Bojase, G.; Gashe, B.A.; Majinda, R.R. Erythrinaline Alkaloids and Antimicrobial Flavonoids from Erythrina Latissima. Planta Med. 2002, 68, 640–642. [Google Scholar] [CrossRef] [PubMed]
- Iinuma, M.; Okawa, Y.; Tanaka, T.; Ho, F.C.; Kobayashi, Y.; Miyauchi, K.I. Anti-Oral Microbial Activity of Isoflavonoids in Root Bark of Ormosia Monosperma. Phytochemistry 1994, 37, 889–891. [Google Scholar] [CrossRef]
- Meragelman, K.M.; McKee, T.C.; Boyd, M.R. Anti-HIV Prenylated Flavonoids from Monotes Africanus. J. Nat. Prod. 2001, 64, 546–548. [Google Scholar] [CrossRef]
- Lyddiard, J.R.A.; Whitfield, P.J.; Bartlett, A. Antischistosomal Bioactivity of Isoflavonoids from Millettia Thonningii (Leguminosae). J. Parasitol. 2002, 88, 163–170. [Google Scholar] [CrossRef]
- Dramane, S.; Koné, M.; Kamanzi, K. Evaluation of Antimicrobial and Free Radical Scavenging Activities of Some Bioactif Taxons from Côte d’Ivoire. Eur. J. Sci. Res. 2010, 40, 307–317. [Google Scholar]
- Niwa, T.; Murakami, K.; Ohtake, T.; Etoh, H.; Shimizu, A.; Shimizu, Y.; Kato, Y.; Tanaka, H. Peroxynitrite Scavenging Activities of Aromatic Compounds Isolated from Konnyaku, Amorphophallus Konjac K.Koch. Biosci. Biotechnol. Biochem. 2002, 66, 1386–1388. [Google Scholar] [CrossRef]
- Togola, A.; Hedding, B.; Theis, A.; Wangensteen, H.; Rise, F.; Smestad Paulsen, B.; Diallo, D.; Egil Malterud, K. 15-Lipoxygenase Inhibitory Effects of Prenylated Flavonoids from Erythrina Senegalensis. Planta Med. 2009, 75, 1168–1170. [Google Scholar] [CrossRef]
- Oh, W.K.; Lee, C.-H.; Seo, J.H.; Chung, M.Y.; Cui, L.; Fomum, Z.T.; Kang, J.S.; Lee, H.S. Diacylglycerol Acyltransferase-Inhibitory Compounds from Erythrina Senegalensis. Arch. Pharm. Res. 2009, 32, 43–47. [Google Scholar] [CrossRef]
- Majeed, I.; Rizwan, K.; Ashar, A.; Rasheed, T.; Amarowicz, R.; Kausar, H.; Zia-Ul-Haq, M.; Marceanu, L.G. A Comprehensive Review of the Ethnotraditional Uses and Biological and Pharmacological Potential of the Genus Mimosa. Int. J. Mol. Sci. 2021, 22, 7463. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, K.; Majeed, I.; Bilal, M.; Rasheed, T.; Shakeel, A.; Iqbal, S. Phytochemistry and Diverse Pharmacology of Genus Mimosa: A Review. Biomolecules 2022, 12, 83. [Google Scholar] [CrossRef]
- Fofana, S.; Ouédraogo, M.; Esposito, R.C.; Ouedraogo, W.P.; Delporte, C.; Van Antwerpen, P.; Mathieu, V.; Guissou, I.P. Systematic Review of Potential Anticancerous Activities of Erythrina Senegalensis DC (Fabaceae). Plants 2021, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zeng, X.; Wu, X. Effect of Oleanolic Acid on Apoptosis and Autophagy of SMMC-7721 Hepatoma Cells. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e921606. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Li, X.; Yu, S.; Liang, L. Inhibition of Cancer Cell Growth by Oleanolic Acid in Multidrug Resistant Liver Carcinoma Is Mediated via Suppression of Cancer Cell Migration and Invasion, Mitochondrial Apoptosis, G2/M Cell Cycle Arrest and Deactivation of JNK/P38 Signalling Pathway. J. BUON 2019, 24, 1964–1969. [Google Scholar] [PubMed]
- Juan, M.E.; Wenzel, U.; Daniel, H.; Planas, J.M. Erythrodiol, a Natural Triterpenoid from Olives, Has Antiproliferative and Apoptotic Activity in HT-29 Human Adenocarcinoma Cells. Mol. Nutr. Food Res. 2008, 52, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Namkoong, S.; Kim, T.-J.; Jang, I.-S.; Kang, K.-W.; Oh, W.-K.; Park, J. Alpinumisoflavone Induces Apoptosis and Suppresses Extracellular Signal-Regulated Kinases/Mitogen Activated Protein Kinase and Nuclear Factor-ΚB Pathways in Lung Tumor Cells. Biol. Pharm. Bull. 2011, 34, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Li, X.; Li, G.; Meng, Y.; Jin, Y.; Shang, S.; Li, Y. Alpinumisoflavone Causes DNA Damage in Colorectal Cancer Cells via Blocking DNA Repair Mediated by RAD51. Life Sci. 2019, 216, 259–270. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, H.; Sun, M.; He, T.; Liu, Y.; Yang, X.; Shi, X.; Liu, X. Alpinumisoflavone Suppresses Hepatocellular Carcinoma Cell Growth and Metastasis via NLRP3 Inflammasome-Mediated Pyroptosis. Pharmacol. Rep. 2020, 72, 1370–1382. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Chang, Y.; Wang, X.; Ban, C.; Zhang, F. Reduction of COX-2 through Modulating MiR-124/SPHK1 Axis Contributes to the Antimetastatic Effect of Alpinumisoflavone in Melanoma. Am. J. Transl. Res. 2017, 9, 986–998. [Google Scholar]
