
Vanillin Induces Relaxation in Rat Mesenteric Resistance Arteries by Inhibiting Extracellular Ca2+ Influx


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
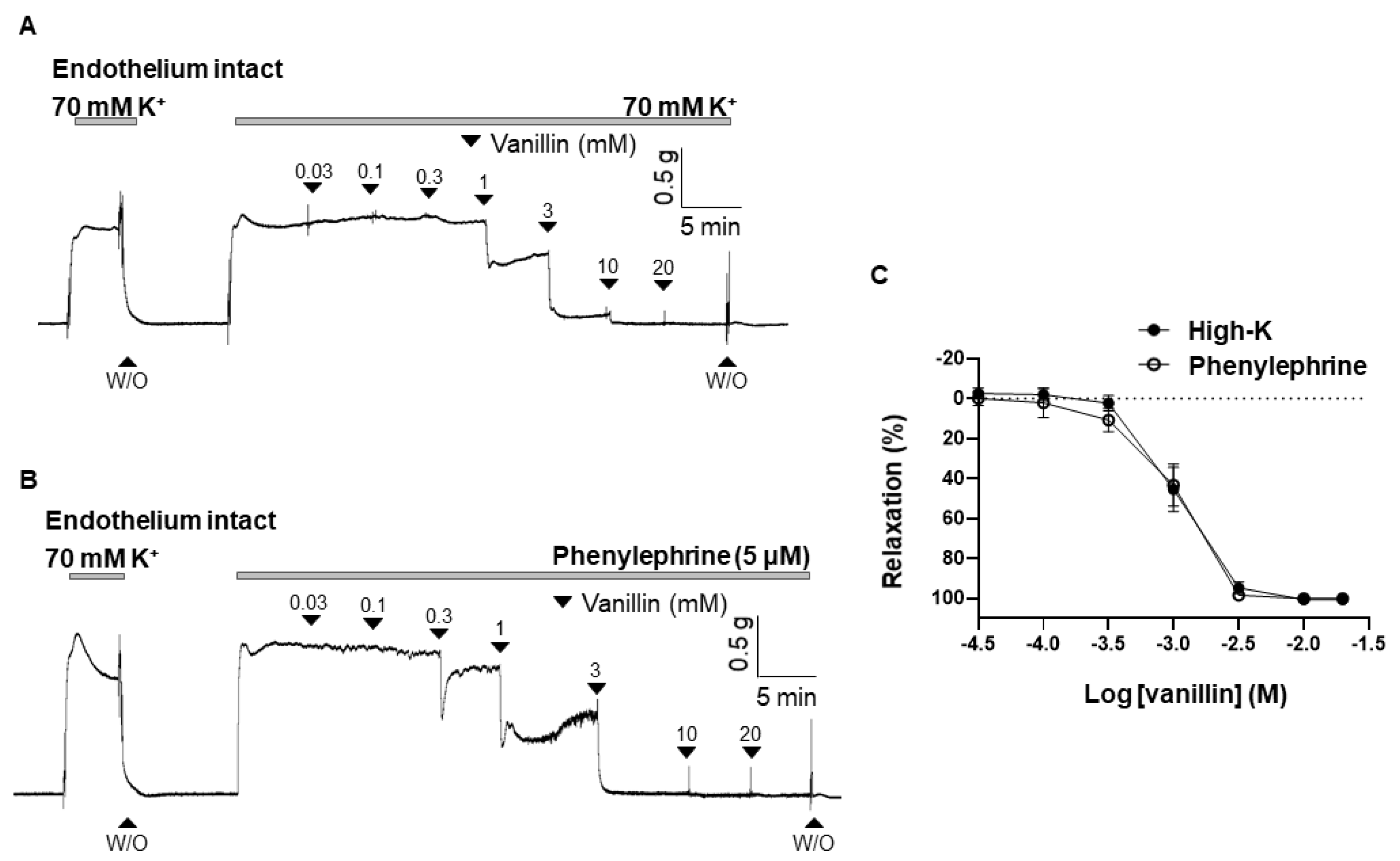
2.1. Effect of Vanillin on the Contraction Induced by High K+ or Phenylephrine
2.2. Endothelium-Independent Vasodilation Induced by Vanillin
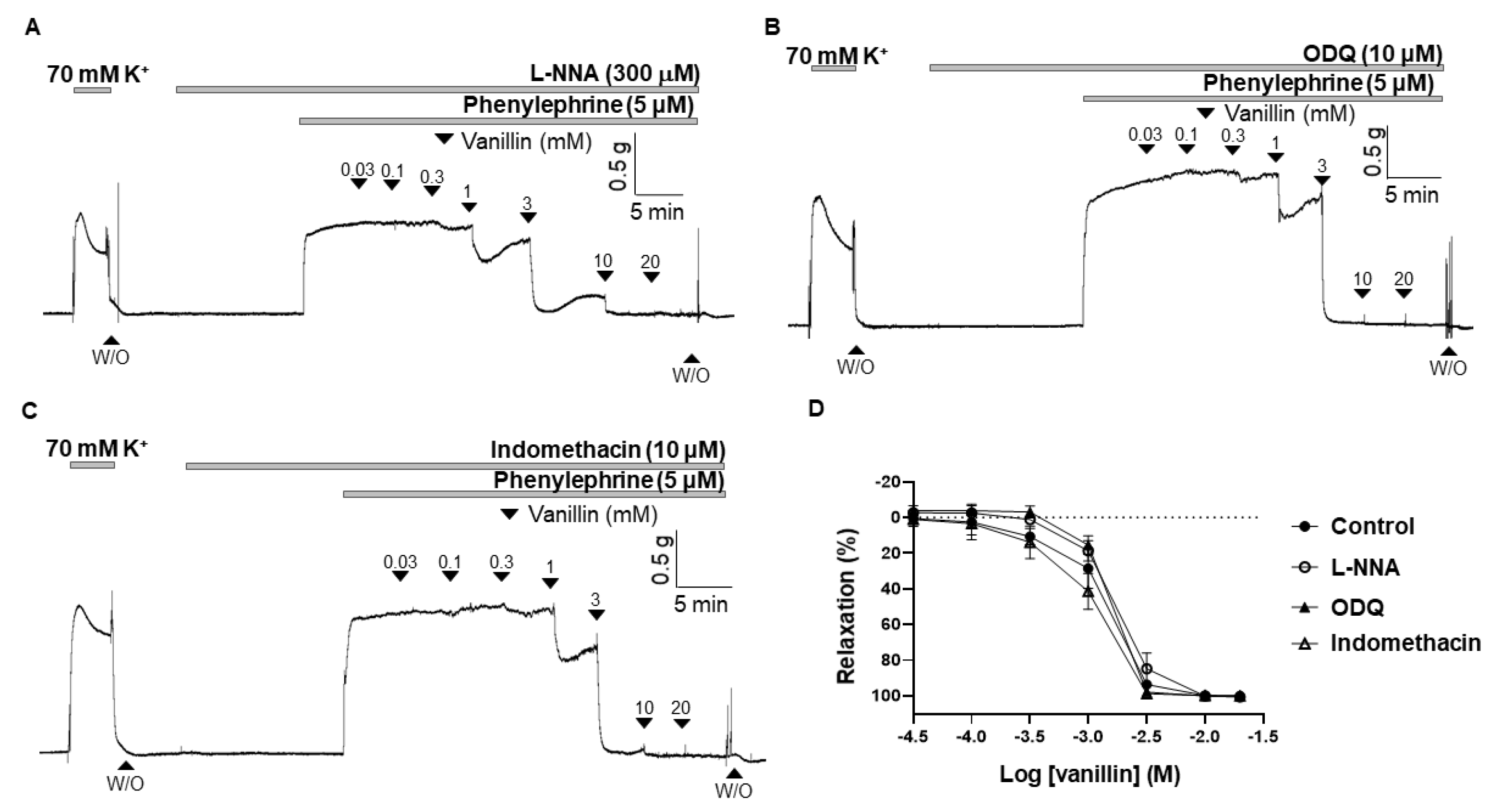
2.3. Effect of eNOS Inhibitor (L-NNA), sGC Inhibitor (ODQ), and COX Inhibitor (Indomethacin) on the Vasodilation Induced by Vanillin
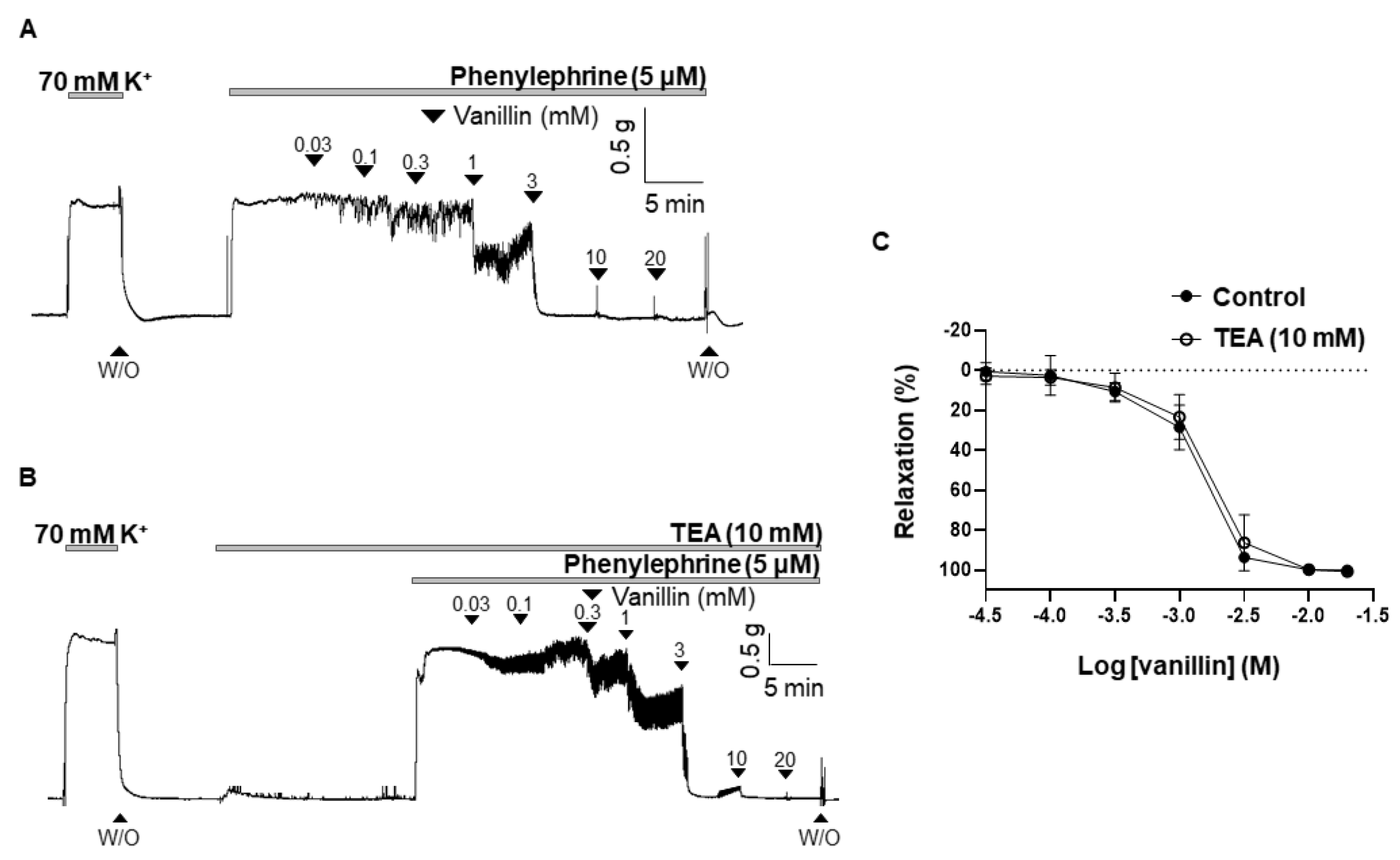
2.4. Effect of TEA on the Vasodilation Induced by Vanillin
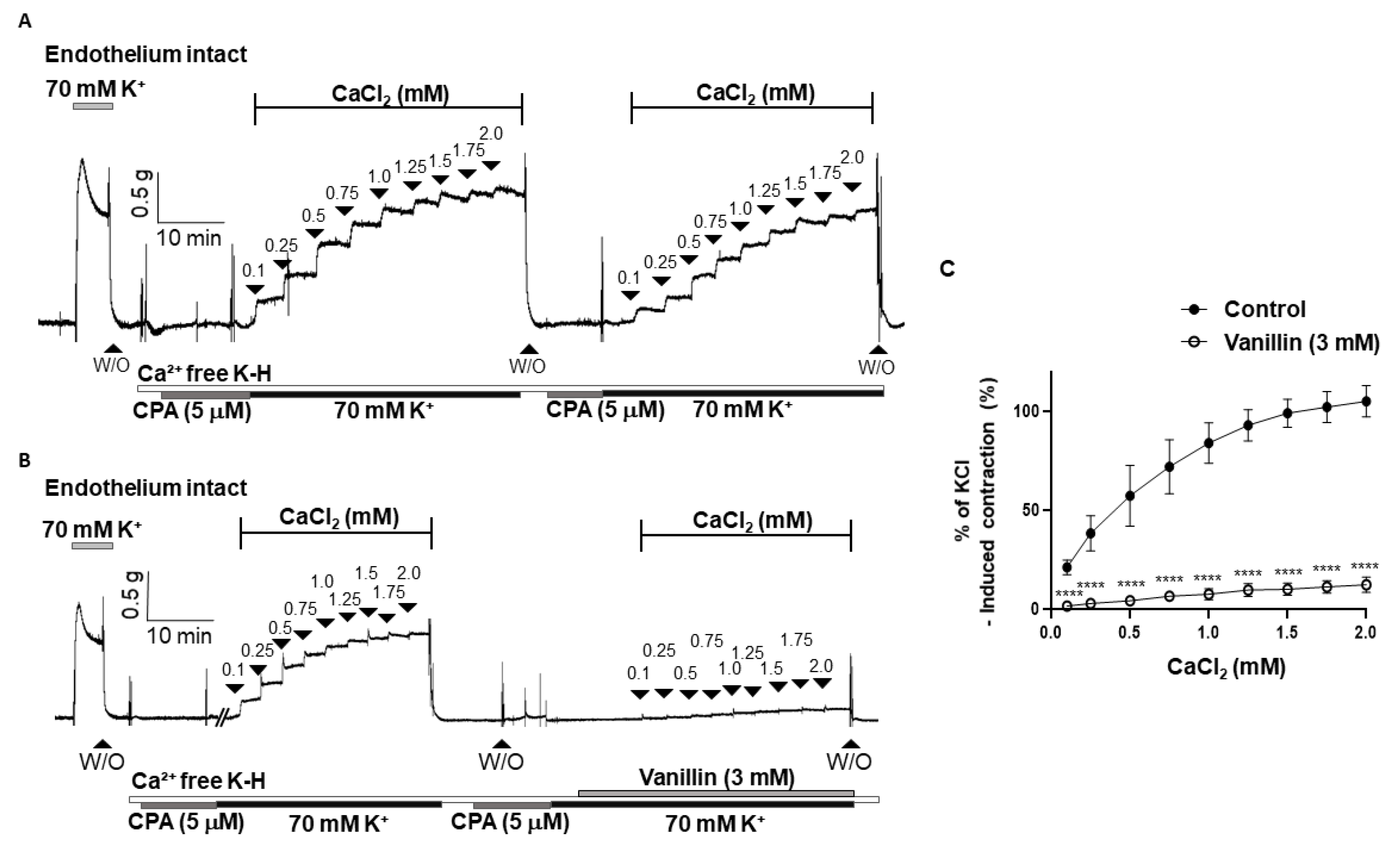
2.5. Effect of Vanillin on Extracellular Ca2+-Induced Vascular Contraction
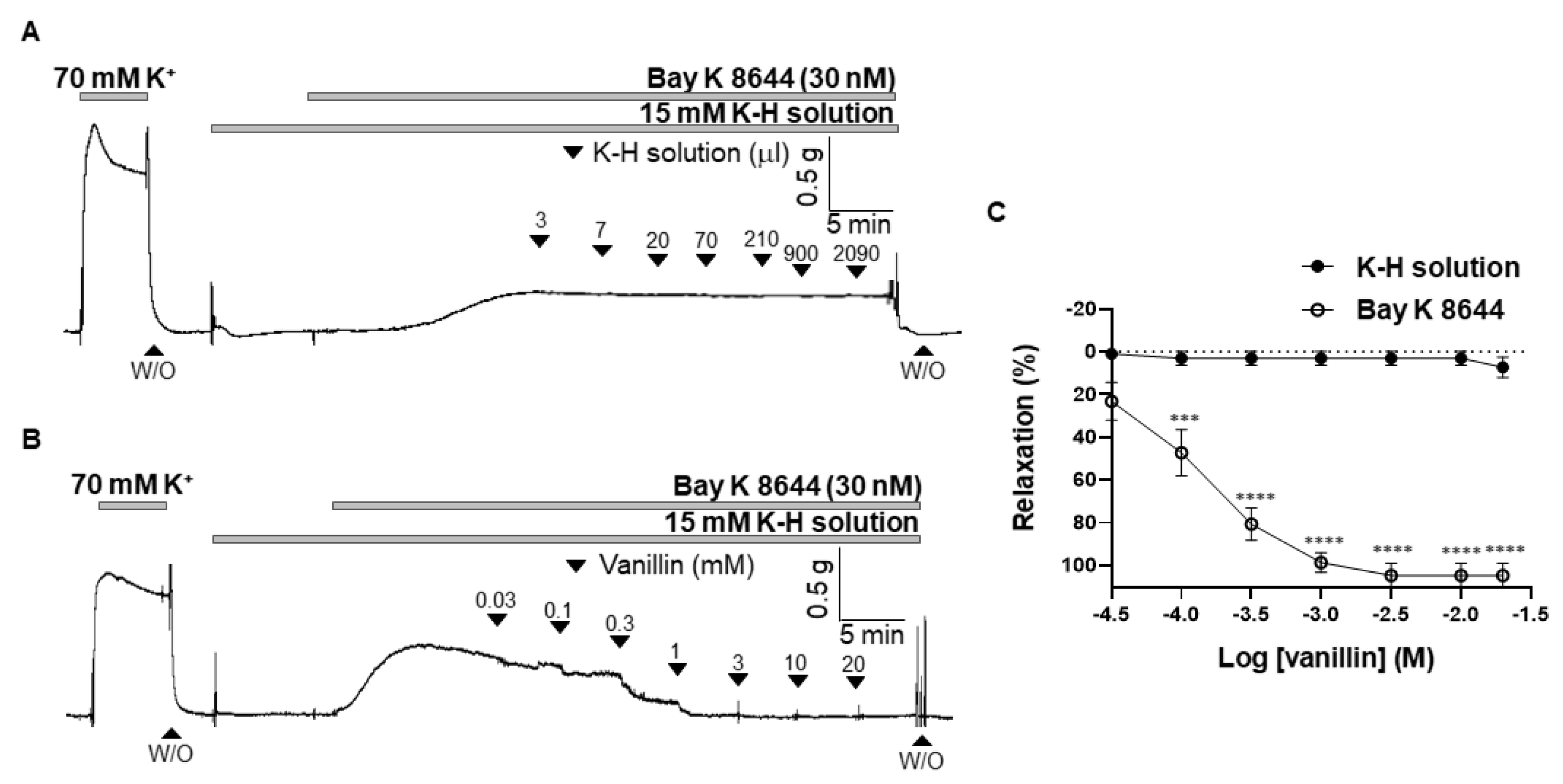
2.6. Effect of Vanillin on the BAY K 8644-Induced Contraction
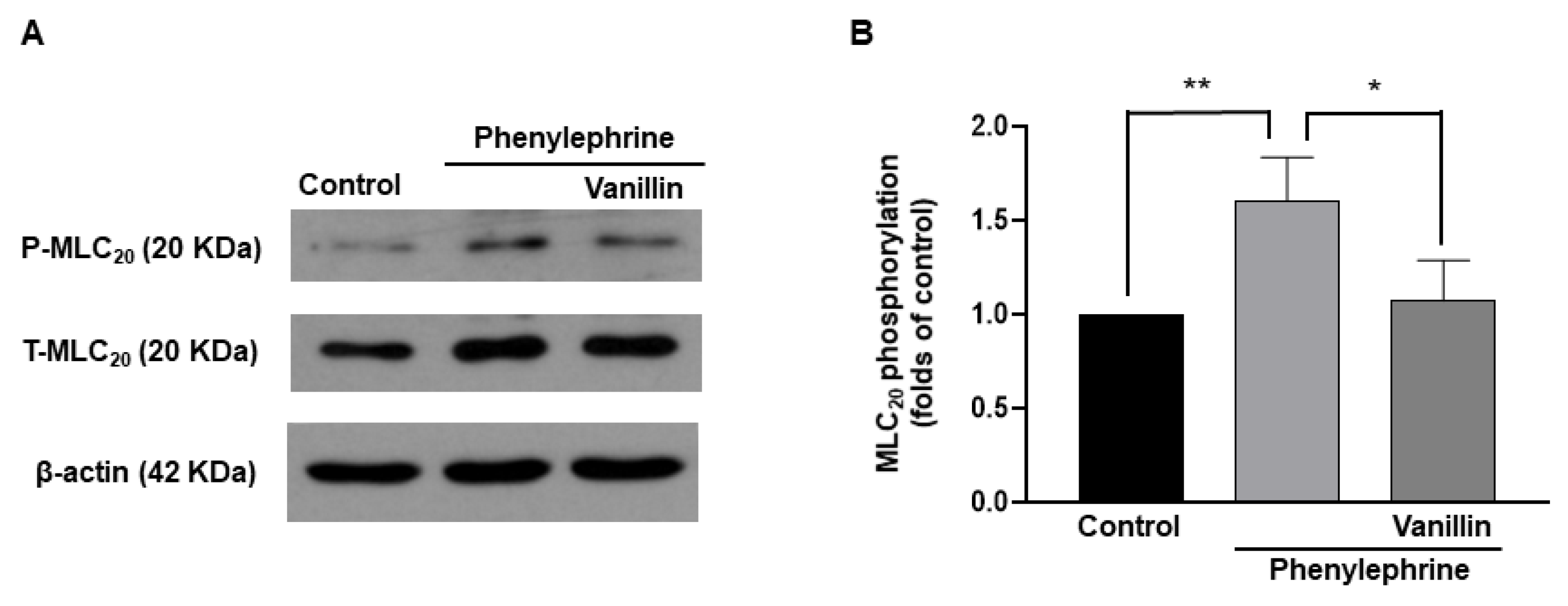
2.7. Decreased Phosphorylation of MLC20 by Vanillin in Vascular Smooth Muscle Cells
3. Discussion
