

Unravelling Novel Phytochemicals and Anticholinesterase Activity in Irish Cladonia portentosa



Abstract
:1. Introduction
2. Results
2.1. HPLC Analysis
2.2. LC-MS Analysis
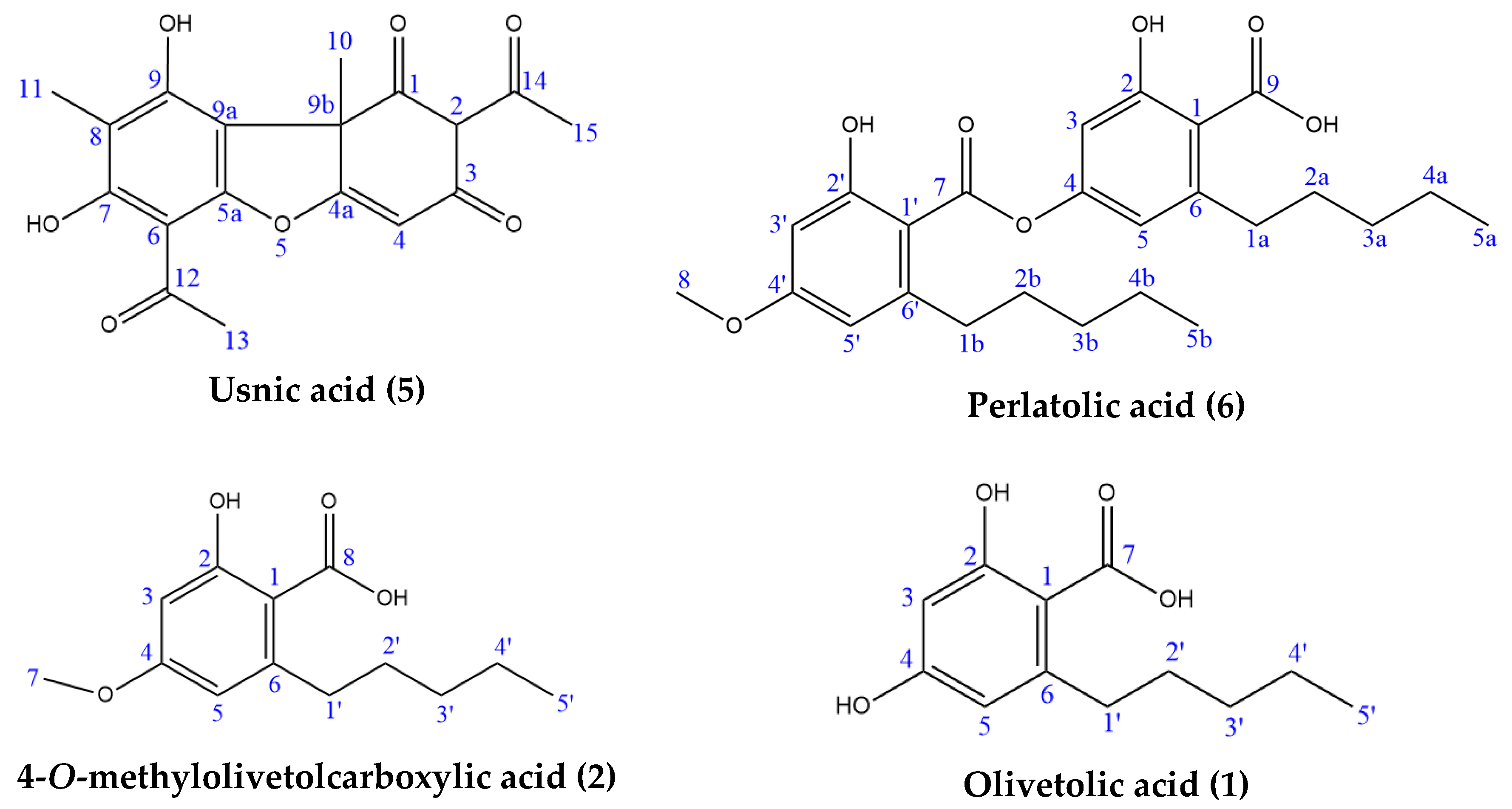
2.3. Isolation and Characterisation of Components
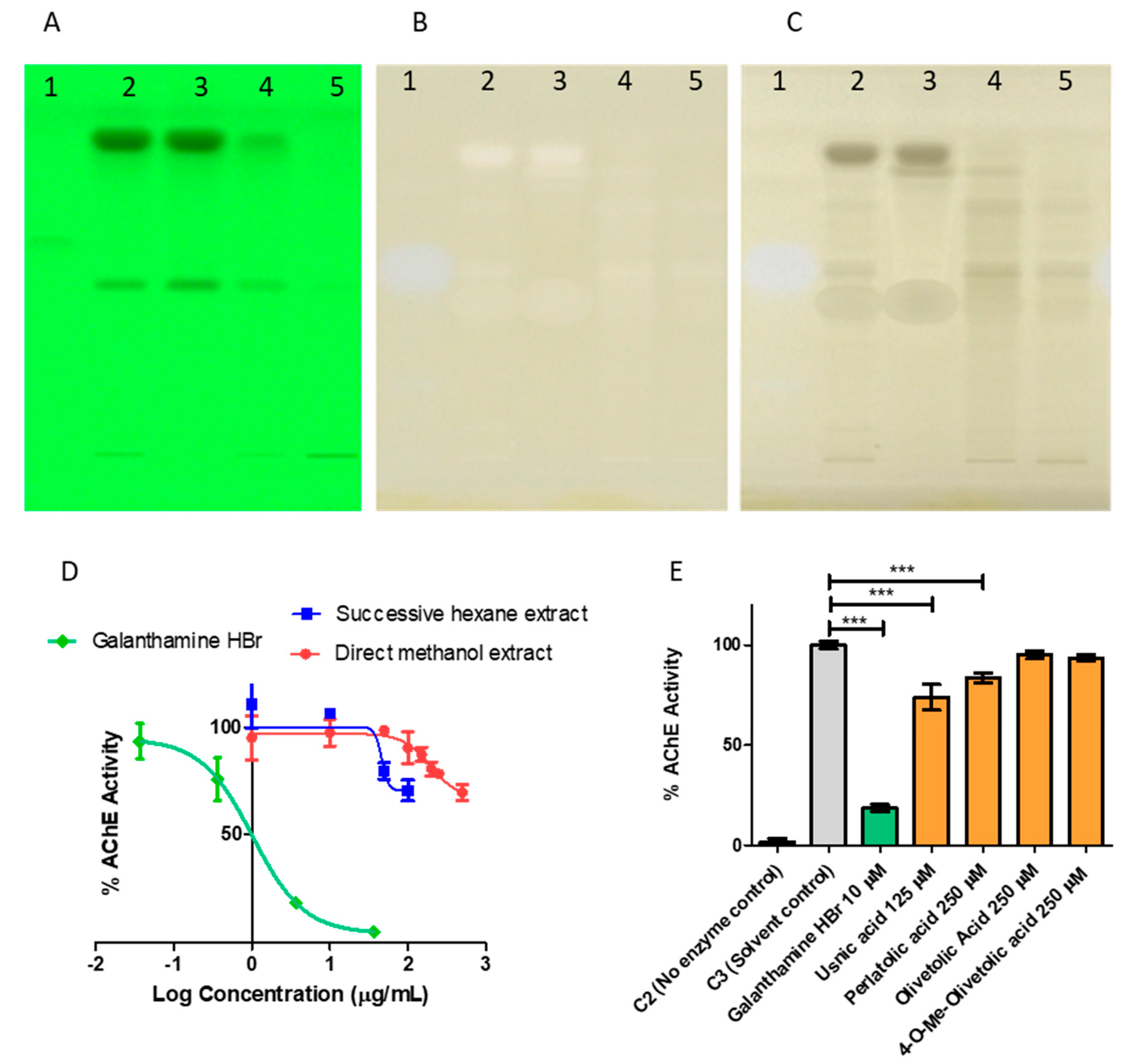
2.4. Probing the Anticholinesterase Activity of Cladonia portentosa

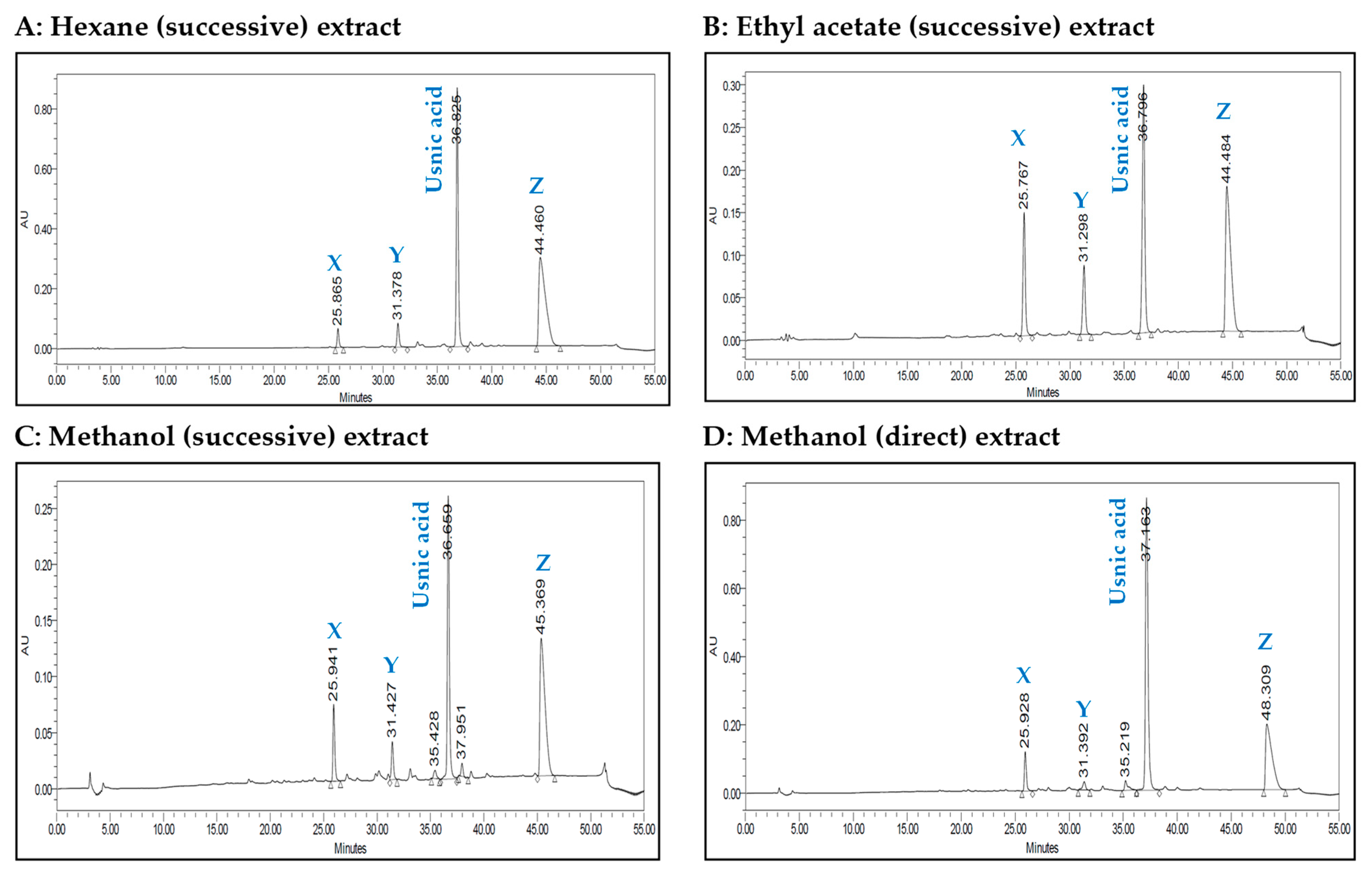{kind=link}
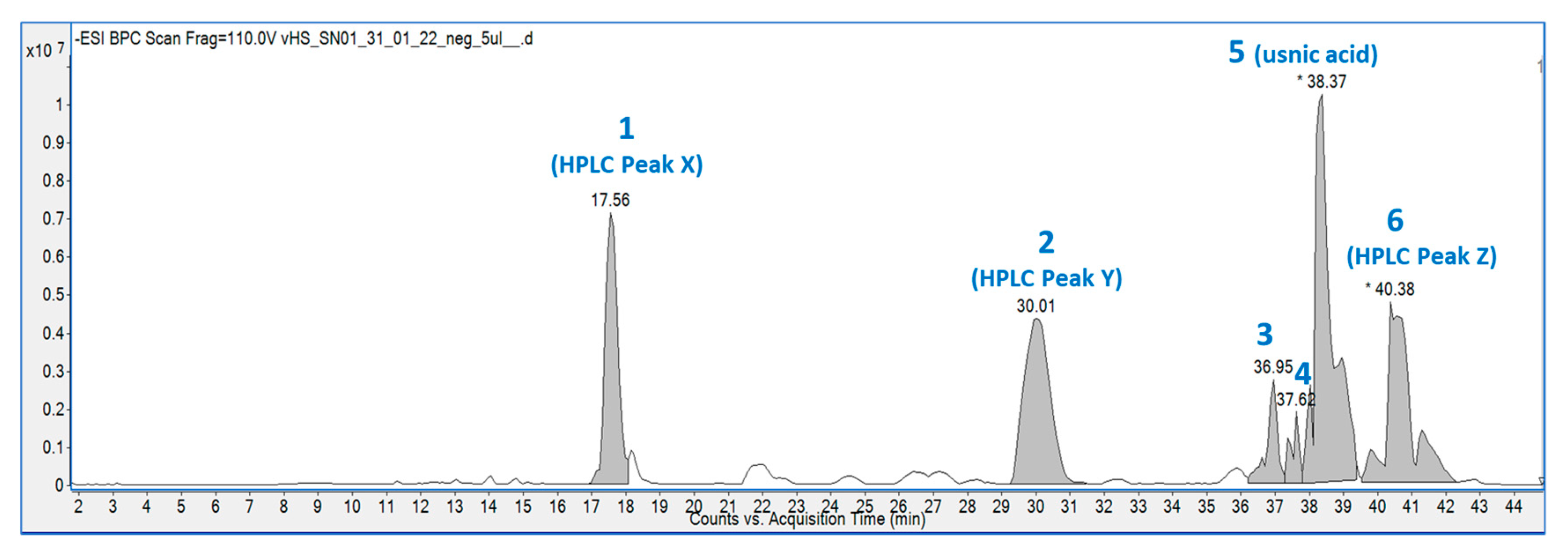{kind=link}
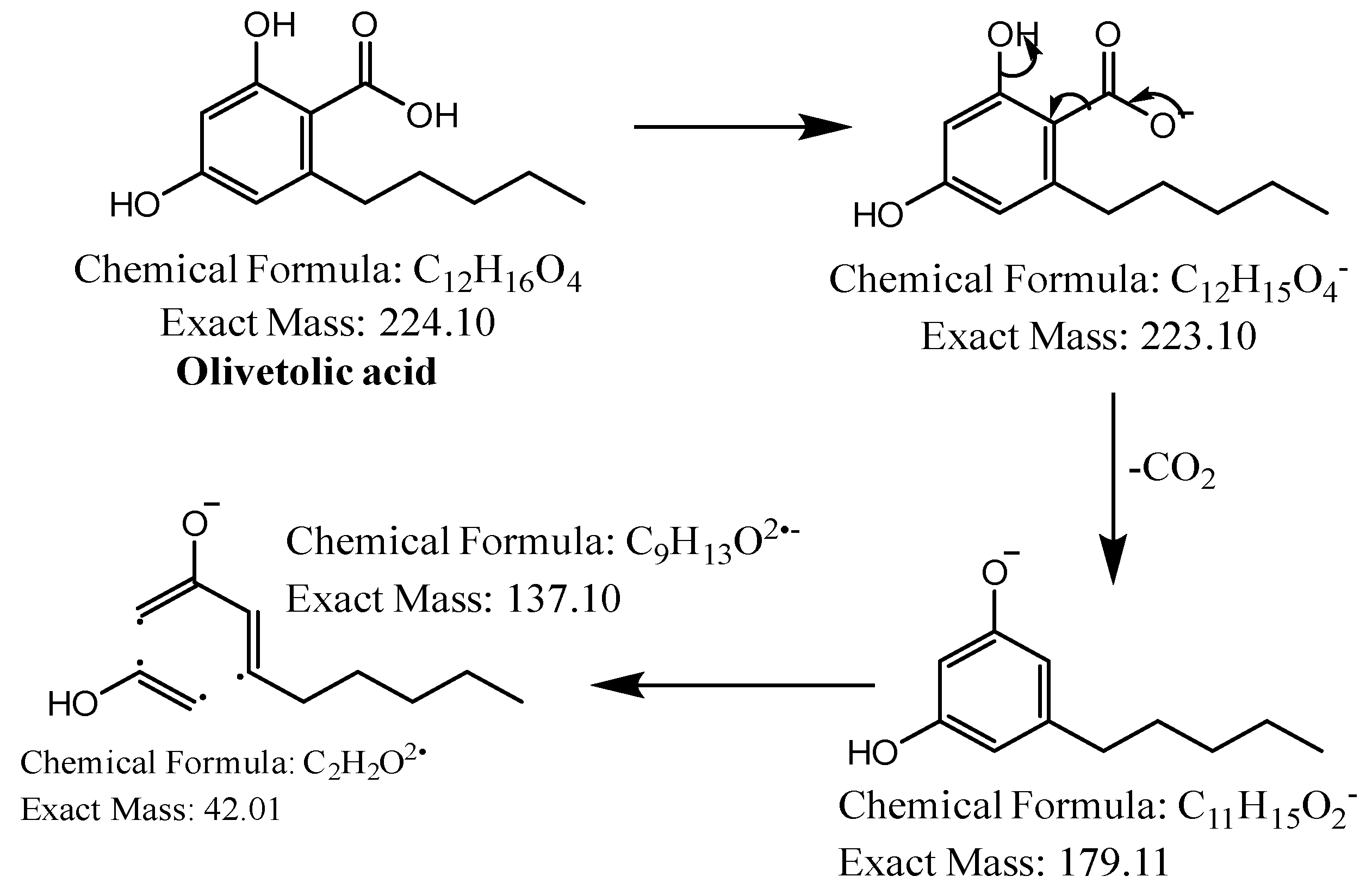{kind=link}
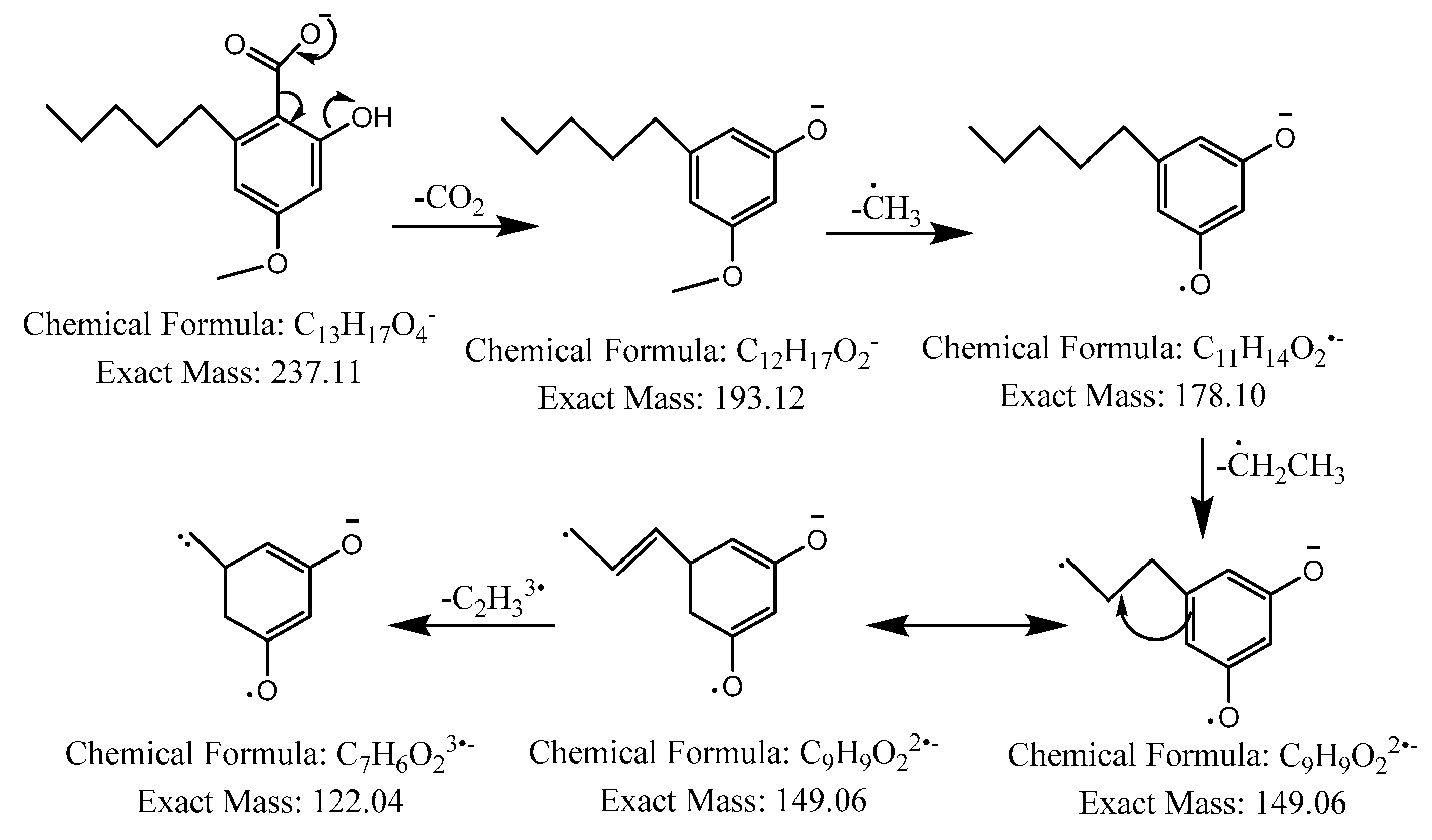{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Usnic Acid | Position | Perlatolic Acid | 4-O-methylolivetolcarboxylic Acid | Olivetolic Acid | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Isolated | Literature [33] | Isolated | Literature [34] | Isolated | Literature [34] | Isolated | Literature [34] | ||||||||||
| δH A | δC A | δH A | δC A | δH A | δC A | δH A | δC B | δH C | δC C | δH C | δC C | δH A | δC A | δH D | δC B | ||
| 1 | - | 197.9 | - | 197.0 | 1 | - | 118.7 | - | 110.9 | - | 111.3 | - | 114.4 | - | 105.2 | - | 104.2 |
| 2 | - | 105.4 | - | 105.8 | 2-OH | - | 156.9 | - | 165.1 | - | 164.9 | - | 165.2 | 10.00 (s) | 163.5 | - | 167.2 |
| 3 | - | 191.4 | - | 190.9 | 3 | 6.59 (d) | 106.8 | 6.57 (d) | 109.2 | 6.33 (d) | 98.9 | 6.35 (s) | 99.0 | 6.12 (d) | 100.5 | 6.14 (d) | 101.6 |
| 3-OH | 11.28 (s) | 11.33 (s) | - | 4 | - | 151.7 | - | 154.9 | - | 166.8 | - | 166.8 | - | - | - | ||
| 4 | 6.31 (s) | 98.3 | 6.31 (s) | 98.8 | 4-OH | - | - | - | - | - | - | - | - | 8.97 (s) | 161.5 | - | 163.4 |
| 4a | 179.3 | 178.8 | 5 | 6.50 (d) | 113.0 | 6.48 (d) | 116.5 | 6.35 (d) | 103.4 | 6.35 (s) | 103.5 | 6.16 (d) | 109.9 | 6.19 (d) | 111.5 | ||
| 5 | - | - | - | - | 6 | - | 143.7 | - | 149.3 | - | 149.5 | - | 149.9 | - | 147.1 | - | 149.8 |
| 5a | 155.4 | - | 155.4 | 7 | - | 166.6 | - | 173.2 | 3.82 (s) | 55.5 | 3.82 (s) | - | - | 172.7 | - | 174.0 | |
| 6 | - | 101.2 | - | 101.1 | 7-OH | - | - | - | - | - | - | - | - | 11.81 (s) | - | - | - |