- Hoang, N.T.M.; Phuong, T.T.; Nguyen, T.T.N.; Tran, Y.T.H.; Nguyen, A.T.N.; Nguyen, T.L.; Bui, K.T.V. In Vitro Characterization of Derrone as an Aurora Kinase Inhibitor. Biol. Pharm. Bull. 2016, 39, 935–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.-J.; Kim, S.-Y.; Kwon, E.-B.; Jo, Y.H.; Lee, M.K.; Lee, H.-S.; Moon, D.-O.; Kim, M.-O. Derrone Induces Autophagic Cell Death through Induction of ROS and ERK in A549 Cells. PLoS ONE 2019, 14, e0218659. [Google Scholar] [CrossRef] [PubMed]
- Ito, C.; Murata, T.; Itoigawa, M.; Nakao, K.; Kumagai, M.; Kaneda, N.; Furukawa, H. Induction of Apoptosis by Isoflavonoids from the Leaves of Millettia Taiwaniana in Human Leukemia HL-60 Cells. Planta Med. 2006, 72, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, H.-Y.; Jung, Y.; Park, K.H.; Oh, W.K.; Lee, H.-Y. Erybraedin A Is a Potential Src Inhibitor That Blocks the Adhesion and Viability of Non-Small-Cell Lung Cancer Cells. Biochem. Biophys. Res. Commun. 2018, 502, 145–151. [Google Scholar] [CrossRef]
- Maurich, T.; Iorio, M.; Chimenti, D.; Turchi, G. Erybraedin C and Bitucarpin A, Two Structurally Related Pterocarpans Purified from Bituminaria Bituminosa, Induced Apoptosis in Human Colon Adenocarcinoma Cell Lines MMR- and P53-Proficient and -Deficient in a Dose-, Time-, and Structure-Dependent Fashion. Chem. Biol. Interact. 2006, 159, 104–116. [Google Scholar] [CrossRef]
- Rukachaisirikul, T.; Saekee, A.; Tharibun, C.; Watkuolham, S.; Suksamrarn, A. Biological Activities of the Chemical Constituents of Erythrina Stricta and Erythrina Subumbrans. Arch. Pharm. Res. 2007, 30, 1398–1403. [Google Scholar] [CrossRef]
- Wätjen, W.; Kulawik, A.; Suckow-Schnitker, A.K.; Chovolou, Y.; Rohrig, R.; Ruhl, S.; Kampkötter, A.; Addae-Kyereme, J.; Wright, C.W.; Passreiter, C.M. Pterocarpans Phaseollin and Neorautenol Isolated from Erythrina Addisoniae Induce Apoptotic Cell Death Accompanied by Inhibition of ERK Phosphorylation. Toxicology 2007, 242, 71–79. [Google Scholar] [CrossRef]
- Nie, H.; Wang, Y.; Qin, Y.; Gong, X.-G. Oleanolic Acid Induces Autophagic Death in Human Gastric Cancer Cells In Vitro and In Vivo. Cell Biol. Int. 2016, 40, 770–778. [Google Scholar] [CrossRef]
- Hosny, S.; Sahyon, H.; Youssef, M.; Negm, A. Oleanolic Acid Suppressed DMBA-Induced Liver Carcinogenesis through Induction of Mitochondrial-Mediated Apoptosis and Autophagy. Nutr. Cancer 2021, 73, 968–982. [Google Scholar] [CrossRef]
- Liu, J.; Zheng, L.; Ma, L.; Wang, B.; Zhao, Y.; Wu, N.; Liu, G.; Lin, X. Oleanolic Acid Inhibits Proliferation and Invasiveness of Kras-Transformed Cells via Autophagy. J. Nutr. Biochem. 2014, 25, 1154–1160. [Google Scholar] [CrossRef]
- Sheu, Y.-W.; Chiang, L.-C.; Chen, I.-S.; Chen, Y.-C.; Tsai, I.-L. Cytotoxic Flavonoids and New Chromenes from Ficus Formosana f. Formosana. Planta Med. 2005, 71, 1165–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukiya, M.; Akihisa, T.; Tokuda, H.; Suzuki, H.; Mukainaka, T.; Ichiishi, E.; Yasukawa, K.; Kasahara, Y.; Nishino, H. Constituents of Compositae Plants III. Anti-Tumor Promoting Effects and Cytotoxic Activity against Human Cancer Cell Lines of Triterpene Diols and Triols from Edible Chrysanthemum Flowers. Cancer Lett. 2002, 177, 7–12. [Google Scholar] [CrossRef]
- Ito, A.; Chai, H.-B.; Kardono, L.B.S.; Setowati, F.M.; Afriastini, J.J.; Riswan, S.; Farnsworth, N.R.; Cordell, G.A.; Pezzuto, J.M.; Swanson, S.M.; et al. Saponins from the Bark of Nephelium Maingayi. J. Nat. Prod. 2004, 67, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Ateba, S.B.; Mvondo, M.A.; Djiogue, S.; Zingué, S.; Krenn, L.; Njamen, D. A Pharmacological Overview of Alpinumisoflavone, a Natural Prenylated Isoflavonoid. Front. Pharmacol. 2019, 10, 952. [Google Scholar] [CrossRef]
