
Proteomic Signature of Extracellular Vesicles Associated with Colorectal Cancer


Abstract
:
1. Introduction
2. Results
2.1. Targeted Proteomic Analysis of EV-Associated Proteins in HT29, HCT-116, and CaCo-2 Cell Lines
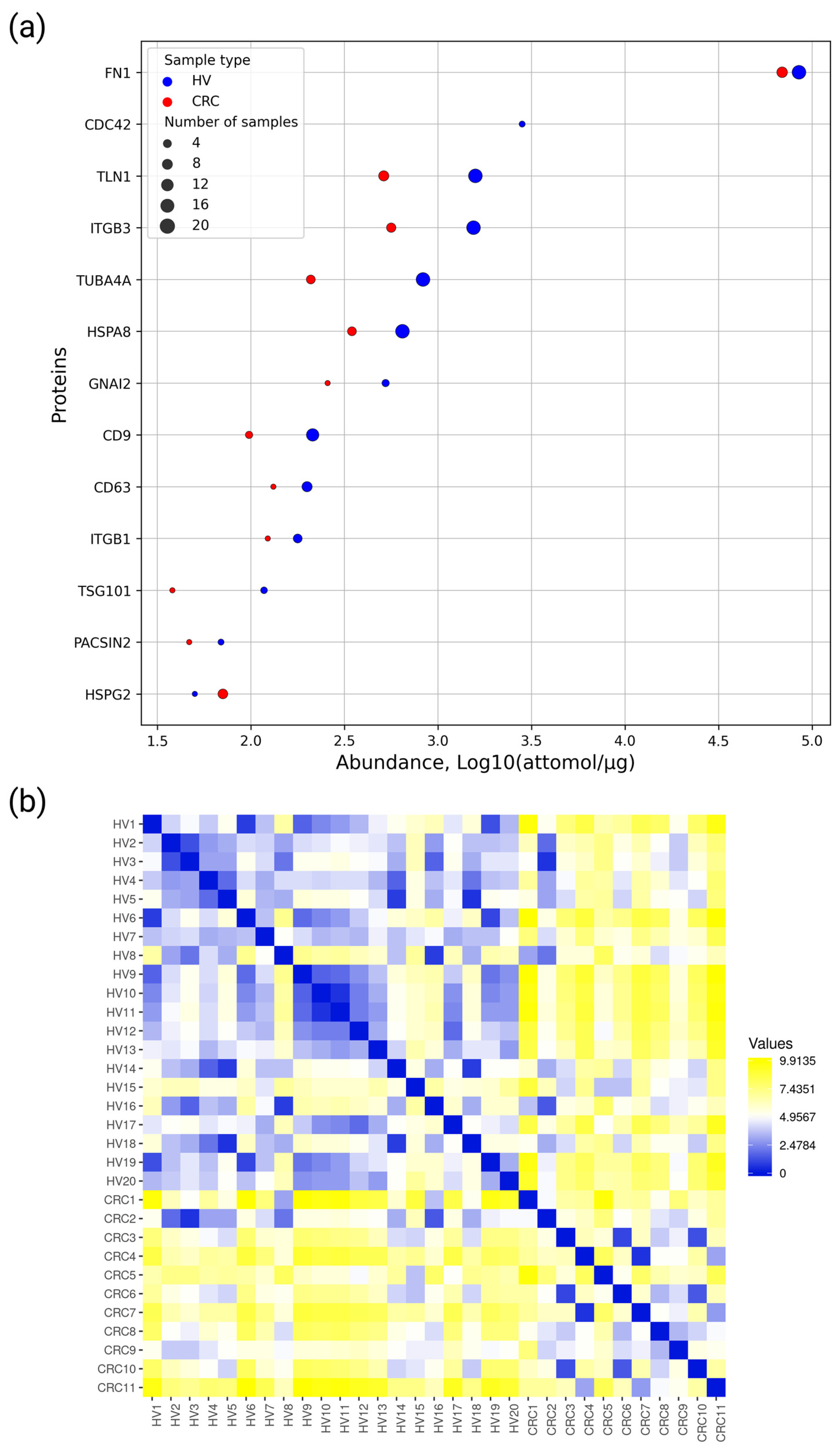
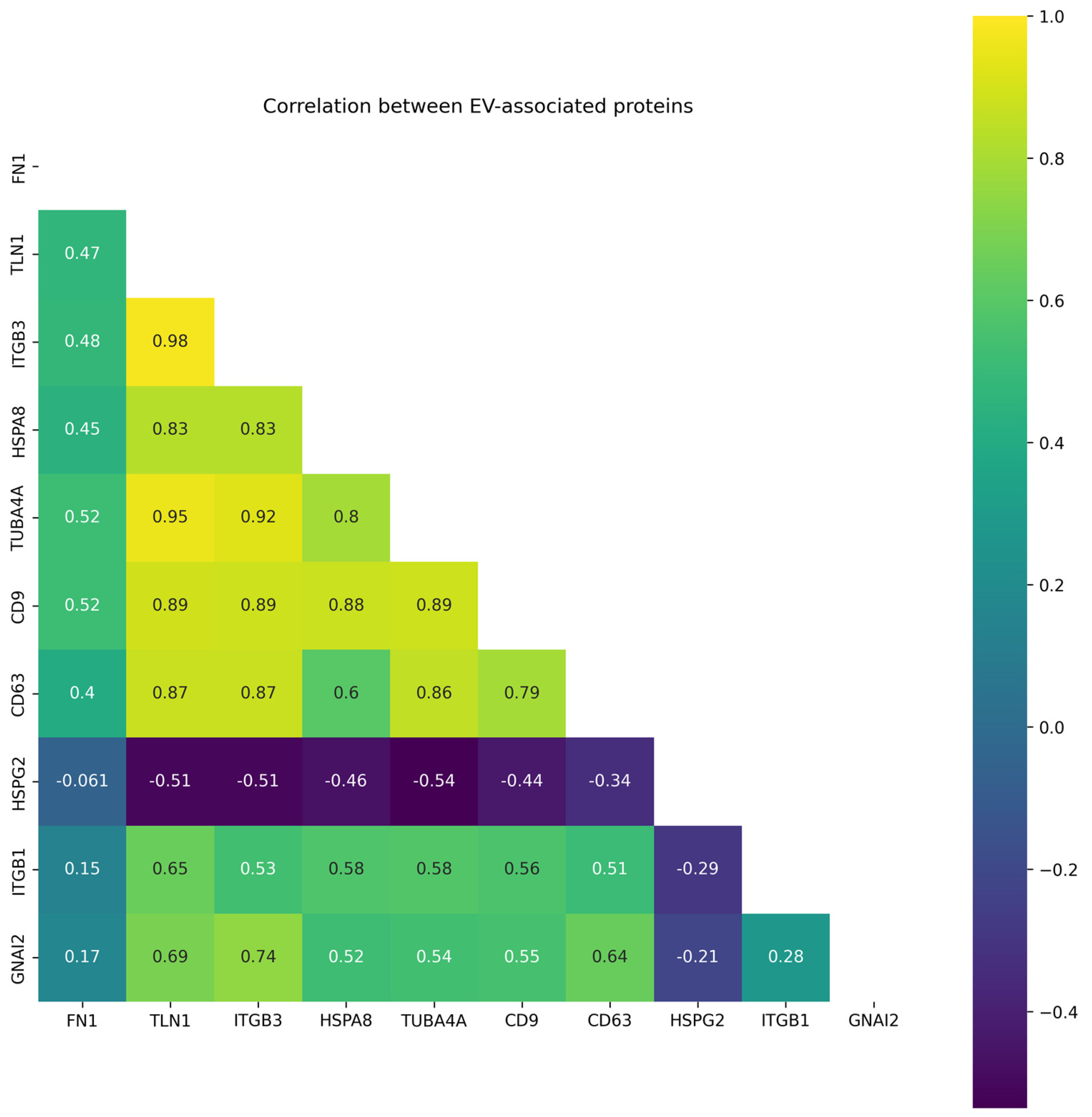
2.2. Targeted Proteomic Analysis of EV-Associated Proteins in EV Samples Derived from Blood Plasma of CRC Patients and Healthy Volunteers
3. Discussion
4. Materials and Methods
4.1. Cultivation of HT29, HCT-116 and CaCo-2 Cell Lines and EVs Isolation
4.2. Clinical Sample Description
4.3. EV Isolation from Blood Plasma and Protein Extraction
4.4. Sample Preparation for Mass Spectrometry Analysis
4.5. Synthesis of SIS
4.6. Quantitative Analysis of EV-Associated Proteins by Targeted Mass Spectrometry
4.7. Statistical and Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Aykan, N.F. Red Meat and Colorectal Cancer. Oncol. Rev. 2015, 9, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal Cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, H.; Kuroda, H.; Imai, Y.; Hiraishi, H. Molecular Pathogenesis of Sporadic Colorectal Cancers. Chin. J. Cancer 2016, 35, 4. [Google Scholar] [CrossRef]
- Holinski-Feder, E.; Morak, M. Familial Adenomatous Polyposis and Other Polyposis Syndromes. Gastroenterologe 2010, 5, 7–15. [Google Scholar] [CrossRef]
- Bien, S.A.; Su, Y.R.; Conti, D.V.; Harrison, T.A.; Qu, C.; Guo, X.; Lu, Y.; Albanes, D.; Auer, P.L.; Banbury, B.L.; et al. Genetic Variant Predictors of Gene Expression Provide New Insight into Risk of Colorectal Cancer. Hum. Genet. 2019, 138, 307–326. [Google Scholar] [CrossRef] [PubMed]
- Worthley, D.L.; Leggett, B.A. Colorectal Cancer: Molecular Features and Clinical Opportunities. Clin. Biochem. Rev. 2010, 31, 31–38. [Google Scholar]
- Grady, W.M. Genomic Instability and Colon Cancer; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; Volume 23. [Google Scholar]
- Nojadeh, J.N.; Sharif, S.B.; Sakhinia, E. Microsatellite Instability in Colorectal Cancer. EXCLI J. 2018, 17, 159–168. [Google Scholar]
- Mojarad, E.N.; Kuppen, P.J.K.; Aghdaei, H.A.; Zali, M.R. The CpG Island Methylator Phenotype (CIMP) in Colorectal Cancer. Gastroenterol. Hepatol. Bed Bench 2013, 6, 120–128. [Google Scholar]
