
A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids


Abstract
:1. Introduction
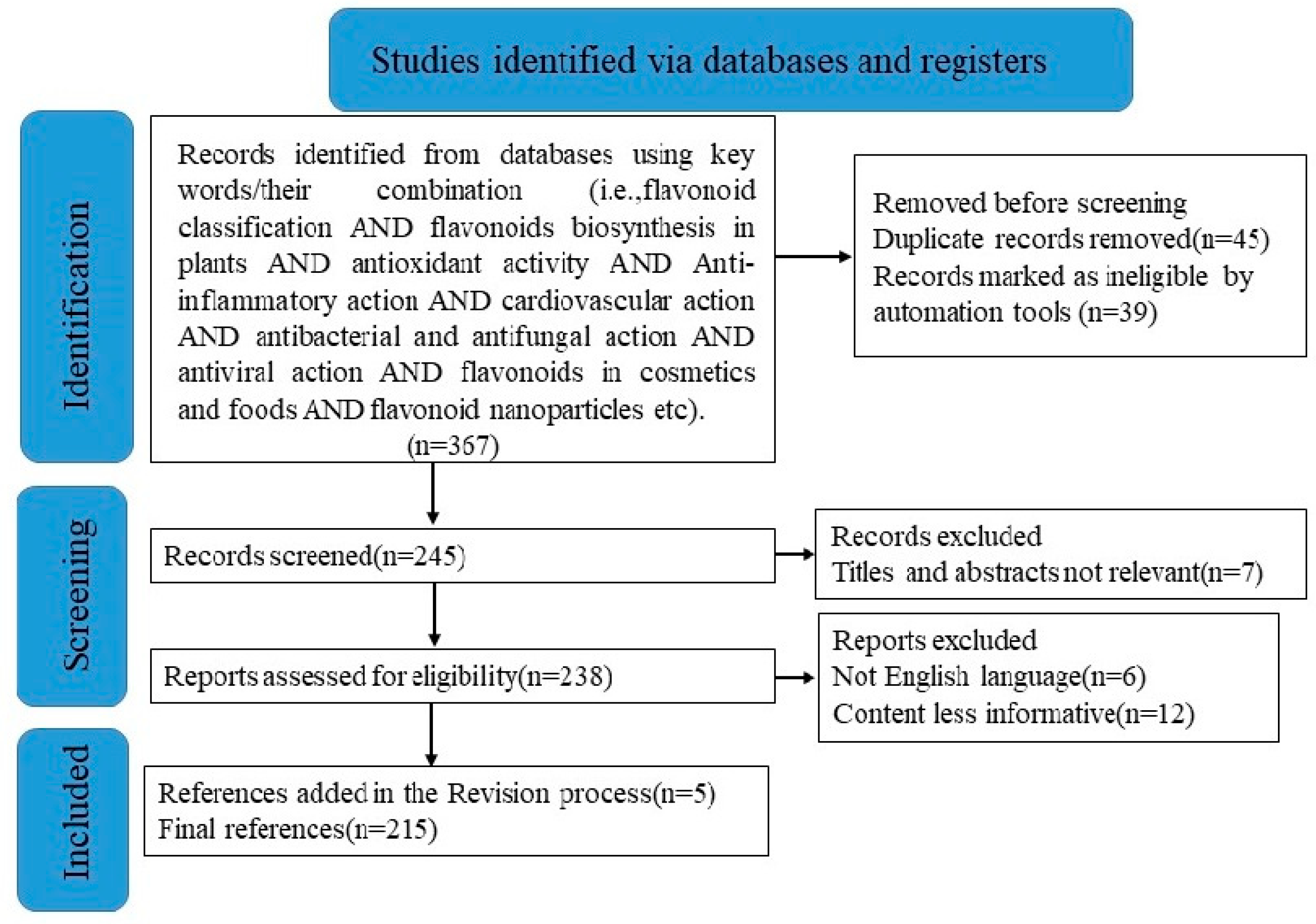
2. Methodology
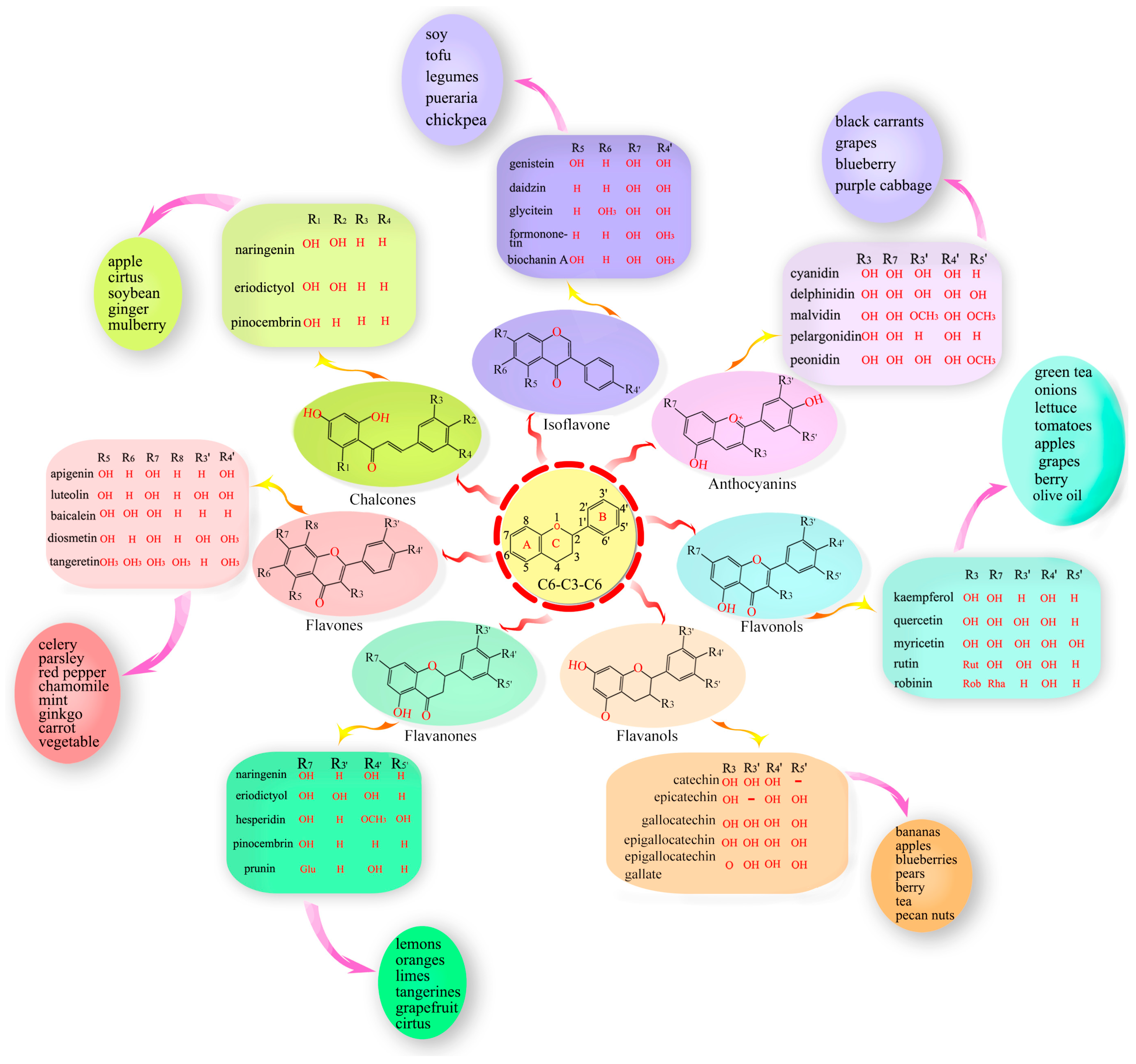
3. Flavonoid Classification
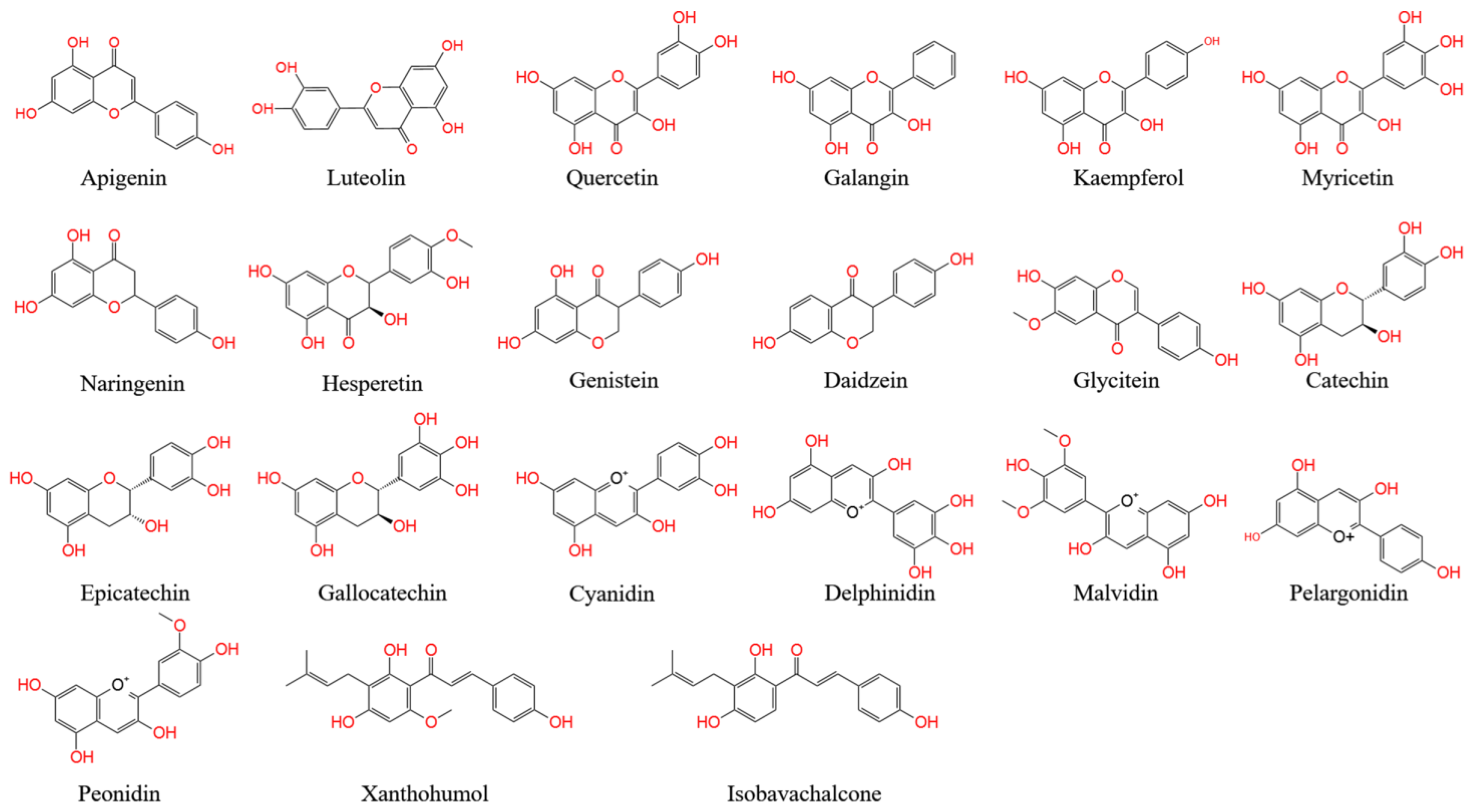
3.1. Flavones
3.2. Flavonols
3.3. Flavanones
3.4. Isoflavonoids
3.5. Flavanols
3.6. Anthocyanins
3.7. Chalcones
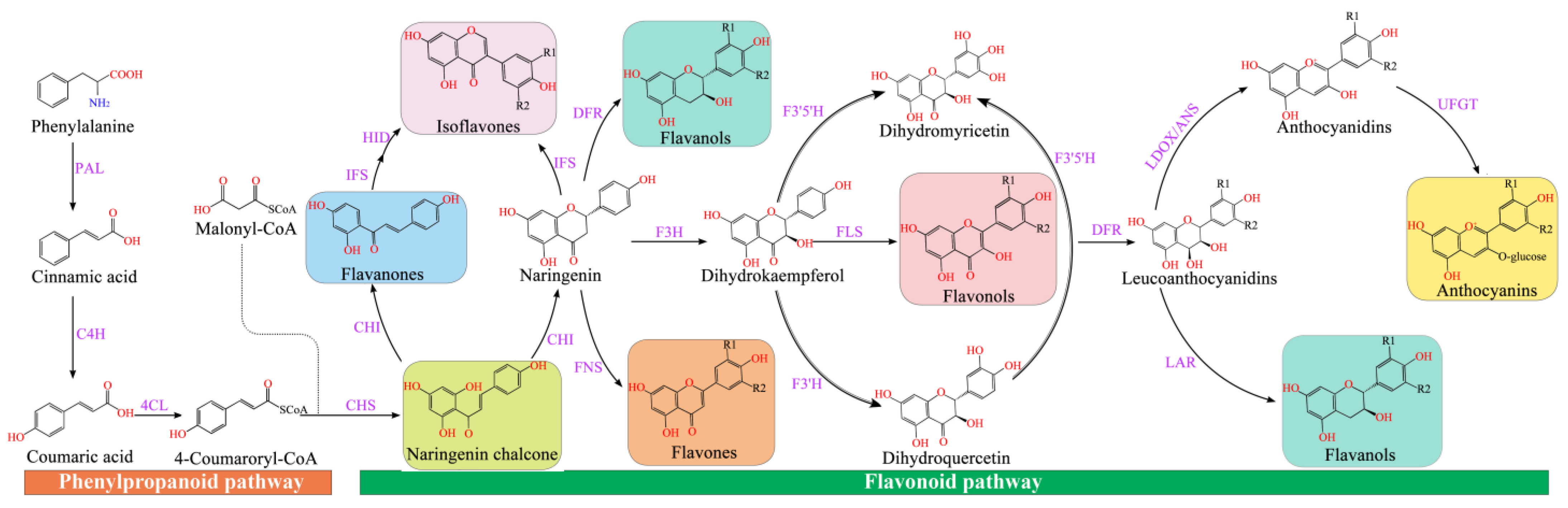
4. Flavonoid Biosynthesis in Plants
4.1. Flavonoid Biosynthetic Pathways
4.2. Transcriptional Regulation of Flavonoid Synthesis
4.3. Non-Coding RNA Regulates Flavonoid Biosynthesis
5. Pharmacological Properties of Flavonoids
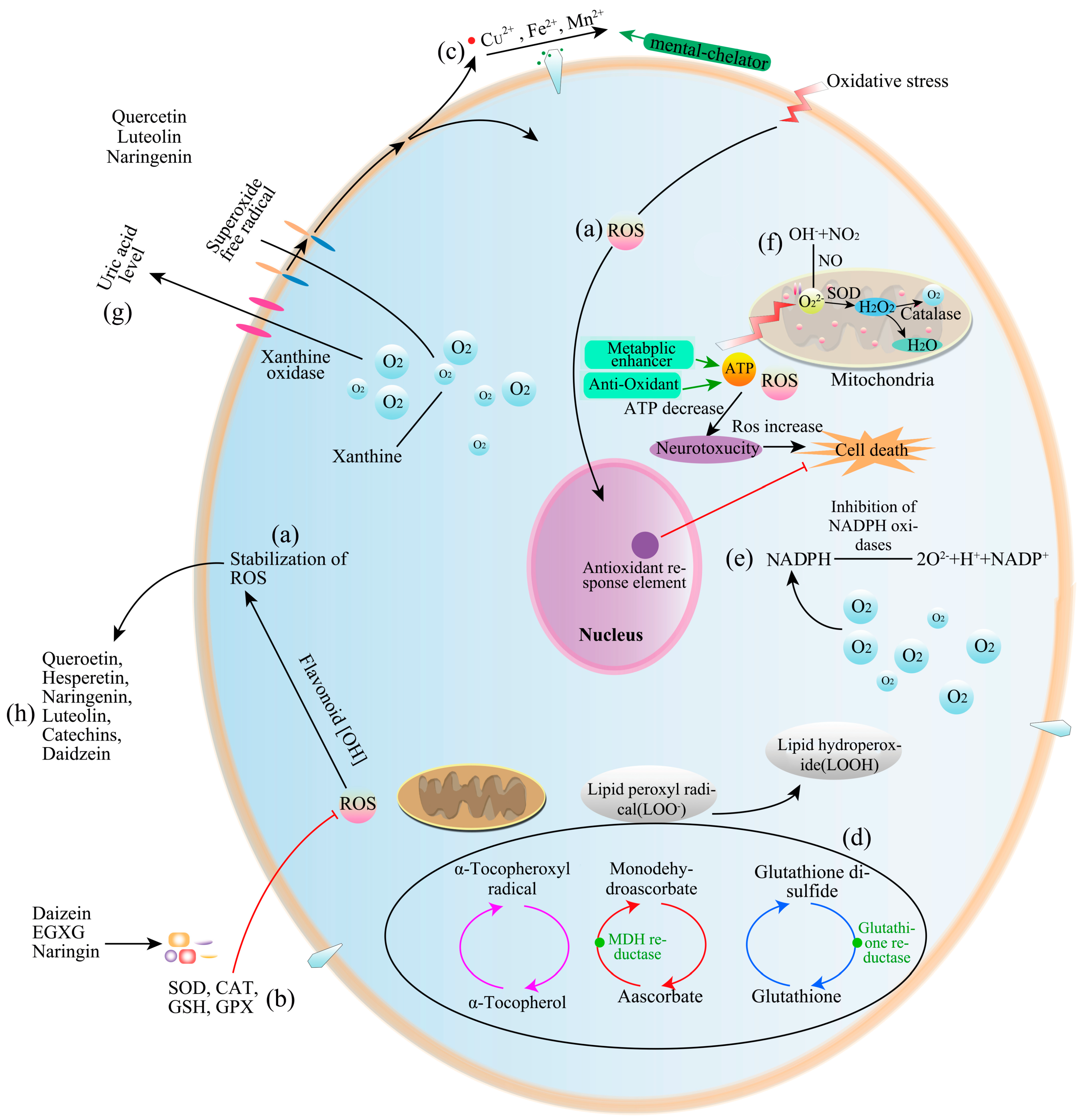
5.1. Antioxidant Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flavonoids | Classification | Pharmacological Activity | Sources of Plant | References |
|---|---|---|---|---|
| Proanthocyanidins | anthocyanins | antioxidant, anti-inflammatory, antibacterial, antifungal and anti-cardiovascular | grapes, apples, sorghum, cherries, and other natural plant | [93] |
| Cyanidin | anthocyanins | anti-inflammatory, antiviral, and anticancer | black rice, black beans, purple potatoes, blueberries | [108] |
| Curcumin | curcuminoids | anti-inflammatory and anticancer | Curcuma longa | [109] |
| Methyl chalcone | chalcones | anti-inflammatory and anticancer | apple, citrus, soybean, ginger, mulberry | [110] |
| Trans-chalcone | chalcones | anti-inflammatory and anticancer | apple, citrus, soybean, ginger, mulberry | [110] |
| Xanthohumol | chalcones | anti-cardiovascular and antiviral | Humulus lupulus | [111] |
| Licochalcone | chalcones | antibacterial and antifungal | Glycyrrhiza uralensis | [112] |
| Catechin | flavanols | antioxidant, anti-inflammatory, antiviral, and anti-cardiovascular | Camellia sinensis | [101,102,113] |
| Epigallocatechin gallate | flavanols | antioxidant, antibacterial, antifungal, anti-cardiovascular, and antiviral | Camellia sinensis | [104,114,115] |
| Naringin | flavanones | antioxidant, anti-inflammatory, anti-cardiovascular, and antiviral | lemons, oranges, grapefruits, citrus | [104,110,116,117,118,119] |
| Hesperidin | flavanones | anti-inflammatory, anti-cardiovascular, and antiviral | lemons, limes, oranges, grapefruits, citrus | [116,117,120,121] |
| Diosmin | flavanones | anti-inflammatory | citrus fruits | [122] |
| Orientin | flavanones | anti-inflammatory | Trollius chinensis, Cajanus cajan, Crataegus laevigata | [123] |
| Vitexin | flavanones | antioxidant, anti-inflammatory, and anticancer | Ficus deltoid, Spirodela polyrhiza | [123] |
| Acacetin | flavanones | anti-cardiovascular, anticancer, and antiviral | Acacia farnesiana | [124,125] |
| Silymarin | flavanones | antioxidant, anti-cardiovascular, and antiviral | Silybum marianum | [126,127] |
| Liquiritigenin | flavanones | anti-inflammatory, antiviral, and anticancer | Glycyrrhiza uralensis | [128] |
| Isorhamnetin | flavanones | antiviral and anticancer | Ginkgo biloba, Hippophae rhamnoides | [125] |
| Apigenin | flavones | antibacterial, antifungal, and antiviral | Apium graveolens | [129,130,131,132] |
| Morin | flavones | antioxidant and anti-inflammatory | Cudrania cochinchinensis, Maclura pomifera | [133] |
| Baicalin | flavones | Anti-cardiovascular, antibacterial, and antifungal | Scutellaria baicalensis | [114,134] |
| Luteolin | flavones | anti-inflammatory, anti-cardiovascular, and antiviral | Dracocephalum integrifolium, Lonicera japonica, Capsicum annuum | [132,135] |
| Fisetin | flavonols | antioxidant | strawberry, apple, onion, cucumber, and other fruits and vegetables | [96] |
| Quercetin | flavonols | antioxidant, anti-inflammatory, anti-cardiovascular, antibacterial, and antifungal | vegetables, fruit, seeds, nuts, tea, and red wine | [100,102,107,120,136,137,138] |
| Rutin | flavonols | antioxidant, anti-inflammatory, and antiviral | rue, tobacco, jujube, apricot, orange, tomato, buckwheat, and citrus fruits | [101,120,126,127] |
| Kaempferol | flavonols | antioxidant, anti-inflammatory, antibacterial, antiviral, and anticancer | fruits, vegetables, herbs, and other natural plants | [101,133,139] |
| Myricetin | flavonols | antioxidant, anti-inflammatory, and anti-cardiovascular | Myrica rubra | [133,140,141] |
| Glabrol | isoflavane | antibacterial and antifungal | Glycyrrhiza uralensis | [112] |
| Genistein | isoflavone | antioxidant, antifungal, antiviral, and anticancer | soybeans and other plants | [120,142,143] |
5.2. Anti-Inflammatory Action
5.3. Cardiovascular Action
5.4. Antibacterial and Antifungal Action
5.5. Antiviral Action
6. Applications of Flavonoids in Cosmetics and Foods
6.1. Applications of Flavonoids in Cosmetics
6.2. Application of Flavonoids in Foods
7. Omics Research and Flavonoid Nanoparticles
8. Conclusions
9. Future Perspectives
9.1. Mining of Functional Genes
9.2. Extraction and Utilization of Bioactive Ingredients
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


| Flavonoids | Biological Activity | Mechanism of Action | References |
|---|---|---|---|
| Proanthocyanidins | antioxidant activity | polymerizing flavonoid monomer and Catechin polymer | [93] |
| Fisetin | antioxidant activity | inhibiting the activities of oxidase | [10,96,97] |
| Hydroxyflavones | antioxidant activity | generating complexes with metal cations | [98] |
| Kaempferol | antioxidant activity | chelate Cu(II) ions | [101] |
| Catechol | antioxidant activity | generating stable metallic complexes | [102] |
| Quercetin | antioxidant activity | interacting with α-tocopherol radicals | [104,106,107] |
| Epigallocatechin Gallate Naringin | antioxidant activity | interacting with α-tocopherol radicals | [104] |
| Apigenin | antiinflammatory action | reducing the steady-state mRNA levels induced by TNF-α | [145] |
| Quercetin catechins | antiinflammatory action | reducing the expression of pro-inflammatory cytokines | [113,133] |
| Kaempferol Myricetin | antiinflammatory action | inhibit the activity of lipoxygenase | [133] |
| Diosmin Hesperidin | antiinflammatory action | inhibiting leukotriene B4 biosynthesis | [122] |
| Chalcones | antiinflammatory action | inhibiting proinflammatory cytokines activity | [110] |
| Vitexin Orientin Rutin | antiinflammatory action | inhibiting iNOS and COX2 expression | [123] |
| Naringenin Hesperetin | anticardiovascular action | reducing high blood pressure by promoting vasodilation | [116,117] |
| Quercetin | anticardiovascular action | protecting the heart against ischemia-reperfusion injury via the cardioprotective effects of quercetin. | [116,117,137] |
| Isoflavones | anticardiovascular action | inhibiting monocyte-endothelial cell adhesion | [153] |
| Anthocyanins | anticardiovascular action | improving systolic blood pressure, and reduce the content of triglycerides, total cholesterol, and low-density lipoprotein cholesterol | [154,155] |
| Xanthohumol | anticardiovascular action | increasing PTEN expression and inhibited AKT/mTOR phosphorylation | [111] |
| Flavonoid | anticardiovascular action | modulating the PI3K/Akt/GSK3β pathway or downregulating the ROS-mediated JNK/p38MAPK/NFκB pathway | [141,156] |
| the citrus flavonoid naringenin | anticardiovascular action | enhancing SIRT1 expression, reduced ROS production | [118] |
| Quercetin | antibacterial action | blocking the biofilm formation and suppressed the DNA replication | [8,138,158,166] |