- Tjahjandarie, T.S.; Tanjung, M. Phenolic Compounds from the Stem Bark of Erythrina Orientalis and Their Cytotoxic and Antioxidant Activities. Pharma Chem. 2015, 7, 206–211. [Google Scholar]
- Akter, K.; Barnes, E.C.; Loa-Kum-Cheung, W.L.; Yin, P.; Kichu, M.; Brophy, J.J.; Barrow, R.A.; Imchen, I.; Vemulpad, S.R.; Jamie, J.F. Antimicrobial and Antioxidant Activity and Chemical Characterisation of Erythrina Stricta Roxb. (Fabaceae). J. Ethnopharmacol. 2016, 185, 171–181. [Google Scholar] [CrossRef]
- Fu, G.; Li, W.; Huang, X.; Zhang, R.; Tian, K.; Hou, S.; Li, Y. Antioxidant and Alpha-Glucosidase Inhibitory Activities of Isoflavonoids from the Rhizomes of Ficus Tikoua Bur. Nat. Prod. Res. 2018, 32, 399–405. [Google Scholar] [CrossRef]
- Bai, X.; Lai, T.; Zhou, T.; Li, Y.; Li, X.; Zhang, H. In Vitro Antioxidant Activities of Phenols and Oleanolic Acid from Mango Peel and Their Cytotoxic Effect on A549 Cell Line. Mol. J. Synth. Chem. Nat. Prod. Chem. 2018, 23, 1395. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Wang, J.; Zhang, C.; Yu, Y.; Cai, H. Proapoptotic Protein Smac Mediates Apoptosis in Ovarian Cancer Cells When Treated with Carpachromene. Arch. Med. Sci. 2021, 17, 1–14. [Google Scholar] [CrossRef]
- Issarachot, P.; Sangkaew, W.; Sianglum, W.; Saeloh, D.; Limsuwan, S.; Voravuthikunchai, S.P.; Joycharat, N. α-Glucosidase Inhibitory, Antibacterial, and Antioxidant Activities of Natural Substances from the Wood of Derris Reticulata Craib. Nat. Prod. Res. 2021, 35, 2858–2865. [Google Scholar] [CrossRef]
- Allouche, Y.; Beltrán, G.; Gaforio, J.J.; Uceda, M.; Mesa, M.D. Antioxidant and Antiatherogenic Activities of Pentacyclic Triterpenic Diols and Acids. Food Chem. Toxicol. 2010, 48, 2885–2890. [Google Scholar] [CrossRef] [PubMed]
- Allouche, Y.; Warleta, F.; Campos, M.; Sánchez-Quesada, C.; Uceda, M.; Beltrán, G.; Gaforio, J.J. Antioxidant, Antiproliferative, and pro-Apoptotic Capacities of Pentacyclic Triterpenes Found in the Skin of Olives on MCF-7 Human Breast Cancer Cells and Their Effects on DNA Damage. J. Agric. Food Chem. 2011, 59, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Ibeas, E.; Carvalho-Tavares, J.; Hernández, M.; Ruiz-Gutierrez, V.; Nieto, M.L. Natural Triterpenic Diols Promote Apoptosis in Astrocytoma Cells through ROS-Mediated Mitochondrial Depolarization and JNK Activation. PLoS ONE 2009, 4, e5975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaunig, J.E. Oxidative Stress and Cancer 2018. Curr. Pharm. Des. 2018, 24, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Mileo, A.M.; Miccadei, S. Polyphenols as Modulator of Oxidative Stress in Cancer Disease: New Therapeutic Strategies. Oxid. Med. Cell. Longev. 2016, 2016, 6475624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.K.; Lee, S.B.; Won, J.; Choi, H.Y.; Kim, K.; Yang, G.-M.; Dayem, A.A.; Cho, S.-G. Correlation between Oxidative Stress, Nutrition, and Cancer Initiation. Int. J. Mol. Sci. 2017, 18, 1544. [Google Scholar] [CrossRef] [Green Version]
- Pilco-Ferreto, N.; Calaf, G.M. Influence of Doxorubicin on Apoptosis and Oxidative Stress in Breast Cancer Cell Lines. Int. J. Oncol. 2016, 49, 753–762. [Google Scholar] [CrossRef] [Green Version]
- Fofana, S.; Gnoula, C.; Ouedraogo Moussa, M.; Palé, E.; Nebié, R.H.; Nikiema, J.-B.; Guissou, I.P.; Simporé, J. DPPH Radical Scavenging and Lipoxygenase Inhibitory Effects in Extracts from Erythrina Senegalensis (Fabaceae) DC. Afr. J. Pharm. Pharmacol. 2016, 10, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.V.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress in Infection and Consequent Disease. Oxid. Med. Cell. Longev. 2017, 2017, 3496043. [Google Scholar] [CrossRef]
- Vasquez, M.; Zuniga, M.; Rodriguez, A. Oxidative Stress and Pathogenesis in Malaria. Front. Cell. Infect. Microbiol. 2021, 11, 768182. [Google Scholar] [CrossRef]