- Ahmed, M. Colon Cancer: A Clinician’s Perspective in 2019. Rev. Gastroenterol. Res. 2020, 13, 1–10. [Google Scholar] [CrossRef]
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedźwiedzka, E.; Arłukowicz, T.; Przybyłowicz, K.E. A Review of Colorectal Cancer in Terms of Epidemiology, Risk Factors, Development, Symptoms and Diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef]
- Reumkens, A.; Rondagh, E.J.A.; Bakker, C.M.; Winkens, B.; Masclee, A.A.M.; Sanduleanu, S. Post-Colonoscopy Complications: A Systematic Review, Time Trends, and Meta-Analysis of Population-Based Studies. Am. J. Gastroenterol. 2016, 111, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Li, Y.; Wang, M.; Gu, J.; Xu, W.; Cai, H.; Fang, X.; Zhang, X. Exosomes as a New Frontier of Cancer Liquid Biopsy. Mol. Cancer 2022, 21, 56. [Google Scholar] [CrossRef] [PubMed]
- Holdenrieder, S.; Pagliaro, L.; Morgenstern, D.; Dayyani, F. Clinically Meaningful Use of Blood Tumor Markers in Oncology. Biomed. Res. Int. 2016, 2016, 9795269. [Google Scholar] [CrossRef]
- Topdagi, O.; Timuroglu, A. Evaluation of the Relationship between Carcinoembryonic Antigen and TNM Stage in Colorectal Cancer. Eurasian J. Med. 2018, 50, 96–98. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Hurley, J.; Roberts, D.; Chakrabortty, S.K.; Enderle, D.; Noerholm, M.; Breakefield, X.O.; Skog, J.K. Exosome-Based Liquid Biopsies in Cancer: Opportunities and Challenges. Ann. Oncol. 2021, 32, 466–477. [Google Scholar] [CrossRef]
- Strotman, L.N.; Millner, L.M.; Valdes, R.; Linder, M.W. Liquid Biopsies in Oncology and the Current Regulatory Landscape. Mol. Diagn. Ther. 2016, 20, 429–436. [Google Scholar] [CrossRef]
- Carnino, J.M.; Hao Kwok, Z.; Jin, Y. Extracellular Vesicles: A Novel Opportunity for Precision Medicine in Respiratory Diseases. Front. Med. 2021, 8, 661679. [Google Scholar] [CrossRef]
- Xie, D.; Qian, B.; Li, X. Nucleic Acids and Proteins Carried by Exosomes from Various Sources: Potential Role in Liver Diseases. Front. Physiol. 2022, 13, 957036. [Google Scholar] [CrossRef]
- Vella, L.J.; Sharples, R.A.; Nisbet, R.M.; Cappai, R.; Hill, A.F. The Role of Exosomes in the Processing of Proteins Associated with Neurodegenerative Diseases. Eur. Biophys. J. 2008, 37, 323–332. [Google Scholar] [CrossRef]