| Apigenin Naringenin Chrysin Genistein Kaempferol Daidzin | Antibacterial action | blocking the biofilm formation or regulate ROS species, decrease lipid peroxidation | [8,129,130,158,166] |
| Baicalein Luteolin Myricetin | antibacterial action | suppressing the DNA replication or suppressed the biosynthesis of ATP | [8,129,130,158] |
| Epigallocatechin gallate | antibacterial action | suppressing the biosynthesis of ATP | [114] |
| Glabrol | antibacterial action | increasing the permeability of the cell membrane and collapsing the proton motive force | [112] |
| Glabridin | antifungal action | inhibiting the biosynthesis of the main components of fungi cell walls | [114] |
| Myricetin Kaempferol Naringenin Genistein Luteolin | antifungal action | suppressing the biosynthesis of DNA, RNA, and protein | [166] |
| Glcyrrhiza Glabra Isoflavones Chalcone | antifungal action | regulating the expression level of phosphatidylserine decarboxylase | [167] |
| Apigenin | antiviral action | inhibiting viruses from entering the host cells, modulate the immune system, and reduce viral load | [131,169] |
| Baicalein | antiviral action | blocking the replication of the avian influenza H5N1 virus | [170] |
| Luteolin | antiviral action | inhibiting HIV-1 reactivation | [135] |
| Kaempferol | antiviral action | blocking the HIV-1 replication | [139] |
| Liquiritigenin | promoted apoptosis | increasing the p53 and Bax gene expression, decreasing Bcl-2 gene expression | [128] |
| Isoflflavone genistein | suppress the proliferation | promoting breast cancer cell arrest at the G2/M phase and subsequent ROS-dependent apoptosis | [125,143] |
| Kaempferol | suppress the proliferation | inducing apoptosis via cell cycle arrest MDA-MB-453 cells | [125,142] |
| Naringenin | inhibit the proliferation and migration | inhibiting ROS formation | [119] |
| Apigenin Luteolin | induce apoptosis | causing the alterations to ROS signaling | [132] |
References
- Santos, E.L.; Maia, B.; Ferriani, A.P.; Teixeira, S.D. Flavonoids: Classification, biosynthesis and chemical ecology. In Flavonoids: From Biosynthesis to Human Health; IntechOpen: London, UK, 2017; Volume 13, pp. 78–94. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.-H.; Jaremko, M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef]
- García-Lafuente, A.; Guillamón, E.; Villares, A.; Rostagno, M.A.; Martínez, J.A. Flavonoids as anti-inflammatory agents: Implications in cancer and cardiovascular disease. Inflamm. Res. 2009, 58, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Karak, P. Biological activities of flavonoids: An overview. Int. J. Pharm. Sci. Res. 2019, 10, 1567–1574. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- do Nascimento, R.P.; dos Santos, B.L.; Amparo, J.A.O.; Soares, J.R.P.; da Silva, K.C.; Santana, M.R.; Almeida, Á.M.A.N.; da Silva, V.D.A.; Costa, M.d.F.D.; Ulrich, H.; et al. Neuroimmunomodulatory Properties of Flavonoids and Derivates: A Potential Action as Adjuvants for the Treatment of Glioblastoma. Pharmaceutics 2022, 14, 116. [Google Scholar] [CrossRef] [PubMed]
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr. Plant. Biol. 2010, 52, 98–111. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S. Plant flavonoids: Chemical characteristics and biological activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Vicente, O.; Boscaiu, M. Flavonoids: Antioxidant compounds for plant defence and for a healthy human diet. Not. Bot. Horti Agrobot. Cluj. 2018, 46, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, W.-B.; Li, Y.-H.; Shu, X.-C.; Pu, Y.-T.; Wang, X.-J.; Wang, T.; Wang, Z. The Classification, Molecular Structure and Biological Biosynthesis of Flavonoids, and Their Roles in Biotic and Abiotic Stresses. Molecules 2023, 28, 3599. [Google Scholar] [CrossRef]
- Pietta, P.; Minoggio, M.; Bramati, L. Plant polyphenols: Structure, occurrence and bioactivity. Stud. Nat. Prod. Chem. 2003, 28, 257–312. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Lubna Kim, K.M. Plant secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Sircaik, S.; Dhiman, K.; Gambhir, G.; Kumar, P.; Srivastava, D.K. Transgenic Implications for Biotic and Abiotic Stress Tolerance in Agricultural Crops. In Agricultural Biotechnology: Latest Research and Trends; Springer: Berlin/Heidelberg, Germany, 2021; pp. 185–221. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef]
- Khuntia, A.; Martorell, M.; Ilango, K.; Bungau, S.G.; Radu, A.F.; Behl, T.; Sharifi-Rad, J. Theoretical evaluation of Cleome species’ bioactive compounds and therapeutic potential: A literature review. Biomed. Pharmacother. 2022, 151, 113161. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Murtaza, G.; Liu, G.; Yin, Y. Flavonoids and type 2 diabetes: Evidence of efficacy in clinical and animal studies and delivery strategies to enhance their therapeutic efficacy. Pharm. Res. 2020, 152, 104629. [Google Scholar] [CrossRef] [PubMed]
- Rehan, M. Biosynthesis of Diverse Class Flavonoids via Shikimate and Phenylpropanoid Pathway. In Bioactive Compounds-Biosynthesis, Characterization and Applications; IntechOpen: London, UK, 2021; p. 75392. [Google Scholar]
- Zakaryan, H.; Arabyan, E.; Oo, A.; Zandi, K. Flavonoids: Promising natural compounds against viral infections. Arch. Virol. 2017, 162, 2539–2551. [Google Scholar] [CrossRef] [PubMed]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. 2020, 60, 626–659. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Sarkar, D.; Bose, A.; Mandal, S.C. Natural flavonoids and its pharmaceutical importance. Pharma Rev. 2018, 94, 61–75. [Google Scholar]
- Tang, D.; Chen, K.; Huang, L.; Li, J. Pharmacokinetic properties and drug interactions of apigenin, a natural flavone. Expert. Opin. Drug Metab. Toxi. 2017, 13, 323–330. [Google Scholar] [CrossRef]
- Ginwala, R.; Bhavsar, R.; Chigbu, D.G.I.; Jain, P.; Khan, Z.K. Potential role of flavonoids in treating chronic inflammatory diseases with a special focus on the anti-inflammatory activity of apigenin. Antioxidants 2019, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Proteggente, A.R.; Pannala, A.S.; Paganga, G.; Buren, L.V.; Wagner, E.; Wiseman, S.; Van De Put, F.; Dacombe, C.; Rice-Evans, C.A. The antioxidant activity of regularly consumed fruit and vegetables reflects their phenolic and vitamin C composition. Free Radic. Res. 2002, 36, 217–233. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Brillante, L.; Kurtural, S.K. Flavonol profile is a reliable indicator to assess canopy architecture and the exposure of red wine grapes to solar radiation. Front. Plant. Sci. 2019, 10, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Parisi, A.; Normanno, G. Polyphenols as emerging antimicrobial agents. In Emerging Modalities in Mitigation of Antimicrobial Resistance; Springer International Publishing: Cham, Switzerland, 2022; pp. 219–259. [Google Scholar] [CrossRef]
- Vandercook, C.E.; Stephenson, R.G. Lemon juice composition. Identification of major phenolic compounds and estimation by paper chromatography. J. Agr. Food Chem. 1966, 14, 450–454. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.J.; Chen, J.B.; Cao, J.P.; Sun, C.D. Citrus flavonoids and their antioxidant evaluation. Crit. Rev. Food Sci. 2022, 62, 3833–3854. [Google Scholar] [CrossRef] [PubMed]
- Najmanová, I.; Vopršalová, M.; Saso, L.; Mladěnka, P. The pharmacokinetics of flavanones. Crit. Rev. Food Sci. 2020, 60, 3155–3171. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Datta, A. Fundamentals of phytochemicals. In Cancer Genetics and Therapeutics; Springer: Singapore, 2019; pp. 49–81. [Google Scholar] [CrossRef]