| 7 | - | 156.7 | - | 157.0 | 8 | 3.74 (s) | 55.2 | 3.73 (s) | 55.8 | - | - | - | - | - | - | - | |
| 7-OH | 13.36 (s) | - | 13.37 (s) | - | 8-OH | - | - | - | - | 11.70 (s) | - | - | - | - | - | - | - |
| 8 | - | 107.2 | - | 107.0 | 9-OH | - | 169.7 | - | - | - | - | - | - | - | - | - | |
| 9 | 162.6 | 162.5 | 1′ | - | 106.8 | - | 105.0 | 2.91 (t) | 36.8 | 2.93 (t) | 36.8 | 2.74 (t) | 35.2 | 2.88 (t) | 37.0 | ||
| 9-OH | 13.36 (s) | 13.37(s) | 2′ | 10.24 (s) | 161.7 | - | 166.4 | 1.59 (m) | 32.1 | 1.60 (m) | 32.1 | 1.47 (m) | 30.9 | 1.57 (m) | 32.2 | ||
| 9a | - | 104.9 | - | 105.1 | 3′ | 6.36 (d) | 98.9 | 6.35 (d) | 99.7 | 1.34 (m) | 31.6 | 1.36 (m) | 31.5 | 1.28 (m) | 31.3 | 1.35 (m) | 32.6 |
| 9b | - | 58.7 | - | 58.2 | 4′ | - | 157.9 | - | 165.5 | 1.34 (m) | 22.6 | 1.36 (m) | 22.6 | 1.28 (m) | 21.8 | 1.35 (m) | 22.9 |
| 10 | 1.74 (s) | 31.5 | 1.74 (s) | 31.6 | 5′ | 6.36 (d) | 111.6 | 6.35 (d) | 111.5 | 0.89 (m) | 14.2 | 0.91 (t) | 14.2 | 0.85 (t) | 13.9 | 0.91 (t) | 14.1 |
| 11 | 2.00 (s) | 7.6 | 2.00 (s) | 7.6 | 6′ | - | 142.9 | - | 148.8 | - | - | - | - | - | - | - | - |
| 12 | - | 201.0 | - | 201.0 | 1a | 2.63 (m) | 33.3 | 2.58 (m) | 36.8 | - | - | - | - | - | - | - | - |
| 13 | 2.67 (s) | 31.1 | 2.67 (s) | 31.1 | 2a | 1.55 (m) | 30.4 | 1.53 (m) | 32.6 | - | - | - | - | - | - | - | - |
| 14 | - | 201.3 | - | 201.0 | 3a | 1.28 (m) | 31.1 | 1.27 (m) | 32.3 | - | - | - | - | - | - | - | - |
| 15 | 2.59 (s) | 27.8 | 2.59 (s) | 28.3 | 4a | 1.28 (m) | 21.8 | 1.27 (m) | 23.0 | - | - | - | - | - | - | - | - |
| - | - | - | - | - | 5a | 0.85 (t) | 13.8 | 0.83 (t) | 14.5 | - | - | - | - | - | - | - | - |
| - | - | - | - | - | 1b | 2.63 (m) | 33.8 | 2.62 (m) | 37.5 | - | - | - | - | - | - | - | - |
| - | - | - | - | - | 2b | 1.55 (m) | 30.8 | 1.53 (m) | 32.8 | - | - | - | - | - | - | - | - |
| - | - | - | - | - | 3b | 1.28 (m) | 31.2 | 1.27 (m) | 32.6 | - | - | - | - | - | - | - | - |
| - | - | - | - | - | 4b | 1.28 (m) | 21.9 | 1.27 (m) | 23.2 | - | - | - | - | - | - | - | - |