- Zhang, Y.; Fonslow, B.R.; Shan, B.; Baek, M.-C.; Yates, J.R. Protein Analysis by Shotgun Proteomics. Mass Spectrom. Chem. Proteom. 2013, 113, 2343–2394. [Google Scholar] [CrossRef]
- Zheng, X.; Xu, K.; Zhou, B.; Chen, T.; Huang, Y.; Li, Q.; Wen, F.; Ge, W.; Wang, J.; Yu, S.; et al. A Circulating Extracellular Vesicles-Based Novel Screening Tool for Colorectal Cancer Revealed by Shotgun and Data-Independent Acquisition Mass Spectrometry. J. Extracell. Vesicles 2020, 9, 1750202. [Google Scholar] [CrossRef] [PubMed]
- Atak, A.; Khurana, S.; Gollapalli, K.; Reddy, P.J.; Levy, R.; Ben-Salmon, S.; Hollander, D.; Donyo, M.; Heit, A.; Hotz-Wagenblatt, A.; et al. Quantitative Mass Spectrometry Analysis Reveals a Panel of Nine Proteins as Diagnostic Markers for Colon Adenocarcinomas. Oncotarget 2018, 9, 13530–13544. [Google Scholar] [CrossRef]
- Heck, K.A.; Lindholm, H.T.; Niederdorfer, B.; Tsirvouli, E.; Kuiper, M.; Flobak, Å.; Lægreid, A.; Thommesen, L. Characterisation of Colorectal Cancer Cell Lines through Proteomic Profiling of Their Extracellular Vesicles. Proteomes 2023, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Kopylov, A.T.; Ponomarenko, E.A.; Ilgisonis, E.V.; Pyatnitskiy, M.A.; Lisitsa, A.V.; Poverennaya, E.V.; Kiseleva, O.I.; Farafonova, T.E.; Tikhonova, O.V.; Zavialova, M.G.; et al. 200+ Protein Concentrations in Healthy Human Blood Plasma: Targeted Quantitative SRM SIS Screening of Chromosomes 18, 13, Y, and the Mitochondrial Chromosome Encoded Proteome. J. Proteome Res. 2019, 18, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Novikova, S.E.; Vakhrushev, I.V.; Tsvetkova, A.V.; Shushkova, N.A.; Farafonova, T.E.; Yarygin, K.N.; Zgoda, V.G. Proteomics of Transcription Factors: Identification of Pool of HL-60 Cell Line-Specific Regulatory Proteins. Biomeditsinskaya Khimiya 2019, 65, 294–305. [Google Scholar] [CrossRef]
- Song, E.; Gao, Y.; Wu, C.; Shi, T.; Nie, S.; Fillmore, T.L.; Schepmoes, A.A.; Gritsenko, M.A.; Qian, W.J.; Smith, R.D.; et al. Targeted Proteomic Assays for Quantitation of Proteins Identified by Proteogenomic Analysis of Ovarian Cancer. Sci. Data 2017, 4, 170091. [Google Scholar] [CrossRef]