- Galleano, M.; Calabro, V.; Prince, P.D.; Litterio, M.C.; Piotrkowski, B.; Vazquez-Prieto, M.A.; Miatello, R.M.; Oteiza, P.I.; Fraga, C.G. Flavonoids and metabolic syndrome. Ann. Acad. Sci. 2012, 1259, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qian, J.; Li, J.; Xing, M.; Grierson, D.; Sun, C.; Xu, C.; Li, X.; Chen, K. Hydroxylation decoration patterns of flavonoids in horticultural crops: Chemistry, bioactivity, and biosynthesis. Hortic. Res. 2022, 9, uhab068. [Google Scholar] [CrossRef]
- Shah, A.; Smith, D.L. Flavonoids in agriculture: Chemistry and roles in, biotic and abiotic stress responses, and microbial associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- Kopečná-Zapletalová, M.; Krasulová, K.; Anzenbacher, P.; Hodek, P.; Anzenbacherová, E. Interaction of isoflavonoids with human liver microsomal cytochromes P450: Inhibition of CYP enzyme activities. Xenobiotica 2017, 47, 324–331. [Google Scholar] [CrossRef]
- Hummelova, J.; Rondevaldova, J.; Balastikova, A.; Lapcik, O.; Kokoska, L. The relationship between structure and in vitro antibacterial activity of selected isoflavones and their metabolites with special focus on antistaphylococcal effect of demethyltexasin. Lett. Appl. Microbiol. 2015, 60, 242–247. [Google Scholar] [CrossRef]
- Lim, Y.J.; Jeong, H.Y.; Gil, C.S.; Kwon, S.J.; Na, J.K.; Lee, C.; Eom, S.H. Isoflavone accumulation and the metabolic gene expression in response to persistent UV-B irradiation in soybean sprouts. Food Chem. 2020, 303, 125376. [Google Scholar] [CrossRef]
- Meng, N.; Yu, B.J.; Guo, J.S. Ameliorative effects of inoculation with Bradyrhizobium japonicum on Glycine max and Glycine soja seedlings under salt stress. Plant. Growth Regul. 2016, 80, 137–147. [Google Scholar] [CrossRef]
- Mori-Yasumoto, K.; Hashimoto, Y.; Agatsuma, Y.; Fuchino, H.; Yasumoto, K.; Shirota, O.; Satake, M.; Sekita, S. Leishmanicidal phenolic compounds derived from Dalbergia cultrata. Nat. Prod. Res. 2021, 35, 4907–4915. [Google Scholar] [CrossRef]
- Dutta, M.S.; Mahapatra, P.; Ghosh, A.; Basu, S. Estimation of the reducing power and electrochemical behavior of few flavonoids and polyhydroxybenzophenones substantiated by bond dissociation energy: A comparative analysis. Mol. Divers. 2022, 26, 1101–1113. [Google Scholar] [CrossRef] [PubMed]
- Pico, J.; Xu, K.; Guo, M.; Mohamedshah, Z.; Ferruzzi, M.G.; Martinez, M.M. Manufacturing the ultimate green banana flour: Impact of drying and extrusion on phenolic profile and starch bioaccessibility. Food Chem. 2019, 297, 124990. [Google Scholar] [CrossRef]
- Ding, T.; Cao, K.; Fang, W.; Zhu, G.; Chen, C.; Wang, X.; Wang, L. Evaluation of phenolic components (anthocyanins, flavanols, phenolic acids, and flavonols) and their antioxidant properties of peach fruits. Sci. Hortic. 2020, 268, 109365. [Google Scholar] [CrossRef]
- Yang, S.; Mi, L.; Wu, J.; Liao, X.; Xu, Z. Strategy for anthocyanins production: From efficient green extraction to novel microbial biosynthesis. Crit. Rev. Food Sci. 2022, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Guven, H.; Arici, A.; Simsek, O. Flavonoids in our foods: A short review. J. Basic. Clin. Health Sci. 2019, 3, 96–106. [Google Scholar] [CrossRef]
- Di Pietro, N.; Baldassarre, M.P.A.; Cichelli, A.; Pandolfi, A.; Formoso, G.; Pipino, C. Role of polyphenols and carotenoids in endothelial dysfunction: An overview from classic to innovative biomarkers. Oxid. Med. Cell. Longev. 2020, 2020, 6381380. [Google Scholar] [CrossRef]
- Fang, J. Classification of fruits based on anthocyanin types and relevance to their health effects. Nutrition 2015, 31, 1301–1306. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Chaves-Silva, S.; Dos Santos, A.L.; Chalfun-Júnior, A.; Peres, L.E.P.; Zhao, J.; Benedito, V.A. Understanding the genetic regulation of anthocyanin biosynthesis in plants–tools for breeding purple varieties of fruits and vegetables. Phytochemistry 2018, 153, 11–27. [Google Scholar] [CrossRef]
- Tohge, T.; de Souza, L.P.; Fernie, A.R. Current understanding of the pathways of flavonoid biosynthesis in model and crop plants. J. Exp. Bot. 2017, 68, 4013–4028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.I.; Hidalgo-Shrestha, C.; Bonawitz, N.D.; Franke, R.B.; Clint, C. Spatio-temporal control of phenylpropanoid biosynthesis by inducible complementation of a cinnamate 4-hydroxylase mutant. J. Exp. Bot. 2021, 72, 3061–3073. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Gonzali, S.; Perata, P. Anthocyanins from purple tomatoes as novel antioxidants to promote human health. Antioxidants 2020, 9, 1017. [Google Scholar] [CrossRef]
- Goyal, K.; Kaur, R.; Goyal, A.; Awasthi, R. Chalcones: A review on synthesis and pharmacological activities. J. Appl. Pharm. Sci. 2021, 11, 001–014. [Google Scholar] [CrossRef]
- Jasim, H.A.; Nahar, L.; Jasim, M.A.; Moore, S.A.; Ritchie, K.J.; Sarker, S.D. Chalcones: Synthetic chemistry follows where nature leads. Biomolecules 2021, 11, 1203. [Google Scholar] [CrossRef]
- Li, M.; Guo, L.; Wang, Y.; Li, Y.; Jiang, X.; Liu, Y.; Xie, D.; Gao, L.; Xia, T. Molecular and biochemical characterization of two 4-coumarate: CoA ligase genes in tea plant (Camellia sinensis). Plant Mol. Biol. 2022, 109, 579–593. [Google Scholar] [CrossRef]
- Sun, T.; Li, S.; Song, X.; Pei, G.; Diao, J.; Cui, J.; Shi, M.; Chen, L.; Zhang, W. Re-direction of carbon flux to key precursor malonyl-CoA via artificial small RNAs in photosynthetic Synechocystis sp. PCC 6803. Biotechnol. Biofuels. 2018, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Liu, X.; Gong, Q.; Cao, J.; Shen, W.; Yin, X.; Grierson, D.; Zhang, B.; Xu, C.; Li, X.; et al. Three AP2/ERF family members modulate flavonoid synthesis by regulating type IV chalcone isomerase in citrus. Plant Biotechnol. J. 2021, 19, 671–688. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.X.; Han, X.J.; Wu, Y.F.; Lou, H.X. The function and catalysis of 2-oxoglutarate-dependent oxygenases involved in plant flavonoid biosynthesis. Int. J. Mol. Sci. 2014, 15, 1080–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dastmalchi, M.; Dhaubhadel, S. Soybean chalcone isomerase: Evolution of the fold, and the differential expression and localization of the gene family. Planta 2015, 241, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.S. Tansley insight. New Phytol. 2022, 236, 2037–2043. [Google Scholar] [CrossRef]
- Baba, S.A.; Ashraf, N. Functional characterization of flavonoid 3′-hydroxylase, CsF3′ H, from Crocus sativus L: Insights into substrate specificity and role in abiotic stress. Arch. Biochem. Biophys. 2019, 667, 70–78. [Google Scholar] [CrossRef]
- Li, H.; Tian, J.; Yao, Y.; Zhang, J.; Song, T.; Li, K.; Yao, Y. Identification of leucoanthocyanidin reductase and anthocyanidin reductase genes involved in proanthocyanidin biosynthesis in Malus crabapple plants. Plant Physiol. Biochem. 2019, 139, 141–151. [Google Scholar] [CrossRef]
- Ma, S.; Hu, R.; Ma, J.; Fan, J.; Wu, F.; Wang, Y.; Huang, L.; Feng, G.; Li, D.; Nie, G.; et al. Integrative analysis of the metabolome and transcriptome provides insights into the mechanisms of anthocyanins and proanthocyanidins biosynthesis in Trifolium repens. Ind. Crop. Prod. 2022, 187, 115529. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Haq, I.U.; Pate, S.; Pan, X.; Naz, S.; Silva, A.S.; Saeed, F.; Suleria, H.A.R. Proanthocyanidins: A comprehensive review. Biomed. Pharmacother. 2019, 116, 108999. [Google Scholar] [CrossRef]