4. Materials and Methods
4.1. Animals and Tissue Preparation
4.2. Measurement of Isometric Tension in Mesenteric Arteries
4.3. Isolation and Culture of Vascular Smooth Muscle Cells
4.4. Western Blot Analysis
4.5. Chemicals and Reagents
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Schiffrin, E.L. Reactivity of small blood vessels in hypertension: Relation with structural changes. State of the art lecture. Hypertension 1992, 19, II1-9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Intengan, H.D.; Schiffrin, E.L. Structure and mechanical properties of resistance arteries in hypertension: Role of adhesion molecules and extracellular matrix determinants. Hypertension 2000, 36, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Lund-Johansen, P. Haemodynamics in early essential hypertension--still an area of controversy. J. Hypertens. 1983, 1, 209–213. [Google Scholar] [PubMed]
- Edwards, J.C.; Ignarro, L.J.; Hyman, A.L.; Kadowitz, P.J. Relaxation of intrapulmonary artery and vein by nitrogen oxide-containing vasodilators and cyclic gmp. J. Pharmacol. Exp. Ther. 1984, 228, 33–42. [Google Scholar] [PubMed]
- Gallage, N.J.; Moller, B.L. Vanillin-bioconversion and bioengineering of the most popular plant flavor and its de novo biosynthesis in the vanilla orchid. Mol. Plant 2015, 8, 40–57. [Google Scholar] [CrossRef] [Green Version]
- Marton, A.; Kusz, E.; Kolozsi, C.; Tubak, V.; Zagotto, G.; Buzas, K.; Quintieri, L.; Vizler, C. Vanillin analogues o-vanillin and 2,4,6-trihydroxybenzaldehyde inhibit nfkb activation and suppress growth of a375 human melanoma. Anticancer. Res. 2016, 36, 5743–5750. [Google Scholar] [CrossRef]
- Gallage, N.J.; Hansen, E.H.; Kannangara, R.; Olsen, C.E.; Motawia, M.S.; Jorgensen, K.; Holme, I.; Hebelstrup, K.; Grisoni, M.; Moller, B.L. Vanillin formation from ferulic acid in vanilla planifolia is catalysed by a single enzyme. Nat. Commun. 2014, 5, 4037. [Google Scholar] [CrossRef] [Green Version]