- Novikova, S.E.; Farafonova, T.E.; Tikhonova, O.V.; Shushkova, N.A.; Pyatnitsky, M.A.; Zgoda, V.G.; Ponomarenko, E.A.; Lisitsa, A.V.; Grigoryev, A.I.; Tutelyan, V.A.; et al. Mass-Spectrometric MRM Analysis of FDA-Approved Proteins in Plasma of Healthy Volunteers. Biochem. Suppl. Ser. B Biomed. Chem. 2021, 15, 40–61. [Google Scholar] [CrossRef]
- Shiromizu, T.; Kume, H.; Ishida, M.; Adachi, J.; Kano, M.; Matsubara, H.; Tomonaga, T. Quantitation of Putative Colorectal Cancer Biomarker Candidates in Serum Extracellular Vesicles by Targeted Proteomics. Sci. Rep. 2017, 7, 12782. [Google Scholar] [CrossRef]
- Dash, S.; Wu, C.C.; Wu, C.C.; Chiang, S.F.; Lu, Y.T.; Yeh, C.Y.; You, J.F.; Chu, L.J.; Yeh, T.S.; Yu, J.S. Extracellular Vesicle Membrane Protein Profiling and Targeted Mass Spectrometry Unveil CD59 and Tetraspanin 9 as Novel Plasma Biomarkers for Detection of Colorectal Cancer. Cancers 2023, 15, 177. [Google Scholar] [CrossRef]
- Novikova, S.; Shushkova, N.; Farafonova, T.; Tikhonova, O.; Kamyshinsky, R.; Zgoda, V. Proteomic Approach for Searching for Universal, Tissue-Specific, and Line-Specific Markers of Extracellular Vesicles in Lung and Colorectal Adenocarcinoma Cell Lines. Int. J. Mol. Sci. 2020, 21, 6601. [Google Scholar] [CrossRef]
- Irmer, B.; Chandrabalan, S.; Maas, L.; Bleckmann, A.; Menck, K. Extracellular Vesicles in Liquid Biopsies as Biomarkers for Solid Tumors. Cancers 2023, 15, 1307. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.; Wildman, D.E. Extracellular Vesicles and the Promise of Continuous Liquid Biopsies. J. Pathol. Transl. Med. 2018, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kustanovich, A.; Schwartz, R.; Peretz, T.; Grinshpun, A. Life and Death of Circulating Cell-Free DNA. Cancer Biol. Ther. 2019, 20, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.W.; Charkhchi, P.; Akbari, M.R. Potential Clinical Utility of Liquid Biopsies in Ovarian Cancer. Mol. Cancer 2022, 21, 114. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, X.; Zhang, W.; Yu, L.; Wang, Y.; Deng, Z.; Liu, M.; Mo, S.; Wang, R.; Zhao, J.; et al. Trends in the Biological Functions and Medical Applications of Extracellular Vesicles and Analogues. Acta Pharm. Sin. B 2021, 11, 2114–2135. [Google Scholar] [CrossRef]
- Jurj, A.; Pop-Bica, C.; Slaby, O.; Ştefan, C.D.; Cho, W.C.; Korban, S.S.; Berindan-Neagoe, I. Tiny Actors in the Big Cellular World: Extracellular Vesicles Playing Critical Roles in Cancer. Int. J. Mol. Sci. 2020, 21, 7688. [Google Scholar] [CrossRef]