- Ni, J.; Zhao, Y.; Tao, R.; Yin, L.; Gao, L.; Strid, Å.; Qian, M.; Li, J.; Li, Y.; Shen, J.; et al. Ethylene mediates the branching of the jasmonate-induced flavonoid biosynthesis pathway by suppressing anthocyanin biosynthesis in red Chinese pear fruits. Plant Biotechnol. J. 2020, 18, 1223–1240. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Paramanantham, A.; Lee, W.S.; Yun, J.W.; Chang, S.H.; Kim, D.C.; Park, H.S.; Choi, Y.H.; Kim, G.S.; Ryu, C.H.; et al. Anthocyanins Derived from Vitis coignetiae Pulliat Contributes Anti-Cancer Effects by Suppressing NF-κB Pathways in Hep3B Human Hepatocellular Carcinoma Cells and In Vivo. Molecules 2020, 25, 5445. [Google Scholar] [CrossRef]
- Fenn, M.A.; Giovannoni, J.J. Phytohormones in fruit development and maturation. Plant J. 2021, 105, 446–458. [Google Scholar] [CrossRef]
- Corso, M.; Perreau, F.; Mouille, G.; Lepiniec, L. Specialized phenolic compounds in seeds: Structures, functions, and regulations. Plant Sci. 2020, 296, 110471. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Xing, C.; Yao, Z.; Huang, X. Pbr MYB 21, a novel MYB protein of Pyrus betulaefolia, functions in drought tolerance and modulates polyamine levels by regulating arginine decarboxylase gene. Plant Biotechnol. J. 2017, 15, 1186–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, W.; Xie, L.; Li, Y.; Liu, H.; Fu, X.; Chen, T.; Hassani, D.; Li, L.; Sun, X.; Tang, K. An R2R3-MYB transcription factor positively regulates the glandular secretory trichome initiation in Artemisia annua L. Front. Plant Sci. 2021, 12, 657156. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.; Yu, W.; Yao, H.; Zhou, P.; Allan, A.C.; Zeng, L. NtMYB3, an R2R3-MYB from narcissus, regulates flavonoid biosynthesis. Int. J. Mol. Sci. 2019, 20, 5456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Ning, Y.; Zhang, W.; Liao, Y.; Li, L.; Cheng, H.; Cheng, S. An R2R3-MYB transcription factor as a negative regulator of the flavonoid biosynthesis pathway in Ginkgo biloba. Funct. Integr. Genom. 2014, 14, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, B.; Zhong, Y.; Yao, L.; Cheng, L.; Wu, T. The soybean R2R3 MYB transcription factor GmMYB100 negatively regulates plant flavonoid biosynthesis. Plant Mol. Biol. 2015, 89, 35–48. [Google Scholar] [CrossRef]
- Premathilake, A.T.; Ni, J.; Bai, S.; Tao, R.; Teng, Y. R2R3-MYB transcription factor PpMYB17 positively regulates flavonoid biosynthesis in pear fruit. Planta 2020, 252, 1–16. [Google Scholar] [CrossRef]
- Sun, Z.; Linghu, B.; Hou, S.; Liu, R.; Wang, L.; Hao, Y.; Han, Y.; Zhou, M.; Liu, L.; Li, H. Tartary buckwheat FtMYB31 gene encoding an R2R3-MYB transcription factor enhances flavonoid accumulation in Tobacco. J. Plant Growth Regul. 2020, 39, 564–574. [Google Scholar] [CrossRef]
- Cultrone, A.; Cotroneo, P.S.; Reforgiato Recupero, G. Cloning and molecular characterization of R2R3-MYB and bHLH-MYC transcription factors from Citrus sinensis. Tree Genet. Genomes 2010, 6, 101–112. [Google Scholar] [CrossRef]
- Xie, X.B.; Li, S.; Zhang, R.F.; Zhao, J.; Chen, Y.C.; Zhao, Q.; Yao, Y.X.; You, C.X.; Zhang, X.S.; Hao, Y.J. The bHLH transcription factor MdbHLH3 promotes anthocyanin accumulation and fruit colouration in response to low temperature in apples. Plant Cell Environ. 2012, 35, 1884–1897. [Google Scholar] [CrossRef]
- Qi, Y.; Zhou, L.; Han, L.; Zou, H.; Wang, Y. PsbHLH1, a novel transcription factor involved in regulating anthocyanin biosynthesis in tree peony (Paeonia suffruticosa). Plant Physiol. Biochem. 2020, 154, 396–408. [Google Scholar] [CrossRef]
- Mano, H.; Ogasawara, F.; Sato, K.; Higo, H.; Minobe, Y. Isolation of a regulatory gene of anthocyanin biosynthesis in tuberous roots of purple-fleshed sweet potato. Plant Physiol. 2007, 143, 1252–1268. [Google Scholar] [CrossRef] [Green Version]
- Peniche-Pavía, H.A.; Guzmán, T.J.; Magaña-Cerino, J.M.; Gurrola-Díaz, C.M.; Tiessen, A. Maize Flavonoid Biosynthesis, Regulation, and Human Health Relevance: A Review. Molecules 2022, 27, 5166. [Google Scholar] [CrossRef]
- Qian, Y.; Zhang, T.; Yu, Y.; Gou, L.; Pi, E. Regulatory mechanisms of bHLH transcription factors in plant adaptive responses to various abiotic stresses. Front. Plant Sci. 2021, 12, 1143. [Google Scholar] [CrossRef]
- Qiu, Z.; Wang, H.; Li, D.; Byu, B.; Cao, B. Identification of Candidate HY5-Dependent and-Independent Regulators of Anthocyanin Biosynthesis in Tomato. Plant Cell Physiol. 2019, 60, 643–656. [Google Scholar] [CrossRef]
- Malacarne, G.; Coller, E.; Czemmel, S.; Vrhovsek, U.; Engelen, K.; Goremykin, V.; Bogs, J.; Moser, C. The grapevine VvibZIPC22 transcription factor is involved in the regulation of flavonoid biosynthesis. J. Exp. Bot. 2016, 67, 3509–3522. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Singh, H.; Singh, N.; Dwivedi, S.; Trivedi, P.K. Tobacco HY5, NtHY5, positively regulates flavonoid biosynthesis and enhances salt stress tolerance. bioRxiv 2022. [Google Scholar] [CrossRef]
- Morishita, T.; Kojima, Y.; Maruta, T.; Nishizawa-Yokoi, A.; Yabuta, Y.; Shigeoka, S. Arabidopsis NAC transcription factor, ANAC078, regulates flavonoid biosynthesis under high-light. Plant Cell Physiol. 2009, 50, 2210–2222. [Google Scholar] [CrossRef] [Green Version]
- An, X.H.; Tian, Y.; Chen, K.Q.; Liu, X.J.; Liu, D.D.; Xie, X.B.; Cheng, C.G.; Cong, P.H.; Hao, Y.J. MdMYB9 and MdMYB11 are involved in the regulation of the JA-induced biosynthesis of anthocyanin and proanthocyanidin in apples. Plant Cell Physiol. 2015, 56, 650–662. [Google Scholar] [CrossRef] [Green Version]
- Skirycz, A.; Jozefczuk, S.; Stobiecki, M.; Muth, D.; Zanor, M.I.; Witt, I.; Mueller-Roeber, B. Transcription factor AtDOF4; 2 affects phenylpropanoid metabolism in Arabidopsis thaliana. New Phytol. 2007, 175, 425–438. [Google Scholar] [CrossRef]
- Wang, N.; Liu, W.; Zhang, T.; Jiang, S.; Xu, H.; Wang, Y.; Zhang, Z.; Wang, C.; Chen, X. Transcriptomic analysis of red-fleshed apples reveals the novel role of MdWRKY11 in flavonoid and anthocyanin biosynthesis. J. Agr. Food Chem. 2018, 66, 7076–7086. [Google Scholar] [CrossRef]
- Liu, S.; Wang, L.; Cao, M.; Pang, S.; Wang, L. Identification and characterization of long non-coding RNAs regulating flavonoid biosynthesis in Ginkgo biloba leaves. Ind. Crop. Prod. 2020, 158, 112980. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, X.; Traore, S.M.; Xin, Z.; Yin, D. Genome-wide identification and analysis of long noncoding RNAs (lncRNAs) during seed development in peanut (Arachis hypogaea L.). BMC Plant Biol. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Gupta, O.P.; Karkute, S.G.; Banerjee, S.; Meena, N.L.; Dahuja, A. Contemporary understanding of miRNA-based regulation of secondary metabolites biosynthesis in plants. Front. Plant Sci. 2017, 8, 374. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Tiwari, M.; Pandey, A.; Bhatia, C.; Sharma, A.; Trivedi, P.K. MicroRNA858 is a potential regulator of phenylpropanoid pathway and plant development. Plant Physiol. 2016, 171, 944–959. [Google Scholar] [CrossRef] [Green Version]
- Glevitzky, I.; Dumitrel, G.A.; Glevitzky, M.; Manuela, B.P. Statistical analysis of the relationship between antioxidant activity and the structure of flavonoid compounds. Rev. Chim. 2019, 70, 3103–3107. [Google Scholar] [CrossRef]