- Nacoulma, O.G. Plantes Médicinales et Pratiques Médicales Traditionnelles Au Burkina Faso: Cas Du Plateau Central. Fac. Sci. Tech. Univ. Ouagadougou 1996, 320, 42–53. [Google Scholar]
- Wandji, J.; Nkengfack, A.E.; Fomum, Z.T.; Ubillas, R.; Killday, K.B.; Tempesta, M.S. A New Prenylated Isoflavone and Long Chain Esters from Two Erythrina Species. J. Nat. Prod. 1990, 53, 1425–1429. [Google Scholar] [CrossRef] [PubMed]
- Wandji, J.; Fomum, Z.T.; Tillequin, F.; Baudouin, G.; Koch, M. Epoxyisoflavones from Erythrina Senegalensis. Phytochemistry 1994, 35, 1573–1577. [Google Scholar] [CrossRef]
- Wandji, J.; Fomum, Z.T.; Tillequin, F.; Libot, F.; Koch, M. Erysenegalenseins B and C, Two New Prenylated Isoflavanones from Erythrina Senegalensis. J. Nat. Prod. 1995, 58, 105–108. [Google Scholar] [CrossRef]
- Wandji, J.; Fomum, Z.T.; Tillequin, F.; Skaltsounis, A.L.; Koch, M. Erysenegalenseins H and I: Two New Isoflavones from Erythrina Senegalensis1. Planta Med. 1994, 60, 178–180. [Google Scholar] [CrossRef]
- Fomum, Z.T.; Ayafor, J.F.; Wandji, J. Erythrisenegalone, a Prenylated-Flavanone from Erythrina Senegalensis. Phytochemistry 1985, 24, 3075–3076. [Google Scholar] [CrossRef]
- Wandji, J.; Awanchiri, S.; Fomum, Z.T.; Tillequin, F.; Libot, F. Isoflavones and Alkaloids from the Stem Bark and Seeds of Erythrina Senegalensis. Phytochemistry 1995, 39, 677–681. [Google Scholar] [CrossRef]
- Oh, W.K.; Lee, H.S.; Ahn, S.C.; Ahn, J.S.; Mbafor, J.T.; Wandji, J.; Fomum, Z.T.; Chang, H.K.; Kim, Y.H. Prenylated Isoflavonoids from Erythrina Senegalensis. Phytochemistry 1999, 51, 1147–1150. [Google Scholar] [CrossRef]
- Taylor, R.B.; Corley, D.G.; Tempesta, M.S.; Fomum, Z.T.; Ayafor, J.F.; Wandji, J.; Ifeadike, P.N. 2,3-Dihydroauriculatin, a New Prenylated Isoflavanone from Erythrina Senegalensis. Application of the Selective Inept Technique. J. Nat. Prod. 1986, 49, 670–673. [Google Scholar] [CrossRef]
- Kuete, V.; Mbaveng, A.T.; Nono, E.C.N.; Simo, C.C.; Zeino, M.; Nkengfack, A.E.; Efferth, T. Cytotoxicity of Seven Naturally Occurring Phenolic Compounds towards Multi-Factorial Drug-Resistant Cancer Cells. Phytomed. Int. J. Phytother. Phytopharm. 2016, 23, 856–863. [Google Scholar] [CrossRef]
- Jaramillo-Rangel, G.; Chávez-Briones, M.-L.; Niderhauser-García, A.; Ortega-Martínez, M. Toxicity and Anticancer Potential of Karwinskia: A Review. Molecules 2020, 25, 5590. [Google Scholar] [CrossRef] [PubMed]
- Semwal, R.B.; Semwal, D.K.; Combrinck, S.; Viljoen, A. Emodin—A Natural Anthraquinone Derivative with Diverse Pharmacological Activities. Phytochemistry 2021, 190, 112854. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Chen, L.-J.; Huang, T.; Ying, J.-Q.; Li, J. The Pharmacology, Toxicology and Therapeutic Potential of Anthraquinone Derivative Emodin. Chin. J. Nat. Med. 2020, 18, 425–435. [Google Scholar] [CrossRef]
- Gnoula, C.; Mégalizzi, V.; De Nève, N.; Sauvage, S.; Ribaucour, F.; Guissou, P.; Duez, P.; Dubois, J.; Ingrassia, L.; Lefranc, F.; et al. Balanitin-6 and -7: Diosgenyl Saponins Isolated from Balanites Aegyptiaca Del. Display Significant Anti-Tumor Activity In Vitro and In Vivo. Int. J. Oncol. 2008, 32, 5–15. [Google Scholar] [CrossRef]
- Sobolewska, D.; Michalska, K.; Podolak, I.; Grabowska, K. Steroidal Saponins from the Genus Allium. Phytochem. Rev. Proc. Phytochem. Soc. Eur. 2016, 15, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, K.; Podolak, I.; Galanty, A.; Żmudzki, P.; Koczurkiewicz, P.; Piska, K.; Pękala, E.; Janeczko, Z. Two New Triterpenoid Saponins from the Leaves of Impatiens Parviflora DC. and Their Cytotoxic Activity. Ind. Crops Prod. 2017, 96, 71–79. [Google Scholar] [CrossRef]
- Moilanen, J.; Karonen, M.; Tähtinen, P.; Jacquet, R.; Quideau, S.; Salminen, J.-P. Biological Activity of Ellagitannins: Effects as Anti-Oxidants, pro-Oxidants and Metal Chelators. Phytochemistry 2016, 125, 65–72. [Google Scholar] [CrossRef]