| - | - | - | - | - | 5b | 0.85 (m) | 13.9 | 0.84 (t) | 14.3 | - | - | - | - | - | - | - | - |
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Sample Collection and Identification
4.2.2. Soxhlet Extractions
4.2.3. HPLC and LC–MS Analysis
4.2.4. Isolation of Lichenic Acids
4.2.5. Characterisation of Lichenic Acids
4.2.6. Anticholinesterase Activity
HPTLC-Bioautography
Colorimetric Microplate Assay for AChE Inhibition
- ∆ATest = absorbance of a test well at t = 10 min minus absorbance at t = 0 min
- ∆ASolvent control = average absorbance of C3 (solvent control) wells at t = 10 min minus absorbance at t = 0 min
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cocchietto, M.; Skert, N.; Nimis, P.; Sava, G. A review on usnic acid, an interesting natural compound. Naturwissenschaften 2002, 89, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Lauterwein, M.; Oethinger, M.; Belsner, K.; Peters, T.; Marre, R. In vitro activities of the lichen secondary metabolites vulpinic acid, (+)-usnic acid, and (−)-usnic acid against aerobic and anaerobic microorganisms. Antimicrob. Agents Chemother. 1995, 39, 2541–2543. [Google Scholar] [CrossRef] [PubMed]
- Elshobary, M.E.; Osman, M.E.; Abo-Shady, A.M.; Komatsu, E.; Perreault, H.; Sorensen, J.; Piercey-Normore, M.D. Algal carbohydrates affect polyketide synthesis of the lichen-forming fungus Cladonia rangiferina. Mycologia 2016, 108, 646–656. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; Daba, G. Lichens, an alternative drugs for modern diseases. Int. J. Res. Pharm. Biosci. 2019, 6, 5–9. [Google Scholar]
- Albaghli, F.; Zhou, Y.; Hsu, C.C.; Nibali, L. The effect of plaque control in the treatment of oral lichen planus with gingival manifestations: A systematic review. Community Dent. Health 2021, 38, 112. [Google Scholar]
- Gheza, G.; Di Nuzzo, L.; Vallese, C.; Barcella, M.; Benesperi, R.; Giordani, P.; Nascimbene, J.; Assini, S. Morphological and chemical traits of Cladonia respond to multiple environmental factors in acidic dry grasslands. Microorganisms 2021, 9, 453. [Google Scholar] [CrossRef]