- Zuo, A.R.; Dong, H.H.; Yu, Y.Y.; Shu, Q.L.; Zheng, L.X.; Yu, X.Y.; Cao, S.W. The antityrosinase and antioxidant activities of flavonoids dominated by the number and location of phenolic hydroxyl groups. Chin. Med. 2018, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kirschweng, B.; Tátraaljai, D.; Földes, E.; Pukánszky, B. Natural antioxidants as stabilizers for polymers. Polym. Degrad. Stabil. 2017, 145, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Engwa, G.A. Free Radicals and the Role of Plant Phytochemicals as Antioxidants Against Oxidative Stress-Related Diseases. In Phytochemicals-Source of Antioxidants and Role in Disease Prevention; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Lee-Hilz, Y.Y.; Boerboom, A.M.; Westphal, A.H.; Berkel, W.J.; Aarts, J.M.; Rietjens, I.M. Pro-oxidant activity of flavonoids induces EpRE-mediated gene expression. Chem. Res. Toxicol. 2006, 19, 1499–1505. [Google Scholar] [CrossRef]
- Zeng, N.; Zhang, G.; Hu, X.; Pan, J.; Gong, D. Mechanism of fisetin suppressing superoxide anion and xanthine oxidase activity. J. Funct. Foods 2019, 58, 1–10. [Google Scholar] [CrossRef]
- Md Idris, M.H.; Mohd Amin, S.N.; Mohd Amin, S.N.; Nyokat, N.; Khong, H.Y.; Selvaraj, M.; Zakaria, Z.A.; Shaameri, Z.; Hamzah, A.S.; Teh, L.K.; et al. Flavonoids as dual inhibitors of cyclooxygenase-2 (COX-2) and 5-lipoxygenase (5-LOX): Molecular docking and in vitro studies. Beni Suef Univ. J. Basic. Appl. Sci. 2022, 11, 117. [Google Scholar] [CrossRef]
- Samsonowicz, M.; Regulska, E.; Kalinowska, M. Hydroxyflavone metal complexes-molecular structure, antioxidant activity and biological effects. Chem. Biol. Interact. 2017, 273, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Kejík, Z.; Kaplánek, R.; Masařík, M.; Babula, P.; Matkowski, A.; Filipenský, P.; Veselá, K.; Gburek, J.; Sýkora, D.; Martásek, P.; et al. Iron Complexes of Flavonoids-Antioxidant Capacity and Beyond. Int. J. Mol. Sci. 2021, 22, 646. [Google Scholar] [CrossRef] [PubMed]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Simunkova, M.; Barbierikova, Z.; Jomova, K.; Hudecova, L.; Valko, M. Antioxidant vs. Prooxidant Properties of the Flavonoid, Kaempferol, in the Presence of Cu(II) Ions: A ROS-Scavenging Activity, Fenton Reaction and DNA Damage Study. Int. J. Mol. Sci. 2021, 22, 1619. [Google Scholar] [CrossRef]
- Zhang, K.; Dai, Z.; Zhang, W.; Gao, Q.; Dai, Y.; Xia, F.; Zhang, X. EDTA-based adsorbents for the removal of metal ions in wastewater. Coord. Chem. Rev. 2021, 434, 213809. [Google Scholar] [CrossRef]
- Grin, P.M.; Dwivedi, D.J.; Chathely, K.M.; Trigatti, B.L.; Prat, A.; Seidah, N.G.; Liaw, P.C.; Fox-Robichaud, A.E. Low-density lipoprotein (LDL)-dependent uptake of Gram-positive lipoteichoic acid and Gram-negative lipopolysaccharide occurs through LDL receptor. Sci. Rep. 2018, 8, 10496. [Google Scholar] [CrossRef] [Green Version]
- Torello, C.O.; Alvarez, M.C.; Olalla Saad, S.T. Polyphenolic Flavonoid Compound Quercetin Effects in the Treatment of Acute Myeloid Leukemia and Myelodysplastic Syndromes. Molecules 2021, 26, 5781. [Google Scholar] [CrossRef]
- Papi, S.; Ahmadizar, F.; Hasanvand, A. The role of nitric oxide in inflammation and oxidative stress. Immunopathol. Persa 2019, 5, e08. [Google Scholar] [CrossRef]
- Gutiérrez-Venegas, G.; Ventura-Arroyo, J.A.; Arreguín-Cano, J.A.; Ostoa-Pérez, M.F. Flavonoids inhibit iNOS production via mitogen activated proteins in lipoteichoic acid stimulated cardiomyoblasts. Int. Immunopharmacol. 2014, 21, 320–327. [Google Scholar] [CrossRef]
- Terao, J. Factors modulating bioavailability of quercetin-related flavonoids and the consequences of their vascular function. Biochem. Pharmacol. 2017, 139, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Li, Z.; Xie, Y.; Liu, L.; Cao, Y. Cyanidin chloride modestly protects Caco-2 cells from ZnO nanoparticle exposure probably through the induction of autophagy. Food Chem. Toxicol. 2019, 127, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Mirza, S.; Sharma, G.; Parshad, R.; Gupta, S.D.; Pandya, P.; Ralhan, R. Expression of DNA methyltransferases in breast cancer patients and to analyze the effect of natural compounds on DNA methyltransferases and associated proteins. J. Breast Cancer 2013, 16, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Quispe, C.; Chamkhi, I.; Omari, N.E.; Balahbib, A.; Sharifi-Rad, J.; Bouyahya, A.; Akram, M.; Iqbal, M.; Docea, A.O.; et al. Pharmacological Properties of Chalcones: A Review of Preclinical Including Molecular Mechanisms and Clinical Evidence. Front. Pharm. 2021, 11, 2068. [Google Scholar] [CrossRef]
- Sun, T.L.; Li, W.Q.; Tong, X.L.; Liu, X.Y.; Zhou, W.H. Xanthohumol attenuates isoprenaline-induced cardiac hypertrophy and fibrosis through regulating PTEN/AKT/mTOR pathway. Eur. J. Pharmacol. 2021, 891, 173690. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Yang, Z.Q.; Liu, F.; Peng, W.J.; Qu, S.Q.; Song, X.B.; Zhu, K.; Shen, J.Z. Antibacterial effect and mode of action of flavonoids from licorice against methicil-lin-resistant Staphylococcus aureus. Front. Microbiol. 2019, 10, 2489. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Xie, X.; Tu, Z.; Fu, J.; Xu, D.; Zhou, Y. The signal pathways and treatment of cytokine storm in COVID-19. Sig. Transduct. Target. Ther. 2021, 6, 255. [Google Scholar] [CrossRef]
- Efenberger-Szmechtyk, M.; Nowak, A.; Czyzowska, A. Plant extracts rich in polyphenols: Antibacterial agents and natural preservatives for meat and meat products. Crit. Rev. Food Sci. Nutr. 2021, 61, 149–178. [Google Scholar] [CrossRef]
- Tang, S.M.; Deng, X.T.; Zhou, J.; Li, Q.P.; Ge, X.X.; Miao, L. Pharmacological basis and new insights of quercetin action in respect to its anti-cancer effects. Biomed. Pharma. 2020, 121, 109604. [Google Scholar] [CrossRef]
- Barreca, D.; Gattuso, G.; Bellocco, E.; Calderaro, A.; Trombetta, D.; Smeriglio, A.; Lagana, G.; Daglia, M.; Menehini, S.; Nabavi, S.M. Flavanones: Citrus phytochemical with health-promoting properties. Biofactors 2017, 43, 495–506. [Google Scholar] [CrossRef]
- Chang, X.; Zhang, T.; Wang, J.; Liu, Y.; Yan, P.; Meng, Q.; Yin, Y.; Wang, S. SIRT5-related desuccinylation modification contributes to quercetin-induced protection against heart failure and high-glucose-prompted cardiomyocytes injured through regulation of mitochondrial quality surveillance. Oxid. Med. Cell. Longev. 2021, 2021, 5876841. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; Piragine, E.; Piano, I.; Flori, L.; Da Pozzo, E.; Miragliotta, V.; Pirone, A.; Citi, V.; Mannelli, L.D.C.; Brogi, S.; et al. The citrus flavonoid naringenin protects the myocardium from ageing-dependent dysfunction: Potential role of SIRT1. Oxid. Med. Cell. Longev. 2020, 2020, 4650207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xu, X.; Jiang, T.; Wu, K.; Ding, C.; Liu, Z.; Zhang, X.; Yu, T.; Song, C. Citrus aurantium Naringenin Prevents Osteosarcoma Progression and Recurrence in the Patients Who Underwent Osteosarcoma Surgery by Improving Antioxidant Capability. Oxid. Med. Cell. Longev. 2018, 7, 8713263. [Google Scholar] [CrossRef] [Green Version]
- Guardia, T.; Rotelli, A.E.; Juarez, A.O.; Pelzer, L.E. Anti-inflammatory properties of plant flavonoids. Effects of rutin, quercetin and hesperidin on adjuvant arthritis in rat. Il Farm. 2001, 56, 683–687. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F. A mechanistic review of the anticancer potential of hesperidin, a natural flavonoid from citrus fruits. Nutr. Res. 2021, 92, 21–31. [Google Scholar] [CrossRef]