- Yang, L.; Xian, D.; Xiong, X.; Lai, R.; Song, J.; Zhong, J. Proanthocyanidins against Oxidative Stress: From Molecular Mechanisms to Clinical Applications. BioMed Res. Int. 2018, 2018, 8584136. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Han, J.; Xiao, H.; Qiao, J.; Han, M. Effect of Tea Polyphenol Compounds on Anticancer Drugs in Terms of Anti-Tumor Activity, Toxicology, and Pharmacokinetics. Nutrients 2016, 8, 762. [Google Scholar] [CrossRef]
- Bouhenna, M.M.; Mameri, N.; Pérez, M.V.; Talhi, O.; Bachari, K.; Silva, A.M.S.; Luyten, W. Anticancer Activity Study of Chromone and Coumarin Hybrids Using Electrical Impedance Spectroscopy. Anticancer Agents Med. Chem. 2018, 18, 854–864. [Google Scholar] [CrossRef]
- Caro, A.A.; Davis, A.; Fobare, S.; Horan, N.; Ryan, C.; Schwab, C. Antioxidant and Pro-Oxidant Mechanisms of (+) Catechin in Microsomal CYP2E1-Dependent Oxidative Stress. Toxicol. In Vitro 2019, 54, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahebkar, A. Dual Effect of Curcumin in Preventing Atherosclerosis: The Potential Role of pro-Oxidant-Antioxidant Mechanisms. Nat. Prod. Res. 2015, 29, 491–492. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Levine, B. Autosis and Autophagic Cell Death: The Dark Side of Autophagy. Cell Death Differ. 2015, 22, 367–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bury, M.; Girault, A.; Mégalizzi, V.; Spiegl-Kreinecker, S.; Mathieu, V.; Berger, W.; Evidente, A.; Kornienko, A.; Gailly, P.; Vandier, C.; et al. Ophiobolin A Induces Paraptosis-like Cell Death in Human Glioblastoma Cells by Decreasing BKCa Channel Activity. Cell Death Dis. 2013, 4, e561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, L.; Liu, H.; Liu, H.; Bian, Z.; Zhang, Q.; Liao, W.; Sun, S. Preparation of warangalone-loaded liposomes and its inhibitory effect on breast cancer cells. J. South. Med. Univ. 2020, 40, 876–883. [Google Scholar] [CrossRef]
- Tchokouaha, R.F.; Alexi, X.; Chosson, E.; Besson, T.; Skaltsounis, A.-L.; Seguin, E.; Alexis, M.N.; Wandji, J. Erymildbraedin A and B, Two Novel Cytotoxic Dimethylpyrano-Isoflavones from the Stem Bark of Erythrina Mildbraedii: Evaluation of Their Activity toward Endocrine Cancer Cells. J. Enzyme Inhib. Med. Chem. 2010, 25, 228–233. [Google Scholar] [CrossRef]
- Nkengfack, A.E.; Azebaze, A.G.; Waffo, A.K.; Fomum, Z.T.; Meyer, M.; van Heerden, F.R. Cytotoxic Isoflavones from Erythrina Indica. Phytochemistry 2001, 58, 1113–1120. [Google Scholar] [CrossRef]
- Zhang, B.; Fan, X.; Wang, Z.; Zhu, W.; Li, J. Alpinumisoflavone Radiosensitizes Esophageal Squamous Cell Carcinoma through Inducing Apoptosis and Cell Cycle Arrest. Biomed. Pharmacother. 2017, 95, 199–206. [Google Scholar] [CrossRef]
- Desta, Z.Y.; Sewald, N.; Majinda, R.R. Cytotoxic Flavonoids from Erythrina Caffra Thunb. Bull. Chem. Soc. Ethiop. 2016, 30, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Byeon, S.E.; Yi, Y.-S.; Lee, J.; Yang, W.S.; Kim, J.H.; Kim, J.; Hong, S.; Kim, J.-H.; Cho, J.Y. Hydroquinone Exhibits In Vitro and In Vivo Anti-Cancer Activity in Cancer Cells and Mice. Int. J. Mol. Sci. 2018, 19, 903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunassee, S.; Davies-Coleman, M. Cytotoxic and Antioxidant Marine Prenylated Quinones and Hydroquinones. Nat. Prod. Rep. 2012, 29, 513–535. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.-C.; Chang, B.-E.; Pan, Y.-H.; Lin, B.-R.; Lian, Y.-C.; Lee, M.-S.; Yeung, S.-Y.; Lin, L.-D.; Jeng, J.-H. Antiplatelet, Antioxidative, and Anti-Inflammatory Effects of Hydroquinone. J. Cell. Physiol. 2019, 234, 18123–18130. [Google Scholar] [CrossRef] [PubMed]
- Sofi, M.S.; Sateesh, M.K.; Bashir, M.; Ganie, M.A.; Nabi, S. Chemopreventive and Anti-Breast Cancer Activity of Compounds Isolated from Leaves of Abrus Precatorius L. 3 Biotech 2018, 8, 371. [Google Scholar] [CrossRef]
- Bae, H.; Song, G.; Lim, W. Stigmasterol Causes Ovarian Cancer Cell Apoptosis by Inducing Endoplasmic Reticulum and Mitochondrial Dysfunction. Pharmaceutics 2020, 12, 488. [Google Scholar] [CrossRef]
- Liao, H.; Zhu, D.; Bai, M.; Chen, H.; Yan, S.; Yu, J.; Zhu, H.; Zheng, W.; Fan, G. Stigmasterol Sensitizes Endometrial Cancer Cells to Chemotherapy by Repressing Nrf2 Signal Pathway. Cancer Cell Int. 2020, 20, 480. [Google Scholar] [CrossRef]