- Paukov, A.; Teptina, A.; Morozova, M.; Kruglova, E.; Favero-Longo, S.E.; Bishop, C.; Rajakaruna, N. The effects of edaphic and climatic factors on secondary lichen chemistry: A case study using saxicolous lichens. Diversity 2019, 11, 94. [Google Scholar] [CrossRef]
- Nielsen, K.E.; Andersen, H.V.; Strandberg, M.; Løfstrøm, P.; Degn, H.J.; Damgaard, C. Relationship between atmospheric ammonia concentration and nitrogen content in terricolous lichen (Cladonia portentosa). Water Air Soil Pollut. 2014, 225, 1–8. [Google Scholar] [CrossRef]
- Munzi, S.; Sheppard, L.J.; Leith, I.D.; Cruz, C.; Branquinho, C.; Bini, L.; Gagliardi, A.; Cai, G.; Parrotta, L. The cost of surviving nitrogen excess: Energy and protein demand in the lichen Cladonia portentosa as revealed by proteomic analysis. Planta 2017, 245, 819–833. [Google Scholar] [CrossRef]
- Armstrong, R.A. Adaptation of lichens to extreme conditions. Plant Adapt. Strateg. Chang. Environ. 2017, 1–27. [Google Scholar] [CrossRef]
- Coyle, J.R.; Hurlbert, A.H. Environmental optimality, not heterogeneity, drives regional and local species richness in lichen epiphytes. Glob. Ecol. Biogeogr. 2016, 25, 406–417. [Google Scholar] [CrossRef]
- Lindsay, D.C. The role of lichens in Antarctic ecosystems. Bryologist 1978, 81, 268–276. [Google Scholar] [CrossRef]
- Boustie, J.; Grube, M. Lichens—A promising source of bioactive secondary metabolites. Plant Genet. Resour. 2005, 3, 273–287. [Google Scholar] [CrossRef]
- Peres, M.T.; Cândido, A.C.S.; Faccenda, O.; Gianini, A.S.; Honda, N.K. Phytotoxicity of Perlatolic Acid and Derivatives. Braz. Arch. Biol. Technol. 2016, 59. [Google Scholar] [CrossRef]
- Nagar, S.; Chen, S.; Woulfe, I.; Ruether, M.; O’Connell, P.; Choi, Y.; Sheridan, H. Unlocking Nature’s Pharmacy-the story of an Irish Bog Lichen. Planta Med. 2021, 87, PC1-1. [Google Scholar]
- Cannon, P. Fungi of Great Britain and Ireland. 2013. Available online: http://fungi.myspecies.info/taxonomy/term/4559/descriptions (accessed on 23 March 2023).