- Abhange, K.; Makler, A.; Wen, Y.; Ramnauth, N.; Mao, W.; Asghar, W.; Wan, Y. Small Extracellular Vesicles in Cancer. Bioact. Mater. 2021, 6, 3705–3743. [Google Scholar] [CrossRef]
- Yee, N.S.; Zhang, S.; He, H.Z.; Zheng, S.Y. Extracellular Vesicles as Potential Biomarkers for Early Detection and Diagnosis of Pancreatic Cancer. Biomedicines 2020, 8, 581. [Google Scholar] [CrossRef]
- Wang, X.; Tian, L.; Lu, J.; Ng, I.O.L. Exosomes and Cancer—Diagnostic and Prognostic Biomarkers and Therapeutic Vehicle. Oncogenesis 2022, 11, 54. [Google Scholar] [CrossRef]
- Furini, S.; Falciani, C. Expression and Role of Heparan Sulfated Proteoglycans in Pancreatic Cancer. Front. Oncol. 2021, 11, 695858. [Google Scholar] [CrossRef]
- Vicente, C.M.; da Silva, D.A.; Sartorio, P.V.; Silva, T.D.; Saad, S.S.; Nader, H.B.; Forones, N.M.; Toma, L. Heparan Sulfate Proteoglycans in Human Colorectal Cancer. Anal. Cell. Pathol. 2018, 2018, 8389595. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Huang, K.; Gao, C.; Lau, Q.C.; Pan, H.; Xie, K.; Li, J.; Liu, R.; Zhang, T.; Xie, N.; et al. Proteomics Identification of ITGB3 as a Key Regulator in Reactive Oxygen Species-Induced Migration and Invasion of Colorectal Cancer. Mol. Cell. Proteom. 2011, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Brennan, K.; Martin, K.; FitzGerald, S.P.; O’Sullivan, J.; Wu, Y.; Blanco, A.; Richardson, C.; Mc Gee, M.M. A Comparison of Methods for the Isolation and Separation of Extracellular Vesicles from Protein and Lipid Particles in Human Serum. Sci. Rep. 2020, 10, 1039. [Google Scholar] [CrossRef]
- Palviainen, M.; Saraswat, M.; Varga, Z.; Kitka, D.; Neuvonen, M.; Puhka, M.; Joenväärä, S.; Renkonen, R.; Nieuwland, R.; Takatalo, M.; et al. Extracellular Vesicles from Human Plasma and Serum Are Carriers of Extravesicular Cargo—Implications for Biomarker Discovery. PLoS ONE 2020, 15, e0236439. [Google Scholar] [CrossRef]
- Shushkova, N.A.; Vavilov, N.E.; Novikova, S.E.; Farafonova, T.E.; Tikhonova, O.V.; Liao, P.-C.; Zgoda, V.G. Quantitative Proteomics of Human Blood Exosomes. Biomed. Khim. 2018, 64, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Ondruššek, R.; Kvokačková, B.; Kryštofová, K.; Brychtová, S.; Souček, K.; Bouchal, J. Prognostic Value and Multifaceted Roles of Tetraspanin CD9 in Cancer. Front. Oncol. 2023, 13, 1140738. [Google Scholar] [CrossRef]