- Sheu, J.H.; Wang, G.H.; Sung, P.J.; Duh, C.Y. New Cytotoxic Oxygenated Fucosterols from the Brown Alga Turbinaria Conoides. J. Nat. Prod. 1999, 62, 224–227. [Google Scholar] [CrossRef]
- Sheu, J.H.; Wang, G.H.; Sung, P.J.; Chiu, Y.H.; Duh, C.Y. Cytotoxic Sterols from the Formosan Brown Alga Turbinaria Ornata. Planta Med. 1997, 63, 571–572. [Google Scholar] [CrossRef]
- Pacheco, B.S.; Dos Santos, M.A.Z.; Schultze, E.; Martins, R.M.; Lund, R.G.; Seixas, F.K.; Colepicolo, P.; Collares, T.; Paula, F.R.; De Pereira, C.M.P. Cytotoxic Activity of Fatty Acids from Antarctic Macroalgae on the Growth of Human Breast Cancer Cells. Front. Bioeng. Biotechnol. 2018, 6, 185. [Google Scholar] [CrossRef]
- Jiang, H.; Li, J.; Chen, A.; Li, Y.; Xia, M.; Guo, P.; Yao, S.; Chen, S. Fucosterol Exhibits Selective Antitumor Anticancer Activity against HeLa Human Cervical Cell Line by Inducing Mitochondrial Mediated Apoptosis, Cell Cycle Migration Inhibition and Downregulation of m-TOR/PI3K/Akt Signalling Pathway. Oncol. Lett. 2018, 15, 3458–3463. [Google Scholar] [CrossRef]
- Shen, T.; Zhang, L.; Wang, Y.-Y.; Fan, P.-H.; Wang, X.-N.; Lin, Z.-M.; Lou, H.-X. Steroids from Commiphora Mukul Display Antiproliferative Effect against Human Prostate Cancer PC3 Cells via Induction of Apoptosis. Bioorg. Med. Chem. Lett. 2012, 22, 4801–4806. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.-T.; Wang, T.; Liu, A.-H.; Li, J.; Yao, L.-G.; Wang, B.; Guo, Y.-W.; Mao, S.-C. PTP1B Inhibitory and Cytotoxic C-24 Epimers of Δ28-24-Hydroxy Stigmastane-Type Steroids from the Brown Alga Dictyopteris Undulata Holmes. Phytochemistry 2018, 146, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Bonel-Pérez, G.C.; Pérez-Jiménez, A.; Gris-Cárdenas, I.; Parra-Pérez, A.M.; Lupiáñez, J.A.; Reyes-Zurita, F.J.; Siles, E.; Csuk, R.; Peragón, J.; Rufino-Palomares, E.E. Antiproliferative and Pro-Apoptotic Effect of Uvaol in Human Hepatocarcinoma HepG2 Cells by Affecting G0/G1 Cell Cycle Arrest, ROS Production and AKT/PI3K Signaling Pathway. Molecules 2020, 25, 4254. [Google Scholar] [CrossRef] [PubMed]
- Bano, Z.; Begum, S.; Ali, S.S.; Kiran, Z.; Siddiqui, B.S.; Ahmed, A.; Khawaja, S.; Fatima, F.; Jabeen, A. Phytochemicals from Carissa Carandas with Potent Cytotoxic and Anti-Inflammatory Activities. Nat. Prod. Res. 2021, 36, 1587–1592. [Google Scholar] [CrossRef] [PubMed]
- Sperandio, S.; de Belle, I.; Bredesen, D.E. An Alternative, Nonapoptotic Form of Programmed Cell Death. Proc. Natl. Acad. Sci. USA 2000, 97, 14376–14381. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M.J.; Lee, A.R.; Jeong, S.A.; Kim, Y.-S.; Kim, J.Y.; Kwon, Y.-J.; Choi, K.S. Release of Ca2+ from the Endoplasmic Reticulum and Its Subsequent Influx into Mitochondria Trigger Celastrol-Induced Paraptosis in Cancer Cells. Oncotarget 2014, 5, 6816–6831. [Google Scholar] [CrossRef] [Green Version]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as Anticancer Mechanism: Function and Dysfunction of Its Modulators and Targeted Therapeutic Strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wen, X.; Zhang, N.; Wang, L.; Hao, D.; Jiang, X.; He, G. Small-Molecule Compounds Target Paraptosis to Improve Cancer Therapy. Biomed. Pharmacother. 2019, 118, 109203. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Boyd-Kimball, D. Oxidative Stress, Amyloid-β Peptide, and Altered Key Molecular Pathways in the Pathogenesis and Progression of Alzheimer’s Disease. J. Alzheimers Dis. 2018, 62, 1345–1367. [Google Scholar] [CrossRef] [Green Version]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Ahotupa, M. Oxidized Lipoprotein Lipids and Atherosclerosis. Free Radic. Res. 2017, 51, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubczyk, K.; Dec, K.; Kałduńska, J.; Kawczuga, D.; Kochman, J.; Janda, K. Reactive Oxygen Species—Sources, Functions, Oxidative Damage. Pol. Merkur. Lek. Organ Pol. Tow. Lek. 2020, 48, 124–127. [Google Scholar]