- Whelan, P. Lichens of Ireland: An Illustrated Introduction to Over 250 Species; Collins Press: Cork, Ireland, 2011. [Google Scholar]
- Yahr, R.; Vilgalys, R.; Depriest, P. Strong fungal specificity and selectivity for algal symbionts in Florida scrub Cladonia lichens. Mol. Ecol. 2004, 13, 3367–3378. [Google Scholar] [CrossRef] [PubMed]
- Halama, P.; Van Haluwin, C. Antifungal activity of lichen extracts and lichenic acids. BioControl 2004, 49, 95–107. [Google Scholar] [CrossRef]
- Bauer, J.; Waltenberger, B.; Noha, S.M.; Schuster, D.; Rollinger, J.M.; Boustie, J.; Chollet, M.; Stuppner, H.; Werz, O. Discovery of depsides and depsidones from lichen as potent inhibitors of microsomal prostaglandin E2 synthase-1 using pharmacophore models. ChemMedChem 2012, 7, 2077–2081. [Google Scholar] [CrossRef]
- Brandão, L.F.G.; Alcantara, G.B.; Matos, M.D.F.C.; Bogo, D.; dos Santos Freitas, D.; Oyama, N.M.; Honda, N.K. Cytotoxic evaluation of phenolic compounds from lichens against melanoma cells. Chem. Pharm. Bull. 2013, 61, 176–183. [Google Scholar]
- Ezra, L. Investigation of the Antibacterial and Antiviral Properties of Compounds from Natural Sources; University of Surrey: Guildford, UK, 2020. [Google Scholar]
- Galanty, A.; Paśko, P.; Podolak, I. Enantioselective activity of usnic acid: A comprehensive review and future perspectives. Phytochem. Rev. 2019, 18, 527–548. [Google Scholar] [CrossRef]
- Pohanka, M. Inhibitors of acetylcholinesterase and butyrylcholinesterase meet immunity. Int. J. Mol. Sci. 2014, 15, 9809–9825. [Google Scholar] [CrossRef]
- Lane, C.A.; Hardy, J.; Schott, J. Alzheimer’s disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Breijyeh, Z.; Karaman, R. Comprehensive review on Alzheimer’s disease: Causes and treatment. Molecules 2020, 25, 5789. [Google Scholar] [CrossRef] [PubMed]
- Smyrska-Wieleba, N.; Mroczek, T. Natural Inhibitors of Cholinesterases: Chemistry, Structure–Activity and Methods of Their Analysis. Int. J. Mol. Sci. 2023, 24, 2722. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.A.; Khan, N.M.; Ahmad, S.; Aziz, R.; Ullah, I.; Almehmadi, M.; Allahyani, M.; Alsaiari, A.A.; Aljuaid, A. Phytochemical profiling, antioxidant, antimicrobial and cholinesterase inhibitory effects of essential oils isolated from the leaves of Artemisia scoparia and Artemisia absinthium. Pharmaceuticals 2022, 15, 1221. [Google Scholar] [CrossRef] [PubMed]
- Kekuda, T.P.; Lavanya, D.; Pooja, R. Lichens as promising resources of enzyme inhibitors: A review. J. Drug Deliv. Ther. 2019, 9, 665–676. [Google Scholar]
- Luo, H.; Li, C.; Kim, J.C.; Liu, Y.; Jung, J.S.; Koh, Y.J.; Hur, J.S. Biruloquinone, an acetylcholinesterase inhibitor produced by lichen-forming fungus Cladonia macilenta. J. Microbiol. Biotechnol. 2013, 23, 161–166. [Google Scholar] [CrossRef]