- Tai, A.; Sawano, T.; Yazama, F.; Ito, H. Evaluation of antioxidant activity of vanillin by using multiple antioxidant assays. Biochim. Biophys. Acta Gen. Subj. 2011, 1810, 170–177. [Google Scholar] [CrossRef]
- Kwon, J.; Kim, J.; Park, S.; Khang, G.; Kang, P.M.; Lee, D. Inflammation-responsive antioxidant nanoparticles based on a polymeric prodrug of vanillin. Biomacromolecules 2013, 14, 1618–1626. [Google Scholar] [CrossRef]
- Lirdprapamongkol, K.; Kramb, J.P.; Suthiphongchai, T.; Surarit, R.; Srisomsap, C.; Dannhardt, G.; Svasti, J. Vanillin suppresses metastatic potential of human cancer cells through pi3k inhibition and decreases angiogenesis in vivo. J. Agric. Food Chem. 2009, 57, 3055–3063. [Google Scholar] [CrossRef]
- Salau, V.F.; Erukainure, O.L.; Olofinsan, K.O.; Msomi, N.Z.; Ijomone, O.M.; Islam, M.S. Vanillin improves glucose homeostasis and modulates metabolic activities linked to type 2 diabetes in fructose-streptozotocin induced diabetic rats. Arch. Physiol. Biochem. 