- Ciulei, I. Methodology for Analysis of Vegetable Drugs. Practical Manual on the Industrial Utilisation of Medicinal and Aromatic Plants.; Ministry of Chemical Industry: Bucarest, Romania, 1982; p. 62. [Google Scholar]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the Total Phenolic, Flavonoid and Proline Contents in Burkina Fasan Honey, as Well as Their Radical Scavenging Activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Arvouet-Grand, A.; Vennat, B.; Pourrat, A.; Legret, P. Standardization of propolis extract and identification of principal constituents. J. Pharm. Belg. 1994, 49, 462–468. [Google Scholar]
- Xiao, F.; Xu, T.; Lu, B.; Liu, R. Guidelines for Antioxidant Assays for Food Components. Food Front. 2020, 1, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Nenadis, N.; Wang, L.-F.; Tsimidou, M.; Zhang, H.-Y. Estimation of Scavenging Activity of Phenolic Compounds Using the ABTS(*+) Assay. J. Agric. Food Chem. 2004, 52, 4669–4674. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Bunea, A.; Rugină, D.; Sconţa, Z.; Pop, R.M.; Pintea, A.; Socaciu, C.; Tăbăran, F.; Grootaert, C.; Struijs, K.; VanCamp, J. Anthocyanin Determination in Blueberry Extracts from Various Cultivars and Their Antiproliferative and Apoptotic Properties in B16-F10 Metastatic Murine Melanoma Cells. Phytochemistry 2013, 95, 436–444. [Google Scholar] [CrossRef]
- Colin, M.; Delporte, C.; Janky, R.; Lechon, A.-S.; Renard, G.; Van Antwerpen, P.; Maltese, W.A.; Mathieu, V. Dysregulation of Macropinocytosis Processes in Glioblastomas May Be Exploited to Increase Intracellular Anti-Cancer Drug Levels: The Example of Temozolomide. Cancers 2019, 11, 411. [Google Scholar] [CrossRef] [Green Version]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS Using Oxidized DCFDA and Flow-Cytometry. In Advanced Protocols in Oxidative Stress II; Humana Press: Totowa, NJ, USA, 2010; Volume 594, pp. 57–72. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemical Groups | Extracts | ||
|---|---|---|---|
| MeOH | CH2Cl2 | MeOH/CH2Cl2 | |
| Alkaloids | − | − | − |
| Anthocyanosides | − | − | − |
| Anthracenosides | + | − | + |
| Cardiotonic glycosides | − | − | − |
| Coumarin derivatives | + | − | + |
| Coumarins | − | + | + |
| Emodols | + | − | + |
| Flavonic aglycones | − | + | + |
| Flavonoids | + | + | + |
| Saponosides | + | NT | + |
| Steroidal and triterpene glucosides | + | + | + |
| Sterols and triterpenes | − | + | + |
| Tannins | + | NT | + |
| Extracts/Fractions | 50% Inhibitory Concentration on Cell Lines | ||||
|---|---|---|---|---|---|
| (Mean ± SEM) µg/mL | |||||
| U373 | MCF-7 | A549 | SKMEL-28 | B16F10 | |
| MeOH extract | 66 ± 3 | 38 ± 2 | 42 ± 1 | 36 ± 1 | 33 ± 2 |
| CH2Cl2/MeOH extract | 77 ± 2 | 41 ± 1 | 48 ± 1 | 36 ± 1 | 33 ± 1 |
| CH2Cl2 extract | 37 ± 2 | 29 ± 2 | 19 ± 3 | 29 ± 1 | 37 ± 1 |
| EtOAc subfraction | 34 ± 1 | 25 ± 2 | 19 ± 2 | 27 ± 2 | 32 ± 1 |
| Extracts | Antioxidant Capacity | Total Phenolic | Total Flavonoids | ||
|---|---|---|---|---|---|
| DPPH | ABTS | FRAP | |||
| MeOH | 0.56 ± 0.01 | 0.76 ± 0.02 | 1.34 ± 0.08 | 132.54 ± 0.02 | 101.23 ± 0.08 |
| CH2Cl2/MeOH | 0.67 ± 0.01 | 1.06 ± 0.03 | 1.01 ± 0.04 | 82.01 ± 1.01 | 47.11 ± 0.52 |
| CH2Cl2 | 6.67 ± 0.04 | 7.92 ± 0.20 | 12.07 ± 0.04 | ND | ND |
| EtOAc | 5.02 ± 0.01 | 8.23 ± 0.12 | 10.98 ± 0.05 | 23 ± 2 | 19 ± 3 |
| Chemical Group | Substances | Extract Source | Chemical Data | |||||
|---|---|---|---|---|---|---|---|---|
| MeOH | EtOAc | RT (min) | Formula | Error (ppm) | Measured m/z | Molecular Species | ||
| Triterpene | Uvaol | + | 16.149 | C30H50O2 | 1.26 | 443.3878 | [M + H]+ | |
| Steroids | 16:3-Glc-Stigmasterol | + | 13.568 | C51H82O7 | 3.87 | 426.2939 | [M + 2Na]2+ | |
| Steroids | 22-Hydroxy-campesterol | + | 16.911 | C28H48O2 | 3.99 | 439.3529 | [M + Na]+ | |
| Steroids | 7-Campestenol | + | + | 15.344 | C28H48O | 2.04 | 423.3606 | [M+ Na]+ |
| Steroids | Ergostanol | + | 16.141 | C28H50O | 4.5 | 425.3773 | [M + Na]+ | |
| Steroids | Feruloyldihydro-β-sitosterol | + | 17.465 | C39H60O4 | 2.41 | 615.4369 | [M + Na]+ | |