- Rhee, I.K.; van de Meent, M.; Ingkaninan, K.; Verpoorte, R. Screening for acetylcholinesterase inhibitors from Amaryllidaceae using silica gel thin-layer chromatography in combination with bioactivity staining. J. Chromatogr. A 2001, 915, 217–223. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Petrová, K.; Bačkorová, M.; Demčišáková, Z.; Petrovová, E.; Goga, M.; Vilková, M.; Frenák, R.; Bačkor, M.; Mojžiš, J.; Kello, M. Usnic acid isolated from Usnea antarctica (Du Rietz) reduced in vitro angiogenesis in VEGF-and bFGF-stimulated HUVECs and ex ovo in quail chorioallantoic membrane (CAM) Assay. Life 2022, 12, 1444. [Google Scholar] [CrossRef]
- Huneck, S.; Yoshimura, I.; Huneck, S.; Yoshimura, I. Identification of Lichen Substances; Springer: Berlin Heidelberg, Germany, 1996. [Google Scholar]
- Cakmak, K.C.; Gülçin, İ. Anticholinergic and antioxidant activities of usnic acid-an activity-structure insight. Toxicol. Rep. 2019, 6, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Reddy, R.G.; Veeraval, L.; Maitra, S.; Chollet-Krugler, M.; Tomasi, S.; Lohezic-Le Devehat, F.; Boustie, J.; Chakravarty, S. Lichen-derived compounds show potential for central nervous system therapeutics. Phytomedicine 2016, 23, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Le Pogam, P.; Schinkovitz, A.; Legouin, B.; Le Lamer, A.C.; Boustie, J.; Richomme, P. Matrix-free UV-laser desorption ionization mass spectrometry as a versatile approach for accelerating dereplication studies on lichens. Anal. Chem. 2015, 87, 10421–10428. [Google Scholar] [CrossRef] [PubMed]
- Areche, C.; Parra, J.R.; Sepulveda, B.; García-Beltrán, O.; Simirgiotis, M.J. UHPLC-MS metabolomic fingerprinting, antioxidant, and enzyme inhibition activities of Himantormia lugubris from Antarctica. Metabolites 2022, 12, 560. [Google Scholar] [CrossRef]
- Cornejo, A.; Salgado, F.; Caballero, J.; Vargas, R.; Simirgiotis, M.; Areche, C. Secondary Metabolites in Ramalina terebrata Detected by UHPLC/ESI/MS/MS and Identification of Parietin as Tau Protein Inhibitor. Int. J. Mol. Sci. 2016, 17, 1303. [Google Scholar] [CrossRef]
- Millot, M.; Dieu, A.; Tomasi, S. Dibenzofurans and derivatives from lichens and ascomycetes. Nat. Prod. Rep. 2016, 33, 801–811. [Google Scholar] [CrossRef]
- Connolly, J.D.; Freer, A.; Huneck, S. Crystal structure of (−)-placodiolic acid, a dibenzofuran derivative from the lichen Rhizoplaca chrysoleuca. Phytochemistry 1984, 23, 702. [Google Scholar] [CrossRef]
- Millot, M.; Girardot, M.; Dutreix, L.; Mambu, L.; Imbert, C. Antifungal and anti-biofilm activities of acetone lichen extracts against Candida albicans. Molecules 2017, 22, 651. [Google Scholar] [CrossRef]