- Yang, J.; Liu, L.; Li, M.; Huang, X.; Yang, H.; Li, K. Naringenin inhibits pro-inflammatory cytokine production in macrophages through inducing MT1G to suppress the activation of NF-κB. Mol. Immunol. 2021, 137, 155–162. [Google Scholar] [CrossRef]
- Zhong, L.; Lin, Y.; Wang, C.; Niu, B.; Xu, Y.; Zhao, G.; Zhao, J. Chemical Profile, Antimicrobial and Antioxidant Activity Assessment of the Crude Extract and Its Main Flavonoids from Tartary Buckwheat Sprouts. Molecules 2022, 27, 374. [Google Scholar] [CrossRef]
- Wu, C.; Chen, R.L.; Wang, Y.; Wu, W.Y.; Li, G. Acacetin alleviates myocardial ischemia/reperfusion injury by inhibiting oxidative stress and apoptosis via the Nrf-2/HO-1 pathway. Pharm. Biol. 2022, 60, 553–561. [Google Scholar] [CrossRef]
- Wu, Q.; Kroon, P.A.; Shao, H.; Needs, P. Differential effects of quercetin and two of its derivatives, isorhamnetin and isorhamnetin-3-glucuronide, in inhibiting the proliferation of human breast-cancer MCF-7 cells. J. Agr. Food Chem. 2018, 66, 7181–7189. [Google Scholar] [CrossRef]
- Lang, S.J.; Schmiech, M.; Hafner, S.; Paetz, C.; Simmet, T. Chrysosplenol d, a flavonol from Artemisia annua, induces ERK1/2-mediated apoptosis in triple negative human breast cancer cells. Int. J. Mol. Sci. 2020, 21, 4090. [Google Scholar] [CrossRef]
- Chen, X.; Wu, Y.; Gu, J.; Liang, P.; Qin, J. Anti-invasive effect and pharmacological mechanism of genistein against colorectal cancer. Biofactors 2020, 46, 620–628. [Google Scholar] [CrossRef]
- Hirchaud, F.; Hermetet, F.; Ablise, M.; Fauconnet, S.; Vuitton, D.A.; Prétet, J.L.; Mougin, C. Isoliquiritigenin induces caspase-dependent apoptosis via downregulation of HPV16 E6 expression in cervical cancer Ca Ski cells. Planta Med. 2013, 79, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Xu, X.; Huang, Y. Daidzin inhibits growth and induces apoptosis through the JAK2/STAT3 in human cervical cancer HeLa cells. Saudi J. Biol. Sci. 2021, 28, 7077–7081. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, M.; Sithisarn, P.; Cinatl, J. Effects of flavonoid-induced oxidative stress on anti-H5N1 influenza a virus activity exerted by baicalein and biochanin A. BMC Res. Notes. 2014, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guntaka, R.V. New Insights to Prevent Liver Fibrosis by Targeting YB-1 and Collagen Genes. OBM Hepatol. Gastroenterol. 2019, 3, 1. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, T.; Wang, Y.; Jiang, Y.; Wang, Y. Mechanisms and Advances in Anti-Ovarian Cancer with Natural Plants Component. Molecules 2021, 26, 5949. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo-Rinhel, A.S.; Santos, E.O.; Kabeya, L.M.; Azzolini, A.E.; Simões-Ambrosio, L.M.; Lucisano-Valim, Y.M. The flavonols quercetin, myricetin, kaempferol, and galangin inhibit the net oxygen consumption by immune complex-stimulated human and rabbit neutrophils. Z. Nat. C J. Biosci. 2014, 69, 346–356. [Google Scholar] [CrossRef]
- Farkhondeh, T.S.; Samarghandian Bafandeh, F. The Cardiovascular Protective Effects of Chrysin: A Narrative Review on Experimental Researches. Cardiovasc. Hematol. Agents Med. Chem. 2019, 17, 17–27. [Google Scholar] [CrossRef]
- Khazeei Tabari, M.A.; Iranpanah, A.; Bahramsoltani, R.; Rahimi, R. Flavonoids as promising antiviral agents against SARS-CoV-2 infection: A mechanistic review. Molecules 2021, 26, 3900. [Google Scholar] [CrossRef]
- Sugawara, T.; Sakamoto, K. Quercetin enhances motility in aged and heat-stressed Caenorhabditis elegans nematodes by modulating both HSF-1 activity, and insulin-like and p38-MAPK signalling. PLoS ONE 2020, 15, e0238528. [Google Scholar] [CrossRef]
- Sanches-Silva, A.; Testai, L.; Nabavi, S.F.; Battino, M.; Devi, K.P.; Tejada, S.; Sureda, A.; Xu, S.W.; Yousefi, B.; Majidinia, M.; et al. Therapeutic potential of polyphenols in cardiovascular diseases: Regulation of mTOR signaling pathway. Pharma. Res. 2020, 152, 104626. [Google Scholar] [CrossRef] [PubMed]
- Kiani, K.; Dhuli, K.; Anpilogov, K.; Bressan, S.; Dautaj, A.; Dundar, M.; Beccari, T.; Ergoren, M.; Bertelli, M. Natural compounds as inhibitors of SARS-CoV-2 endocytosis: A promising approach against COVID-19. Acta Med. 2020, 91, 13. [Google Scholar] [CrossRef]
- Kandhari, K.; Mishra, J.P.N.; Singh, R.P. Acacetin inhibits cell proliferation, survival, and migration in human breast cancer cells. Int. J. Pharma. Biol. Sci. 2019, 9, 443–452. [Google Scholar] [CrossRef]
- Zhou, B.; Fang, L.; Dong, Y.; Yang, J.; Chen, X.; Zhang, N.; Zhu, Y.; Huang, T. Mitochondrial quality control protects photoreceptors against oxidative stress in the H2O2-induced models of retinal degeneration diseases. Cell Death Dis. 2021, 12, 413. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, Y.; Pan, R.L.; Wang, R.Y.; Ding, S.L.; Dong, W.R.; Sun, X.B. Protective effects of Myrica rubra flavonoids against hypoxia/reoxygenation-induced cardiomyocyte injury via the regulation of the PI3K/Akt/GSK3β pathway. Int. J. Mol. Med. 2019, 43, 2133–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Yang, Y.; An, Y.; Fang, G. The mechanism of anticancer action and potential clinical use of kaempferol in the treatment of breast cancer. Biomed. Pharm. 2019, 117, 109086. [Google Scholar] [CrossRef]
- Chan, K.K.L.; Siu, M.K.Y.; Jiang, Y.X.; Wang, J.J.; Leung, T.H.Y.; Ngan, H.Y.S. Estrogen receptor modulators genistein, daidzein and ERB-041 inhibit cell migration, invasion, proliferation and sphere formation via modulation of FAK and PI3K/AKT signaling in ovarian cancer. Cancer Cell Int. 2018, 18, 65. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.J.; Deng, A.J.; Zhang, Z.H.; Yu, Z.H.; Liu, Y.; Peng, S.Y.; Wu, L.Q.; Qin, H.L.; Wang, W.J. The protective effect of epicatechin on experimental ulcerative colitis in mice is mediated by increasing antioxidation and by the inhibition of NF-κB pathway. Pharm. Rep. 2016, 68, 514–520. [Google Scholar] [CrossRef]
- Prince, P.S.M. A biochemical, electrocardiographic, electrophoretic, histopathological and in vitro study on the protective effects of (−) epicatechin in isoproterenol-induced myocardial infarcted rats. Eur. J. Pharm. 2011, 671, 95–101. [Google Scholar] [CrossRef]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets. Sig. Transduct. Target. Ther. 2021, 6, 94. [Google Scholar] [CrossRef]
- Ribeiro, D.; Freitas, M.; Tomé, S.M.; Silva, A.M.S.; Laufer, S.; Lima, J.L.F.C.; Fernandes, E. Flavonoids Inhibit COX-1 and COX-2 Enzymes and Cytokine/Chemokine Production in Human Whole Blood. Inflammation 2014, 38, 858–870. [Google Scholar] [CrossRef] [PubMed]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yue, S.J. Editorial: Flavonoids and Cardiovascular Metabolism. Front. Nutr. 2022, 9, 939798. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.M.; Liu, Y.J.; Huang, Y.; Yu, H.J.; Yuan, S.; Tang, B.W.; Wang, P.G.; He, Q.Q. Dietary total flavonoids intake and risk of mortality from all causes and cardiovascular disease in the general population: A systematic review and meta-analysis of cohort studies. Mol. Nutr. Food Res. 2017, 61, 1601003. [Google Scholar] [CrossRef]
- Sanchez, M.; Romero, M.; Gomez-Guzman, M.; Tamargo, J.; Perez-Vizcaino, F.; Duarte, J. Cardiovascular effects of flavonoids. Curr. Med. Chem. 2019, 26, 6991–7034. [Google Scholar] [CrossRef]
- Mulvihill, E.E.; Burke, A.C.; Huff, M.W. Citrus Flavonoids as Regulators of Lipoprotein Metabolism and Atherosclerosis. Annu. Rev. Nutr. 2016, 36, 275–299. [Google Scholar] [CrossRef]