| Steroidal glucoside | Isofucosterol 3-O- 6-O- [Hexadecanoyl-b-D-glucopyranoside] | + | + | 12.896 | C51H88O7 | 2.16 | 445.2887 | [M + 2K]2+ |
| Fatty acids | 9,12-Octadecadienoic acid (Z,Z) | + | + | 15.057 | C18H32O2 | 0.19 | 298.274 | [M + NH4]+ |
| Fatty acids | α-linolinic acid | + | + | 14.833 | C18H30O2 | 1.64 | 279.2314 | [M + H]+ |
| Fatty acids | Hexadecanoic acid | + | 16.223 | C16H32O2 | 0.03 | 257.2475 | [M + H]+ | |
| Diphenol | 1,4-Benzenediol (hydroquinone) | + | 5.477 | C6H6O2 | 1.6 | 128.0704 | [M + NH4]+ | |
| Chalcone | 2′,4′,6′-Trihydroxy chalcone | + | 15.848 | C15H12O4 | 0.25 | 257.0809 | [M + H]+ | |
| Chemical Group | Substances | Extract Source | Chemical Data | |||||
|---|---|---|---|---|---|---|---|---|
| MeOH | EtOAc | RT (min) | Formula | Error (ppm) | Measured m/z | Molecular Species | ||
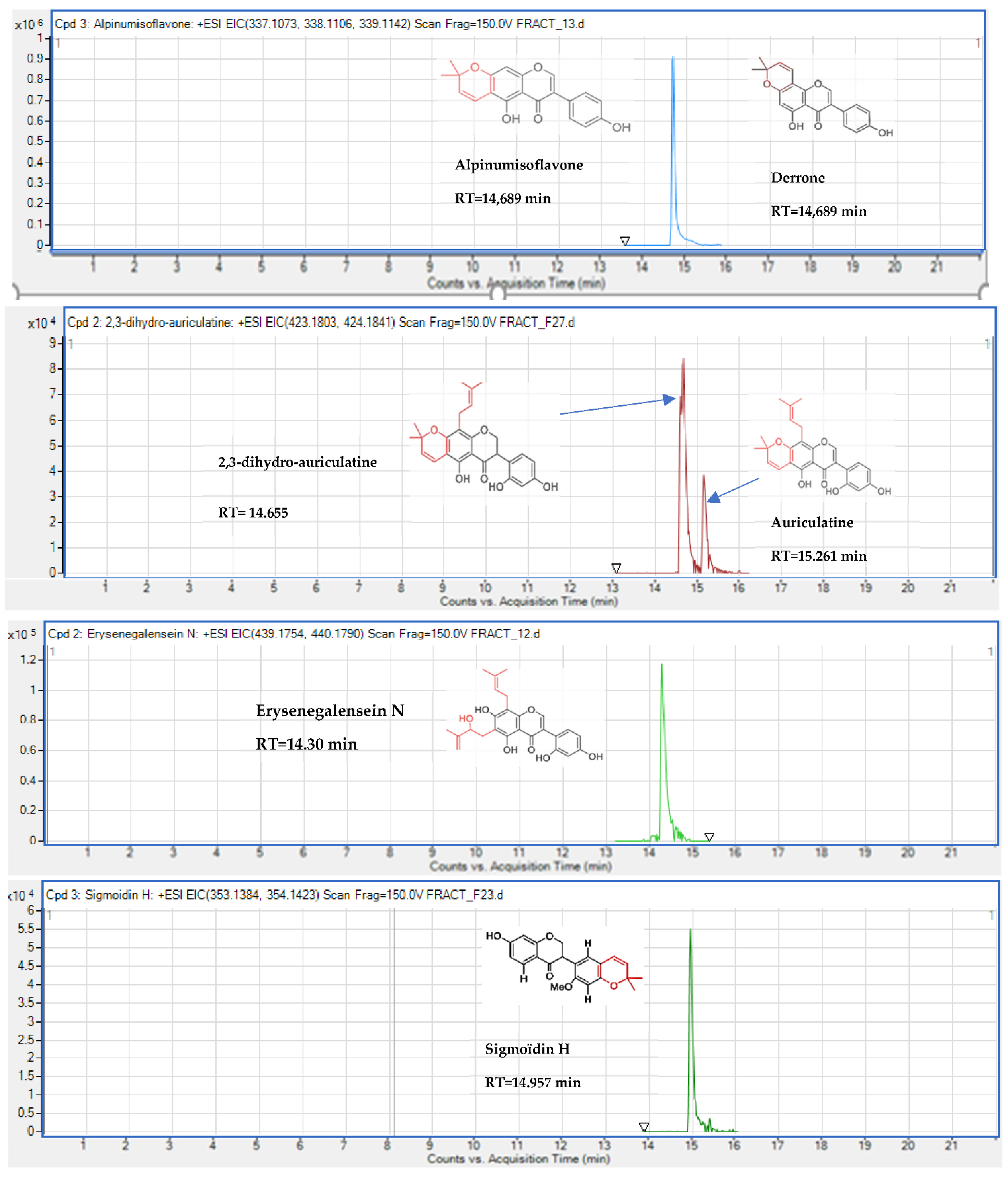
| Isoflavonoids | 2,3-dihydro-auriculatine | + | + | 14.615 | C25H26O6 | 0.91 | 423.1806 | [M + H]+ |
| Isoflavonoids | Alpinumisoflavone or derrone | + | + | 14.689 | C20H16O5 | 1.63 | 337.1076 | [M + H]+ |
| Isoflavonoids | Auriculatin or auriculasin | + | + | 15.261 | C25H24O6 | 1.75 | 421.1653 | [M + H]+ |
| Isoflavonoids | Erysenegalensein N | + | 14.64 | C25H26O7 | 1.75 | 439.1759 | [M + H]+ | |
| Isoflavonoids | Sigmoidin H | + | + | 14.96 | C21H20O5 | 0.42 | 353.1385 | [M + H]+ |
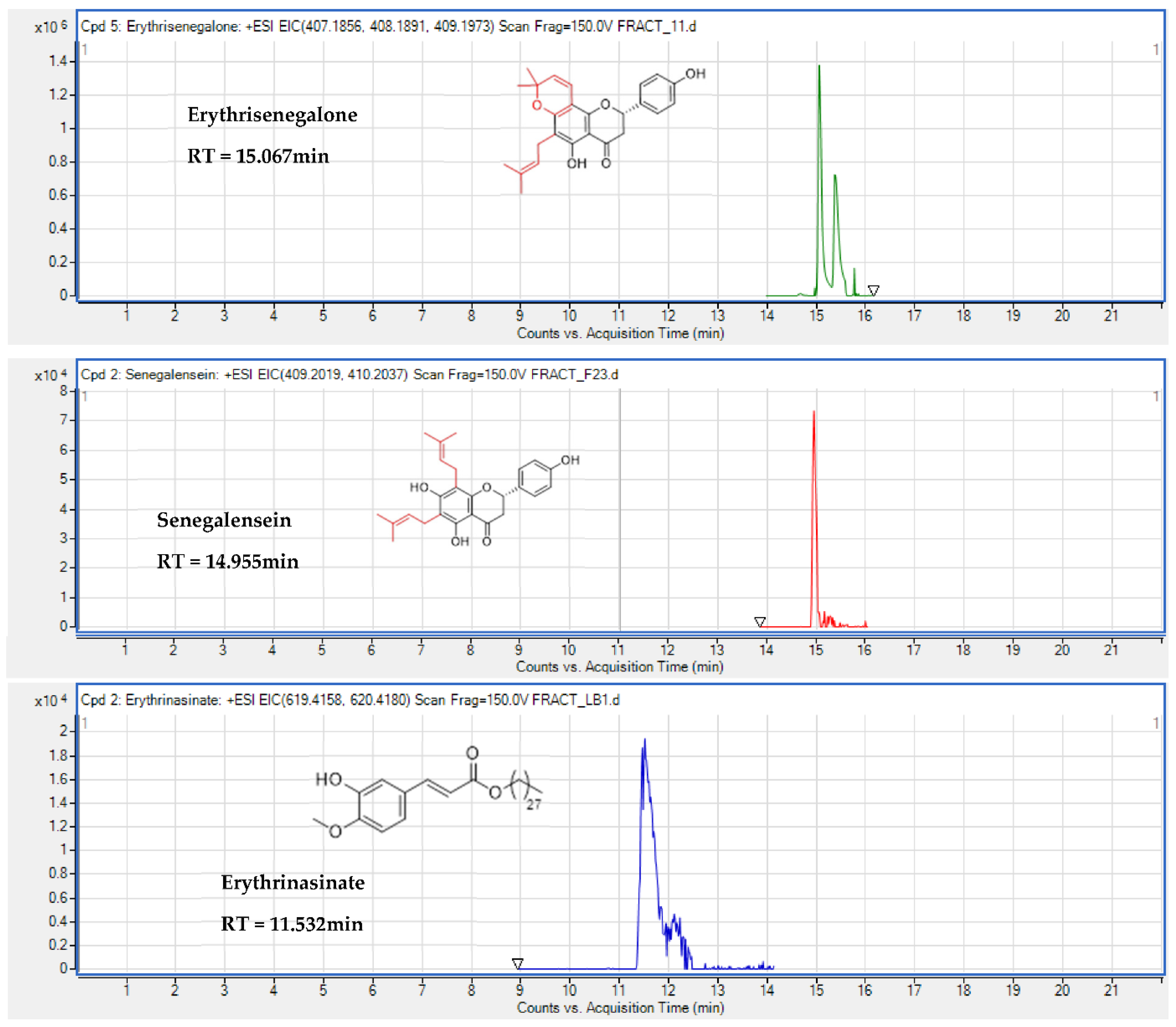
| Flavonoids | Erythrisenegalone | + | + | 15.067 | C25H26O5 | 0.74 | 407.1856 | [M + H]+ |
| Flavanone | Senegalensein | + | 14.955 | C25H28O5 | 1.1 | 409.2014 | [M + H]+ | |
| Cinnamate | Erythrinasinate | + | 11.532 | C38H60O4 | 4.49 | 619.4151 | [M + K]+ | |
| N° | Putative Formula | RT (min) | Measured m/z | Error (ppm) | Molecular Species |
|---|---|---|---|---|---|
| 1 | C8H4O3 | 16.764 | 149.0231 | 1.48 | [M + H]+ |
| 2 | C8H6O4 | 16.757 | 167.0341 | 1.29 | [M + H]+ |
| 3 | C14H29NO | 15.246 | 228.2321 | 0.4 | [M + H]+ |
| 4 | C16H22O4 | 16.757 | 279.1592 | 0.41 | [M + H]+ |
| 5 | C16H31NO | 15.4 | 254.2478 | 0.16 | [M + H]+ |
| 6 | C16H33NO | 15.815 | 256.2634 | 0.36 | [M + H]+ |
| 7 | C16H34O7 | 12.67 | 339.238 | 0.8 | [M + H]+ |
| 8 | C18H32O | 15.945 | 282.2788 | 1.21 | [M + NH4]+ |
| 9 | C18H35NO | 16.217 | 282.2791 | 0.15 | [M + H]+ |
| 10 | C18H37NO | 16.343 | 284.2947 | 0.32 | [M + H]+ |
| 11 | C20H42O9 | 12.956 | 444.3164 | 0.69 | [M + NH4]+ |
| 12 | C22H46O10 | 13.037 | 488.343 | 0.16 | [M + NH4]+ |
| 13 | C23H48O10 | 13.25 | 502.3586 | 0.05 | [M + NH4]+ |
| 14 | C24H38O4 | 16.778 | 391.284 | 0.73 | [M + H]+ |
| 15 | C28H43N | 17.562 | 394.3464 | 1.08 | [M + H]+ |
| 16 | C28H46O4 | 17.581 | 447.3469 | 0.03 | [M + H]+ |
| 17 | C36H70N2O2 | 15.931 | 563.5498 | 2.14 | [M + H]+ |
| 18 | C40H69N3O9 | 19.395 | 736.5099 | 1.03 | [M + H]+ |
| 19 | C44H58N2O3 | 19.388 | 663.4528 | 1.18 | [M + H]+ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fofana, S.; Delporte, C.; Calvo Esposito, R.; Ouédraogo, M.; Van Antwerpen, P.; Guissou, I.P.; Semdé, R.; Mathieu, V. In Vitro Antioxidant and Anticancer Properties of Various E. senegalensis Extracts. Molecules 2022, 27, 2583. https://doi.org/10.3390/molecules27082583
Fofana S, Delporte C, Calvo Esposito R, Ouédraogo M, Van Antwerpen P, Guissou IP, Semdé R, Mathieu V. In Vitro Antioxidant and Anticancer Properties of Various E. senegalensis Extracts. Molecules. 2022; 27(8):2583. https://doi.org/10.3390/molecules27082583
Chicago/Turabian StyleFofana, Souleymane, Cédric Delporte, Rafaèle Calvo Esposito, Moussa Ouédraogo, Pierre Van Antwerpen, Innocent Pierre Guissou, Rasmané Semdé, and Véronique Mathieu. 2022. "In Vitro Antioxidant and Anticancer Properties of Various E. senegalensis Extracts" Molecules 27, no. 8: 2583. https://doi.org/10.3390/molecules27082583
APA StyleFofana, S., Delporte, C., Calvo Esposito, R., Ouédraogo, M., Van Antwerpen, P., Guissou, I. P., Semdé, R., & Mathieu, V. (2022). In Vitro Antioxidant and Anticancer Properties of Various E. senegalensis Extracts. Molecules, 27(8), 2583. https://doi.org/10.3390/molecules27082583