- Shishido, T.K.; Wahlsten, M.; Laine, P.; Rikkinen, J.; Lundell, T.; Auvinen, P. Microbial Communities of Cladonia Lichens and Their Biosynthetic Gene Clusters Potentially Encoding Natural Products. Microorganisms 2021, 9, 1347. [Google Scholar] [CrossRef]
- Kutney, J.P.; Sanchez, I.; Yee, T. Mass spectral fragmentation studies in usnic acid and related compounds. Org. Mass Spectrom. 1974, 8, 129–146. [Google Scholar] [CrossRef]
- Xu, M.; Heidmarsson, S.; Thorsteinsdottir, M.; Eiriksson, F.F.; Omarsdottir, S.; Olafsdottir, E.S. DNA barcoding and LC-MS metabolite profiling of the lichen-forming genus Melanelia: Specimen identification and discrimination focusing on Icelandic taxa. PLoS ONE 2017, 12, e0178012. [Google Scholar] [CrossRef] [PubMed]
- Taibon, J.; Ankli, A.; Schwaiger, S.; Magnenat, C.; Boka, V.I.; Simões-Pires, C.; Aligiannis, N.; Cuendet, M.; Skaltsounis, A.L.; Reich, E.; et al. Prevention of false-positive results: Development of an HPTLC autographic assay for the detection of natural tyrosinase inhibitors. Planta Med. 2015, 81, 1198–1204. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, Y.; Wang, R.; Wang, Z.; Yang, B.; Kuang, H. An evolving technology that integrates classical methods with continuous technological developments: Thin-layer chromatography bioautography. Molecules 2021, 26, 4647. [Google Scholar] [CrossRef] [PubMed]
- Studzińska-Sroka, E.; Majchrzak-Celińska, A.; Zalewski, P.; Szwajgier, D.; Baranowska-Wójcik, E.; Kaproń, B.; Plech, T.; Żarowski, M.; Cielecka-Piontek, J. Lichen-Derived Compounds and Extracts as Biologically Active Substances with Anticancer and Neuroprotective Properties. Pharmaceuticals 2021, 14, 1293. [Google Scholar] [CrossRef] [PubMed]
- Egbert, S.; Hoffman, J.R.; McMullin, R.T.; Lendemer, J.C.; Sorensen, J.L. Unraveling usnic acid: A comparison of biosynthetic gene clusters between two reindeer lichen (Cladonia rangiferina and C. uncialis). Fungal Biol. 2022, 126, 697–706. [Google Scholar] [CrossRef]
- Pinho, B.R.; Ferreres, F.; Valentão, P.; Andrade, P.B. Nature as a source of metabolites with cholinesterase-inhibitory activity: An approach to Alzheimer’s disease treatment. J. Pharm. Pharmacol. 2013, 65, 1681–1700. [Google Scholar] [CrossRef]
- Polovinka, M.P.; Komarova, N.I.; Korchagina, D.V.; Sokolov, D.N.; Luzina, O.A.; Vlasenko, N.G.; Malyuga, A.A.; Romanova, E.V.; Salakhutdinov, N.F. Secondary metabolites of the lichen Cladonia stellaris. Chem. Nat. Compd. 2012, 48, 392–395. [Google Scholar] [CrossRef]
- González, A.G.; Barrera, J.B.; Ma Rodríguez Pérez, E.; Hernández Padrón, C.E. Chemical constituents of the lichen Cladina macaronesica. Z. Für Nat. C 1991, 46, 12–18. [Google Scholar] [CrossRef]
- Culberson, C.F. Biogenetic relationships of the lichen substances in the framework of systematics. Bryologist 1986, 89, 91–98. [Google Scholar] [CrossRef]