2021, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, K.; Sawabe, A.; Sugimoto, T.; Koga, M.; Okuda, H.; Kitayama, T.; Shirai, M.; Komai, K.; Komemushi, S.; Matsuda, K. Inhibitory actions of several natural products on proliferation of rat vascular smooth muscle cells induced by hsp60 from chlamydia pneumoniae j138. J. Agric. Food Chem. 2004, 52, 6326–6329. [Google Scholar] [CrossRef]
- Raffai, G.; Khang, G.; Vanhoutte, P.M. Vanillin and vanillin analogs relax porcine coronary and basilar arteries by inhibiting l-type Ca2+ channels. J. Pharmacol. Exp. Ther. 2015, 352, 14–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haam, C.E.; Byeon, S.; Choi, S.; Oh, E.Y.; Choi, S.-K.; Lee, Y.-H. Vasorelaxant effect of Trachelospermi caulis extract on rat mesenteric resistance arteries. Molecules 2022, 27, 5300. [Google Scholar] [CrossRef]
- Webb, R.C. Smooth muscle contraction and relaxation. Adv. Physiol. Educ. 2003, 27, 201–206. [Google Scholar] [CrossRef]
- Pi, X.; Xie, L.; Patterson, C. Emerging roles of vascular endothelium in metabolic homeostasis. Circ. Res. 2018, 123, 477–494. [Google Scholar] [CrossRef] [PubMed]
- Sandoo, A.; van Zanten, J.J.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The endothelium and its role in regulating vascular tone. Open Cardiovasc. Med. J. 2010, 4, 302–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munzel, T.; Daiber, A.; Ullrich, V.; Mulsch, A. Vascular consequences of endothelial nitric oxide synthase uncoupling for the activity and expression of the soluble guanylyl cyclase and the cgmp-dependent protein kinase. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1551–1557. [Google Scholar] [CrossRef]
- Ricciotti, E.; Yu, Y.; Grosser, T.; Fitzgerald, G.A. Cox-2, the dominant source of prostacyclin. Proc. Natl. Acad. Sci. USA 2013, 110, E183. [Google Scholar] [CrossRef] [Green Version]