- Kim, K.J.; Kwon, H.J.; Kim, M.C.; Bae, Y.K. CD9 Expression in Colorectal Carcinomas and Its Prognostic Significance. J. Pathol. Transl. Med. 2016, 50, 459–468. [Google Scholar] [CrossRef]
- Koh, H.M.; Jang, B.G.; Lee, D.H.; Hyun, C.L. Increased CD9 Expression Predicts Favorable Prognosis in Human Cancers: A Systematic Review and Meta-Analysis. Cancer Cell Int. 2021, 21, 472. [Google Scholar] [CrossRef]
- Zeng, P.; Si, M.; Sun, R.X.; Cheng, X.; Li, X.Y.; Chen, M. Bin Prognostic Value of CD9 in Solid Tumor: A Systematic Review and Meta-Analysis. Front. Oncol. 2021, 11, 764630. [Google Scholar] [CrossRef]
- Kaprio, T.; Hagström, J.; Andersson, L.C.; Haglund, C. Tetraspanin CD63 Independently Predicts Poor Prognosis in Colorectal Cancer. Histol. Histopathol. 2020, 35, 887–892. [Google Scholar]
- Prodduturvar, P.; McCormick, B.; Mneimneh, W.; Zotto, V.; Escobar, D.; Manne, A.; Grimm, L.; Rider, P.; Hunter, J.; Iliff, G.; et al. Exosomal Marker CD63 Expression Pattern Using Immunohistochemistry (IHC) in Patients with Rectal Adenocarcinoma in Comparison with Left-Sided Colon Cancer. J. Clin. Oncol. 2020, 38, e16096. [Google Scholar] [CrossRef]
- Valdés-Mora, F.; Locke, W.J.; Bandrés, E.; Gallego-Ortega, D.; Cejas, P.; García-Cabezas, M.A.; Colino-Sanguino, Y.; Gómez, T.; Lacal, J.C. Clinical Relevance of the Transcriptional Signature Regulated by CDC42 in Colorectal Cancer. Oncotarget 2017, 8, 26755–26770. [Google Scholar] [CrossRef] [PubMed]
- Gómez, T.; Pulgar, D.E.L.; Valdés-Mora, F.; Bandrés, E.V.A.; Pérez-Palacios, R.; Espina, C.; Cejas, P.; García-Cabezas, M.A.; Nistal, M.; Casado, E.; et al. Cdc42 Is Highly Expressed in Colorectal Adenocarcinoma and Downregulates ID4 through an Epigenetic Mechanism. Int. J. Oncol. 2008, 33, 185–193. [Google Scholar]
- Pascal, L.E.; True, L.D.; Campbell, D.S.; Deutsch, E.W.; Risk, M.; Coleman, I.M.; Eichner, L.J.; Nelson, P.S.; Liu, A.Y. Correlation of MRNA and Protein Levels: Cell Type-Specific Gene Expression of Cluster Designation Antigens in the Prostate. BMC Genom. 2008, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.; Wu, G.; Zhang, W. Correlation of MRNA Expression and Protein Abundance Affected by Multiple Sequence Features Related to Translational Efficiency in Desulfovibrio Vulgaris: A Quantitative Analysis. Genetics 2006, 174, 2229–2243. [Google Scholar] [CrossRef] [PubMed]