- Herz, W.; Grisebach, H.; Kirby, G.W.; Elix, J.A.; Whitton, A.A.; Sargent, M.V. Recent Progress in the Chemistry of Lichen Substances; Springer: Vienna, Austria, 1984. [Google Scholar]
- Abdolshahi, A.; Majd, M.H.; Rad, J.S.; Taheri, M.; Shabani, A.; Teixeira da Silva, J.A. Choice of solvent extraction technique affects fatty acid composition of pistachio (Pistacia vera L.) oil. J. Food Sci. Technol. 2015, 52, 2422–2427. [Google Scholar] [CrossRef]
- Kim, H.K.; Choi, Y.; Verpoorte, R. NMR-based metabolomic analysis of plants. Nat. Protoc. 2010, 5, 536–549. [Google Scholar] [CrossRef] [PubMed]










| S. No. | Extract | Direct/Successive | Yield (%) |
|---|---|---|---|
| 1 | Methanol | Direct, Soxhlet | 14.40 |
| 2 | Hexane | Successive, Soxhlet | 1.36 |
| 3 | Ethyl acetate | Successive, Soxhlet | 0.98 |
| 4 | Methanol | Successive, Soxhlet | 7.56 |
| No | Ion.+/- | RT [min] | Proposed Structure | Molecular Formula | m/z Theoretical | m/z Experimental | Error | DBE | MS/MS Fragmentation | Proposed Compound |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | - | 17.6 |  | C12H16O4 | 223.0976 | 223.0997 | −9.45 | 5 | 205, 179, 137 | Olivetolic acid (1) |
| 2 | - | 29.9 |  | C13H18O4 | 237.1132 | 237.1158 | −10.78 | 5 | 193, 178, 149, 122 | 4-O-methylolivetolcarboxylic acid (2) |
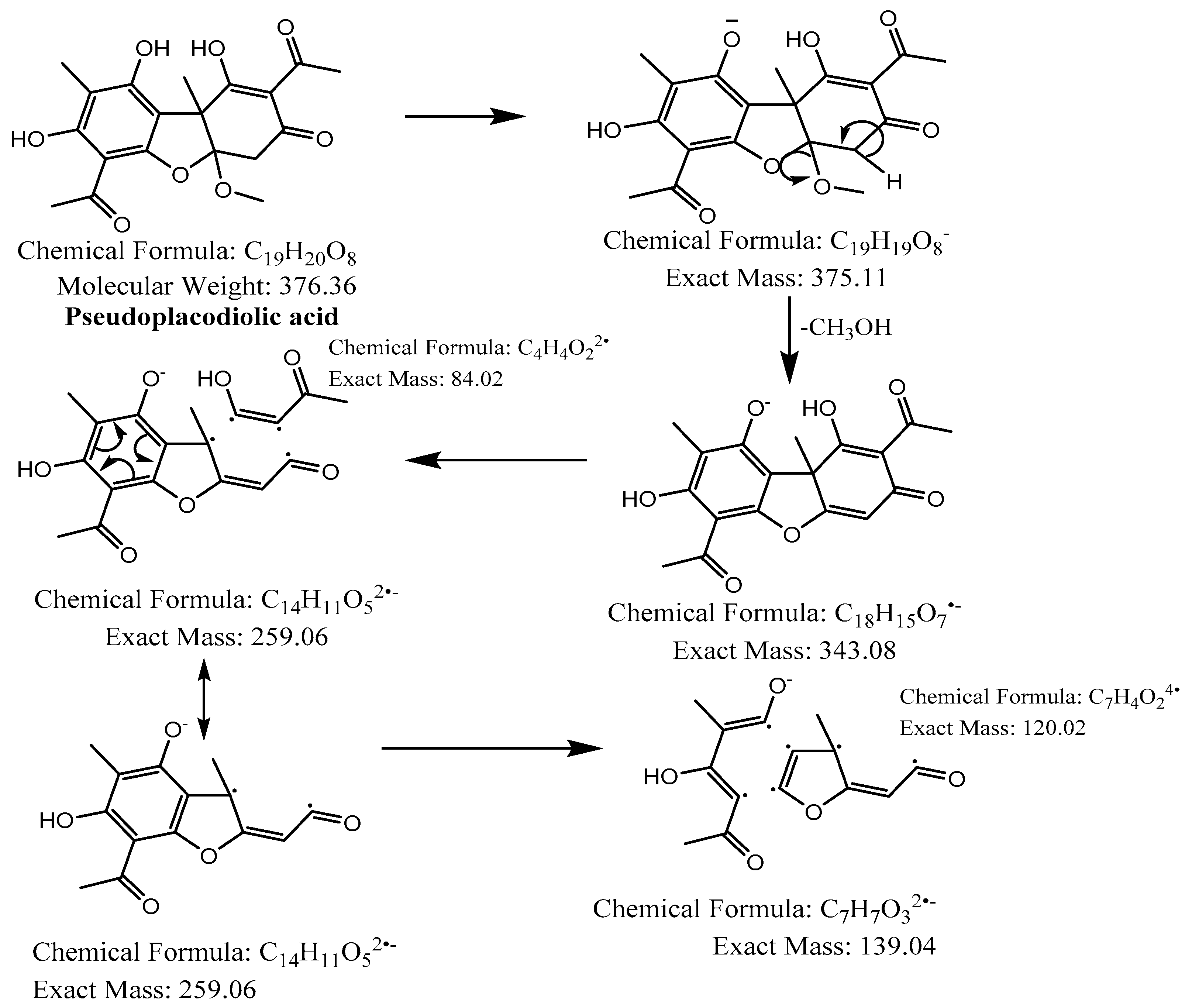
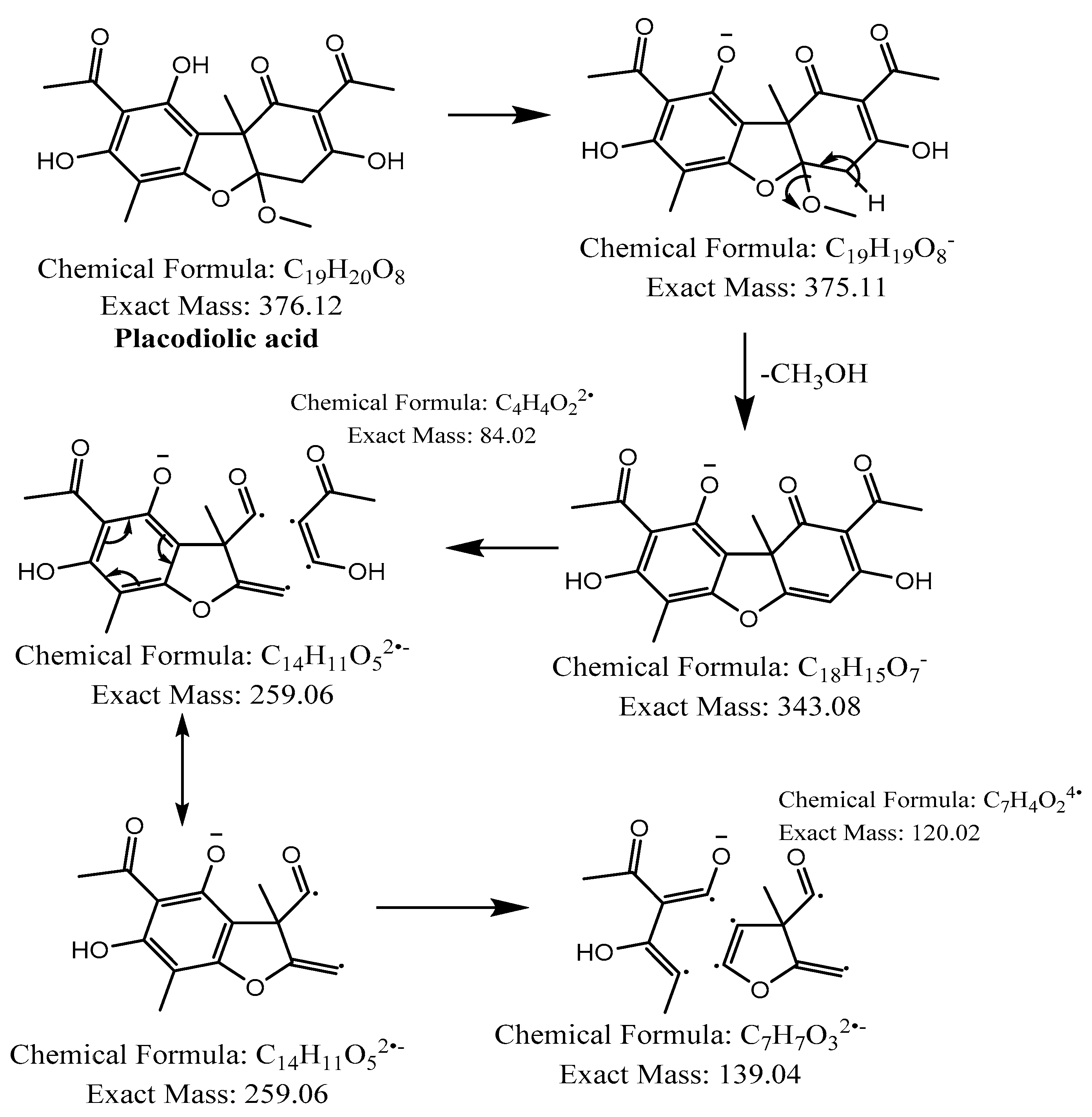
| 3,4 | - | 36.9 and 37.6 |  | C19H20O8 | 375.1085 | 375.1134 | −10.26 | 10 | 343, 299, 259, 180, 139 | Placodiolic acid (3) or Pseudoplacodiolic acid (4) |
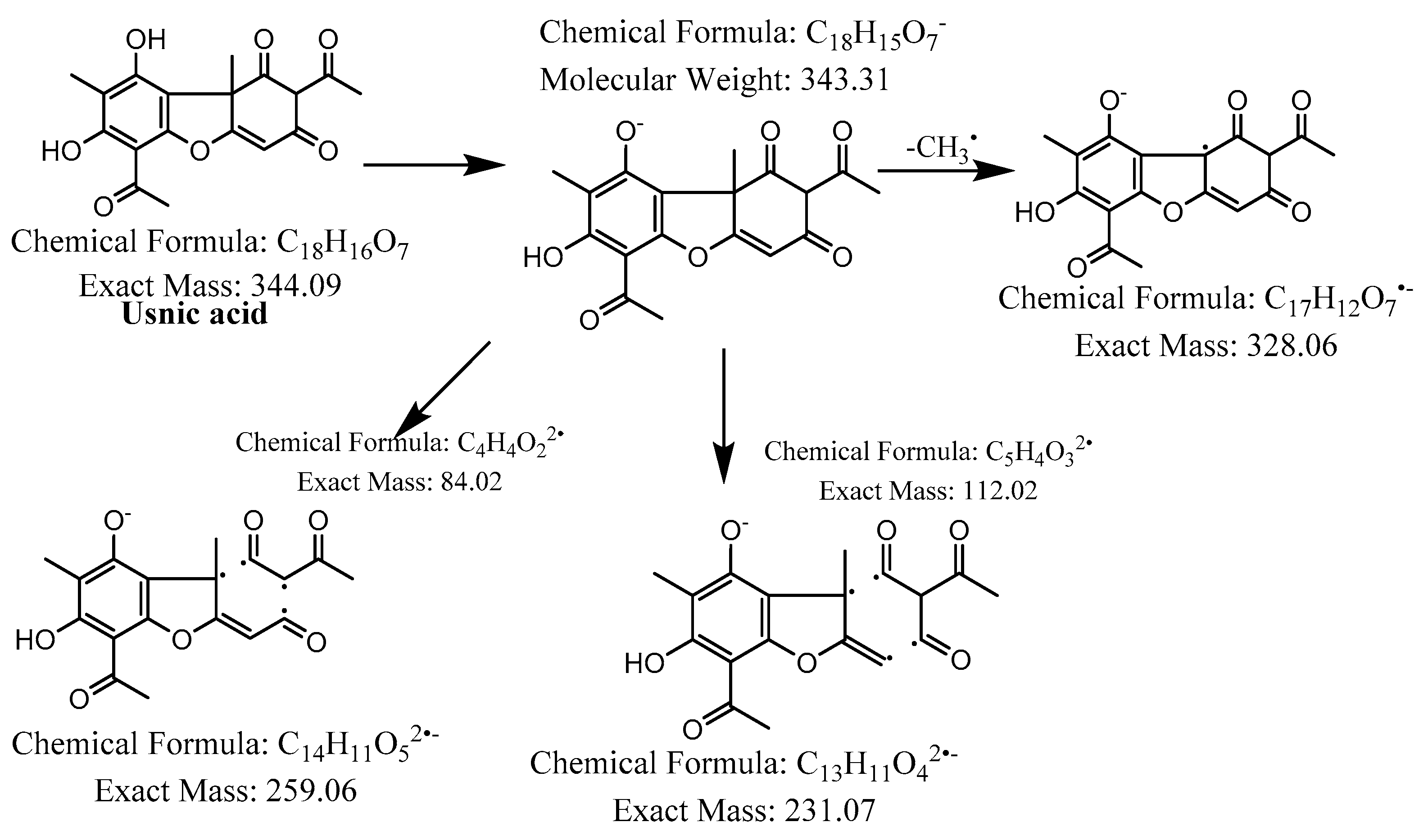
| 5 | - | 38.2 |  | C18H16O7 | 343.0955 | 343.0862 | −11.3 | 11 | 328, 299, 259, 231 | Usnic acid (5) |
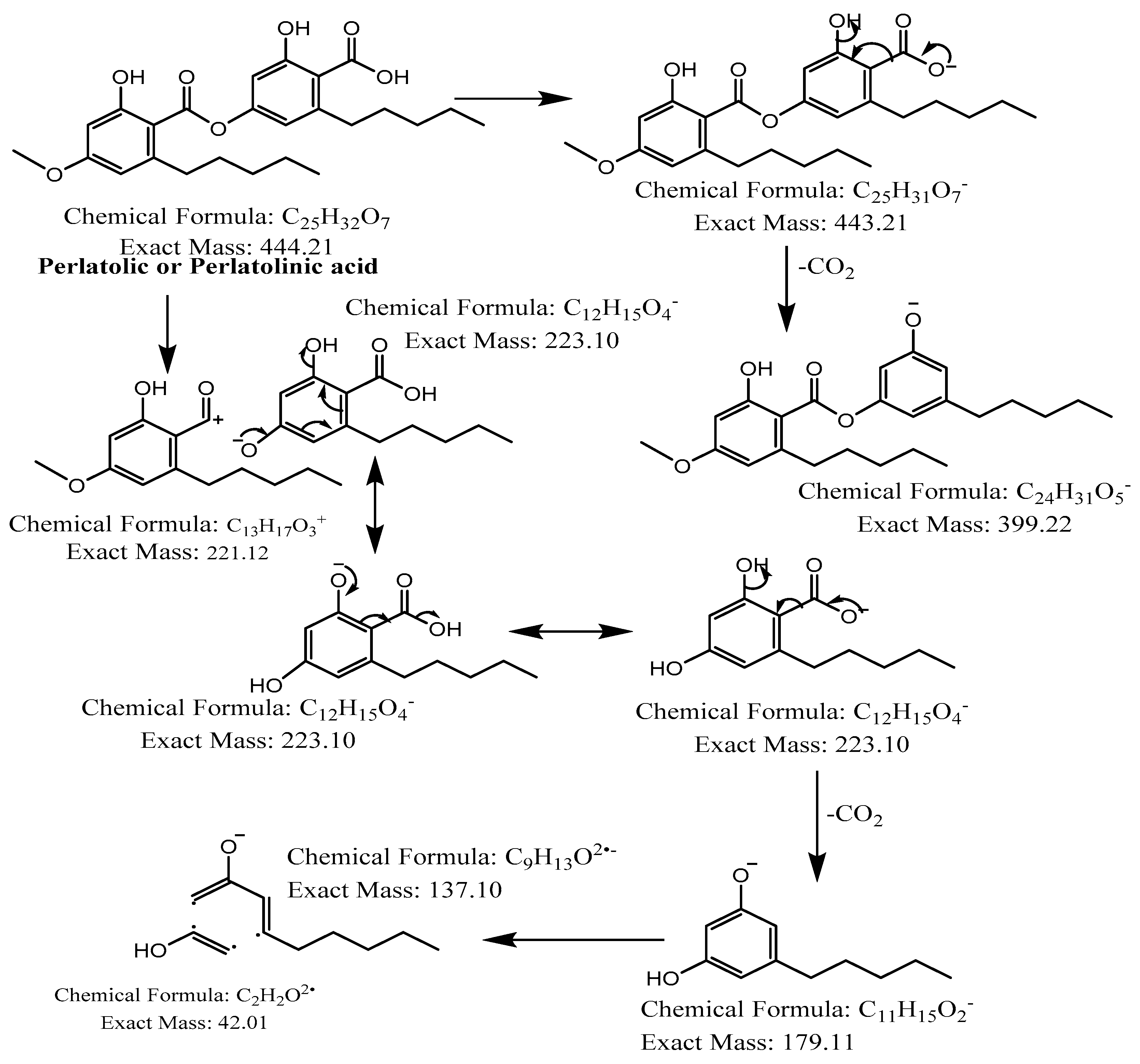
| 6 | - | 40.3 |  | C25H32O7 | 443.2075 | 443.2109 | −7.59 | 10 | 399, 355, 223, 205, 179 | Perlatolic acid (6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagar, S.; Pigott, M.; Kukula-Koch, W.; Sheridan, H. Unravelling Novel Phytochemicals and Anticholinesterase Activity in Irish Cladonia portentosa. Molecules 2023, 28, 4145. https://doi.org/10.3390/molecules28104145
Nagar S, Pigott M, Kukula-Koch W, Sheridan H. Unravelling Novel Phytochemicals and Anticholinesterase Activity in Irish Cladonia portentosa. Molecules. 2023; 28(10):4145. https://doi.org/10.3390/molecules28104145
Chicago/Turabian StyleNagar, Shipra, Maria Pigott, Wirginia Kukula-Koch, and Helen Sheridan. 2023. "Unravelling Novel Phytochemicals and Anticholinesterase Activity in Irish Cladonia portentosa" Molecules 28, no. 10: 4145. https://doi.org/10.3390/molecules28104145
APA StyleNagar, S., Pigott, M., Kukula-Koch, W., & Sheridan, H. (2023). Unravelling Novel Phytochemicals and Anticholinesterase Activity in Irish Cladonia portentosa. Molecules, 28(10), 4145. https://doi.org/10.3390/molecules28104145