- Chacko, B.K.; Chandler, R.T.; D’Alessandro, T.L.; Mundhekar, A.; Khoo, N.K.H.; Botting, N.; Barnes, S.; Patel, R.P. Anti-inflammatory effects of isoflavones are dependent on flow and human endothelial cell PPARgamma. J. Nutr. 2007, 137, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Wallace, T.C.; Slavin, M.; Frankenfeld, C.L. Systematic Review of Anthocyanins and Markers of Cardiovascular Disease. Nutrients 2016, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Chu, Q.; Zhang, S.; Chen, M.; Ha, W.; Ji, R.; Chen, W.; Zheng, X. Cherry anthocyanins regulate NAFLD by promoting autophagy pathway. Oxid. Med. Cell. Longev. 2019, 2019, 4825949. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Zhang, B.Y.; Peng, Y.F.; Chang, L.C.; Li, Z.Q.; Zhang, X.X.; Zhang, D.J. Mechanism of action of flavonoids of oxytropis falcata on the alleviation of myocardial ischemia–reperfusion injury. Molecules 2022, 27, 1706. [Google Scholar] [CrossRef]
- Sebghatollahi, Z.; Ghanadian, M.; Agarwal, P.; Ghaheh, H.S.; Mahato, N.; Yogesh, R. Citrus flavonoids: Biological activities, implementation in skin health, and topical applications: A Review. ACS Food Sci. Technol. 2022, 2, 1417–1432. [Google Scholar] [CrossRef]
- Chambers, C.S.; Viktorová, J.; Rěhorŏvá, K.; Biedermann, D.; Turková, L.; Macek, T.; Křen, V.; Valentová, K. Defying multidrug resistance! Modulation of related transporters by flavonoids and flavonolignans. J. Agric. Food Chem. 2019, 68, 1763–1779. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, X.; He, D.; Zou, D.; Zhao, R.; Wang, H.; Li, S.; Xu, Y.; Abudureheman, B. Optimization of flavonoid extraction from Xanthoceras sorbifolia Bunge flowers, and the antioxidant and antibacterial capacity of the extract. Molecules 2021, 27, 113. [Google Scholar] [CrossRef]
- Yuan, G.; Guan, Y.; Yi, H.; Lai, S.; Sun, Y.; Cao, S. Antibacterial activity and mechanism of plant flavonoids to gram-positive bacteria predicted from their lipophilicities. Sci. Rep. 2021, 11, 10471. [Google Scholar] [CrossRef] [PubMed]
- Al Aboody, M.S.; Mickymaray, S. Anti-fungal efficacy and mechanisms of flavonoids. Antibiotics 2020, 9, 45. [Google Scholar] [CrossRef] [Green Version]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Ag. 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Pasupuleti, V.R.; Arigela, C.S.; Gan, S.H.; Salam, S.K.N.; Krishnan, K.T.; Rahman, N.A.; Jeffree, M.S. A review on oxidative stress, diabetic complications, and the roles of honey polyphenols. Oxid. Med. Cell. Longev. 2020, 2020, 8878172. [Google Scholar] [CrossRef]
- Morkunas, I.; Ratajczak, L. The role of sugar signaling in plant defense responses against fungal pathogens. Acta Physiol. Plant. 2014, 36, 1607–1619. [Google Scholar] [CrossRef] [Green Version]
- Traganos, F.; Kimmel, M.; Bueti, C.; Darzynkiewicz, Z. Effects of inhibition of RNA or protein synthesis on CHO cell cycle progression. J. Cell. Physiol. 1987, 133, 277–287. [Google Scholar] [CrossRef]
- Rajendran, P.; Rengarajan, T.; Nandakumar, N.; Palaniswami, R.; Nishigaki, Y.; Nishigaki, I. Kaempferol, a potential cytostatic and cure for inflammatory disorders. Eur. J. Med. Chem. 2014, 86, 103–112. [Google Scholar] [CrossRef]
- Li, A.; Zhao, Z.; Zhang, S.; Zhang, Z.; Shi, Y. Fungicidal activity and mechanism of action of glabridin from Glycyrrhiza glabra L. Int. J. Mol. Sci. 2021, 22, 10966. [Google Scholar] [CrossRef]
- Lalani, S.; Poh, C.L. Flavonoids as antiviral agents for Enterovirus A71 (EV-A71). Viruses 2020, 12, 184. [Google Scholar] [CrossRef] [Green Version]
- Roschek, B.; Fink, R.C.; McMichael, M.D.; Li, D.; Alberte, R.S.J.P. Elderberry flavonoids bind to and prevent H1N1 infection in vitro. Phytochemistry 2009, 70, 1255–1261. [Google Scholar] [CrossRef]
- Sithisarn, P.; Michaelis, M.; Schubert-Zsilavecz, M.; Cinatl, J., Jr. Differential antiviral and anti-inflammatory mechanisms of the flavonoids biochanin A and baicalein in H5N1 influenza A virus-infected cells. Antivir. Res. 2013, 97, 41–48. [Google Scholar] [CrossRef]
- Badshah, S.L.; Faisal, S.; Muhammad, A.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Antiviral activities of flavonoids. Biomed. Pharmacother. 2021, 140, 111596. [Google Scholar] [CrossRef] [PubMed]
- Ezzati, M.; Yousefi, B.; Velaei, K.; Safa, A. A review on anti-cancer properties of Quercetin in breast cancer. Life Sci. 2020, 248, 117463. [Google Scholar] [CrossRef]
- Veeramuthu, D.; Raja, W.R.T.; Al-Dhabi, N.A.; Savarimuthu, I. Flavonoids: Anticancer properties. In Flavonoids: From Biosynthesis to Human Health; IntechOpen: London, UK, 2017; p. 287. [Google Scholar]
- Coelho, P.L.C.; Oliveira, M.N.; da Silva, A.B.; Pitanga, B.P.; Silva, V.D.; Faria, G.P.; Sampaio, G.P.; Costa, M.D.D.; Braga-de-Souza, S.; Costa, S.L. The flavonoid apigenin from Croton betulaster Mull inhibits proliferation, induces differentiation and regulates the inflammatory profile of glioma cells. Anticancer Drug. 2016, 27, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.A.; Jordan, H.R.; Tollefsbol, T.O. Effffects of SAHA and EGCG on Growth Potentiation of Triple-Negative Breast Cancer Cells. Cancers 2018, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Adinew, G.M.; Taka, E.; Mendonca, P.; Messeha, S.S.; Soliman, K.F. The anticancer effects of flavonoids through miRNAs modulations in triple-negative breast cancer. Nutrients 2021, 13, 1212. [Google Scholar] [CrossRef] [PubMed]
- Lecumberri, E.; Dupertuis, Y.M.; Miralbell, R.; Pichard, C. Green tea polyphenol epigallocatechin-3-gallate (EGCG) as adjuvant in cancer therapy. Clin. Nutr. 2013, 32, 894–903. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Tollefsbol, T.O. Combinational Proanthocyanidins and Resveratrol Synergistically Inhibit Human Breast Cancer Cells and Impact Epigenetic-Mediating Machinery. Int. J. Mol. Sci. 2018, 19, 2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvakumar, P.; Badgeley, A.; Murphy, P.; Anwar, H.; Sharma, U.; Lawrence, K.; Lakshmikuttyamma, A. Flavonoids and Other Polyphenols Act as Epigenetic Modifiers in Breast Cancer. Nutrients 2020, 12, 761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Liu, Y.; Zhao, Z.; Qiu, J. Oxidative stress in the skin: Impact and related protection. Int. J. Cosmet. Sci. 2021, 43, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Fokou, P.V.T.; Azzini, E.; Peluso, I.; et al. Lifestyle, oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.T.; Moon, J.Y.; Lee, Y.C. Natural Antioxidants from Plant Extracts in Skincare Cosmetics: Recent Applications, Challenges and Perspectives. Cosmetics 2021, 8, 106. [Google Scholar] [CrossRef]
- Saewan, N.; Jimtaisong, A. Natural products as photoprotection. J. Cosmet. Dermatol. 2015, 14, 47–63. [Google Scholar] [CrossRef]
- Saewan, N.; Jimtaisong, A. Photoprotection of natural flavonoids. J. Appl. Pharma Sci. 2013, 3, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Mottaghipisheh, J.; Taghrir, H.; Boveiri Dehsheikh, A.; Zomorodian, K.; Irajie, C.; Mahmoodi Sourestani, M.; Iraji, A. Linarin, a Glycosylated Flavonoid, with Potential Therapeutic Attributes: A Comprehensive Review. Pharmaceuticals 2021, 14, 1104. [Google Scholar] [CrossRef]
- Atif, A.; Naveed, A.; Muhammad, S.K.; Fatima, R.; Furqan, M.I.; Muhammad, T.; Minhaj, U.; Ehsan, E. Moisturizing effect of cream containing Moringa oleifera (Sohajana) leaf extract by biophysical techniques: In vivo evaluation. J. Med. Plants Res. 2013, 7, 386–391. [Google Scholar] [CrossRef]
- Cordeiro, M.L.S.; Martins, V.G.Q.A.; Silva, A.P.; Rocha, H.A.O.; Rachetti, V.P.S.; Scortecci, K.C. Phenolic Acids as Antidepressant Agents. Nutrients 2022, 14, 4309. [Google Scholar] [CrossRef]