- Vafaei, S.; Saeednejad Zanjani, L.; Habibi Shams, Z.; Naseri, M.; Fattahi, F.; Gheytanchi, E.; Alemrajabi, M.; Ebrahimi, M.; Madjd, Z. Low Expression of Talin1 Is Associated with Advanced Pathological Features in Colorectal Cancer Patients. Sci. Rep. 2020, 10, 17786. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. High Expression of HSPA8 Is A Favorable Prognostic Factor in Colon Cancer. Res. Sq. 2021, 1–19. [Google Scholar] [CrossRef]
- Suhovskih, A.V.; Aidagulova, S.V.; Kashuba, V.I.; Grigorieva, E.V. Proteoglycans as Potential Microenvironmental Biomarkers for Colon Cancer. Cell Tissue Res. 2015, 361, 833–844. [Google Scholar] [CrossRef]
- Novikova, S.E.; Soloveva, N.A.; Farafonova, T.E.; Tikhonova, O.V.; Liao, P.-C.; Zgoda, V.G. Proteomic Signature of Extracellular Vesicles for Lung Cancer Recognition. Molecules 2021, 26, 6145. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CRC | HVs | |
|---|---|---|
| Total | 11 | 20 |
| Age | 50–70 | 45–74 |
| Male | 7 | 10 |
| Female | 4 | 10 |
| Stage 3, 3A | 4 | - |
| Stage 3B, 3C | 5 | - |
| Stage 4 | 2 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soloveva, N.; Novikova, S.; Farafonova, T.; Tikhonova, O.; Zgoda, V. Proteomic Signature of Extracellular Vesicles Associated with Colorectal Cancer. Molecules 2023, 28, 4227. https://doi.org/10.3390/molecules28104227
Soloveva N, Novikova S, Farafonova T, Tikhonova O, Zgoda V. Proteomic Signature of Extracellular Vesicles Associated with Colorectal Cancer. Molecules. 2023; 28(10):4227. https://doi.org/10.3390/molecules28104227
Chicago/Turabian StyleSoloveva, Natalia, Svetlana Novikova, Tatiana Farafonova, Olga Tikhonova, and Victor Zgoda. 2023. "Proteomic Signature of Extracellular Vesicles Associated with Colorectal Cancer" Molecules 28, no. 10: 4227. https://doi.org/10.3390/molecules28104227
APA StyleSoloveva, N., Novikova, S., Farafonova, T., Tikhonova, O., & Zgoda, V. (2023). Proteomic Signature of Extracellular Vesicles Associated with Colorectal Cancer. Molecules, 28(10), 4227. https://doi.org/10.3390/molecules28104227