- Ahn, D.S.; Kim, Y.B.; Lee, Y.H.; Kang, B.S.; Kang, D.H. Fatty acids directly increase the activity of Ca(2+)-activated k+ channels in rabbit coronary smooth muscle cells. Yonsei Med. J. 1994, 35, 10–24. [Google Scholar] [CrossRef]
- Jackson, W.F. Potassium channels in regulation of vascular smooth muscle contraction and growth. Adv. Pharmacol. 2017, 78, 89–144. [Google Scholar] [PubMed] [Green Version]
- Heginbotham, L.; MacKinnon, R. The aromatic binding site for tetraethylammonium ion on potassium channels. Neuron 1992, 8, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Kamm, K.E.; Stull, J.T. The function of myosin and myosin light chain kinase phosphorylation in smooth muscle. Annu. Rev. Pharm. Toxicol. 1985, 25, 593–620. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.K.; Ahn, D.S.; Lee, Y.H. Comparison of contractile mechanisms of sphingosylphosphorylcholine and sphingosine-1- phosphate in rabbit coronary artery. Cardiovasc. Res. 2009, 82, 324–332. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.; Haam, C.E.; Oh, E.-Y.; Byeon, S.; Choi, S.-K.; Lee, Y.-H. Vanillin Induces Relaxation in Rat Mesenteric Resistance Arteries by Inhibiting Extracellular Ca2+ Influx. Molecules 2023, 28, 288. https://doi.org/10.3390/molecules28010288
Choi S, Haam CE, Oh E-Y, Byeon S, Choi S-K, Lee Y-H. Vanillin Induces Relaxation in Rat Mesenteric Resistance Arteries by Inhibiting Extracellular Ca2+ Influx. Molecules. 2023; 28(1):288. https://doi.org/10.3390/molecules28010288
Chicago/Turabian StyleChoi, Sooyeon, Chae Eun Haam, Eun-Yi Oh, Seonhee Byeon, Soo-Kyoung Choi, and Young-Ho Lee. 2023. "Vanillin Induces Relaxation in Rat Mesenteric Resistance Arteries by Inhibiting Extracellular Ca2+ Influx" Molecules 28, no. 1: 288. https://doi.org/10.3390/molecules28010288
APA StyleChoi, S., Haam, C. E., Oh, E. -Y., Byeon, S., Choi, S. -K., & Lee, Y. -H. (2023). Vanillin Induces Relaxation in Rat Mesenteric Resistance Arteries by Inhibiting Extracellular Ca2+ Influx. Molecules, 28(1), 288. https://doi.org/10.3390/molecules28010288