- Aboonabi, A.; Singh, I. Chemopreventive role of anthocyanins in atherosclerosis via activation of Nrf2–ARE as an indicator and modulator of redox. Biomed. Pharmacother. 2015, 72, 30–36. [Google Scholar] [CrossRef]
- Tran, P.L.P.; Le, H.P. Plant Flavonoids as Potential Natural Antioxidants in Phytocosmetics. J. Tech. Educ. Sci. 2022, 70B, 86–93. [Google Scholar] [CrossRef]
- Bjorklund, G.; Shanaida, M.; Lysiuk, R.; Butnariu, M.; Peana, M.; Sarac, I.; Strus, O.; Smetanina, K.; Chirumbolo, S. Natural Compounds and Products from an Anti-Aging Perspective. Molecules 2022, 27, 7084. [Google Scholar] [CrossRef]
- Micek, I.; Nawrot, J.; Seraszek-Jaros, A.; Jenerowicz, D.; Schroeder, G.; Spiżewski, T.; Suchan, A.; Pawlaczyk, M.; Gornowicz-Porowska, J. Taxifolin as a Promising Ingredient of Cosmetics for Adult Skin. Antioxidants 2021, 10, 1625. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Chang, B.Y.; Kim, D.S.; Cho, H.K.; Kiml, S.Y. Effects of the Syzygium aromaticum L. extract on antioxidation and inhibition of matrix metalloproteinase in human dermal fibroblast. Asian Pac. J. Trop. Bio. 2019, 9, 53. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phung, H.M.; Lee, S.; Hong, S.; Lee, S.; Jung, K.; Kang, K.S. Protective Effect of Polymethoxyflavones Isolated from Kaempferia parviflora against TNF-α-Induced Human Dermal Fibroblast Damage. Antioxidants 2021, 10, 1609. [Google Scholar] [CrossRef]
- Lee, S.; Jang, T.; Kim, K.H.; Kang, K.S. Improvement of Damage in Human Dermal Fibroblasts by 3, 5, 7-Trimethoxyflavone from Black Ginger (Kaempferia parviflora). Antioxidants 2022, 11, 425. [Google Scholar] [CrossRef]
- Chaiprasongsuk, A.; Panich, U. Role of phytochemicals in skin photoprotection via regulation of Nrf2. Front. Pharma. 2022, 13, 823881. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Twinomuhwezi, H.; Victory, I.S.; Ikechukwu, A.O. Food Additives and Food Preservatives for Domestic and Industrial Food Applications. J. Anim. Health 2020, 2, 1–16. [Google Scholar]
- Ruiz-Cruz, S.; Chaparro-Hernández, S.; Hernández-Ruiz, K.L.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Gassos-Ortega, L.E.; Ornelas-Paz, J.; Lopez Mata, M.A. Flavonoids: Important biocompounds in food. In Flavonoids: From Biosynthesis to Human. Health; IntechOpen: London, UK, 2017; pp. 353–369. [Google Scholar] [CrossRef] [Green Version]
- Prakash, D.; Kumar, N. Cost Effective Natural Antioxidants. Nutrients, Dietary Supplements, and Nutriceuticals; Humana Press: Totowa, NJ, USA, 2011; pp. 163–187. [Google Scholar] [CrossRef]
- Babuskin, S.; Babu, P.A.S.; Sasikala, M.; Sabina, K.; Archana, G.; Sivarajan, M.; Sukumar, M. Antimicrobial and antioxidant effects of spice extracts on the shelf life extension of raw chicken meat. Int. J. Food Microbiol. 2014, 171, 32–40. [Google Scholar] [CrossRef]
- Ji, H.F.; Li, X.J.; Zhang, H.Y. Natural products and drug discovery: Can thousands of years of ancient medical knowledge lead us to new and powerful drug combinations in the fight against cancer and dementia? EMBO Rep. 2009, 10, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-del-Río, I.; López-Ibáñez, S.; Magadán-Corpas, P.; Fernández-Calleja, L.; Pérez-Valero, A.; Tuñón-Granda, M.; Miguelez, E.; Villar, C.; Lombo, F. Terpenoids and polyphenols as natural antioxidant agents in food preservation. Antioxidants 2021, 10, 1264. [Google Scholar] [CrossRef]
- Rathod, N.B.; Ranveer, R.C.; Benjakul, S.; Kim, S.K.; Pagarkar, A.U.; Patange, S.; Ozogul, F. Recent developments of natural antimicrobials and antioxidants on fish and fishery food products. Compre. Rev. Food Sci. Food Saf. 2021, 20, 4182–4210. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, C.; Kia, D.A.; Vandrovcova, J.; Hardy, J.; Wood, N.W.; Lewis, P.A.; Ferrari, R. Genome, transcriptome and proteome: The rise of omics data and their integration in biomedical sciences. Brief. Bioinform. 2018, 19, 286–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Ruan, Z.; Fei, Z.; Yan, J.; Tang, G. Integrated transcriptome and metabolome analysis revealed that flavonoid biosynthesis may dominate the resistance of Zanthoxylum bungeanum against stem canker. J. Agric. Food Chem. 2021, 69, 6360–6378. [Google Scholar] [CrossRef]
- Yuan, Y.; Zuo, J.; Zhang, H.; Zu, M.; Yu, M.; Liu, S. Transcriptome and metabolome profiling unveil the accumulation of flavonoids in Dendrobium officinale. Genomics 2022, 114, 110324. [Google Scholar] [CrossRef]
- Zhang, Z.; Tian, C.; Zhang, Y.; Li, C.; Li, X.; Yu, Q.; Wang, S.; Wang, X.; Chen, X.; Feng, S. Transcriptomic and metabolomic analysis provides insights into anthocyanin and procyanidin accumulation in pear. BMC Plant Biol. 2020, 20, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, L.; Liu, Z.; Zhao, Z.; Zhao, J.; Wang, Z.; Zhou, G.; Liu, P.; Liu, M. Transcriptome and metabolome profiling unveil the mechanisms of Ziziphus jujuba Mill. peel coloration. Food Chem. 2020, 312, 125903. [Google Scholar] [CrossRef]
- Peng, X.Q.; Ai, Y.J.; Pu, Y.T.; Wang, X.J.; Li, Y.H.; Wang, Z.; Zhuang, W.B.; Yu, B.J.; Zhu, Z.Q. Transcriptome and metabolome analyses reveal molecular mechanisms of anthocyanin-related leaf color variation in poplar (Populus deltoides) cultivars. Front. Plant Sci. 2023, 14, 554. [Google Scholar] [CrossRef]
- Teng, H.; Zheng, Y.; Cao, H.; Huang, Q.; Xiao, J.; Chen, L. Enhancement of bioavailability and bioactivity of diet-derived flavonoids by application of nanotechnology: A review. Crit. Rev. Food Sci. Nutr. 2023, 63, 378–393. [Google Scholar] [CrossRef]
- Jannat, K.; Paul, A.K.; Bondhon, T.A.; Hasan, A.; Nawaz, M.; Jahan, R.; Mahboob, T.; Nissapatorn, V.; Wilairatana, P.; Pereira, M.d.L.; et al. Nanotechnology Applications of Flavonoids for Viral Diseases. Pharmaceutics 2021, 13, 1895. [Google Scholar] [CrossRef] [PubMed]
- Dobrzynska, M.; Napierala, M.; Florek, E. Flavonoid nanoparticles: A promising approach for cancer therapy. Biomolecules 2020, 10, 1268. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Adhami, V.M.; Bharali, D.J.; Hafeez, B.B.; Asim, M.; Khwaja, S.I.; Ahmad, N.; Cui, H.D.; Mousa, S.A.; Mukhtar, H. Introducing nanochemoprevention as a novel approach for cancer control: Proof of principle with green tea polyphenol epigallocatechin-3-gallate. Cancer Res. 2009, 69, 1712–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.J.; Liu, Y.; Chang, K.L.; Lim, B.K.; Chiu, G.N. Perorally active nanomicellar formulation of quercetin in the treatment of lung cancer. Int. J. Nanomed. 2012, 7, 651–661. [Google Scholar] [CrossRef] [Green Version]
- Sindhu, R.K.; Verma, R.; Salgotra, T.; Rahman, M.H.; Shah, M.; Akter, R.; Murad, W.; Mubin, S.; Bibi, P.; Qusti, S.; et al. Impacting the remedial potential of nano delivery-based flavonoids for breast cancer treatment. Molecules 2021, 26, 5163. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Wang, X.; Cheng, Y.; Gao, H.; Chen, X. A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids. Molecules 2023, 28, 4982. https://doi.org/10.3390/molecules28134982
Chen S, Wang X, Cheng Y, Gao H, Chen X. A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids. Molecules. 2023; 28(13):4982. https://doi.org/10.3390/molecules28134982
Chicago/Turabian StyleChen, Shen, Xiaojing Wang, Yu Cheng, Hongsheng Gao, and Xuehao Chen. 2023. "A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids" Molecules 28, no. 13: 4982. https://doi.org/10.3390/molecules28134982
APA StyleChen, S., Wang, X., Cheng, Y., Gao, H., & Chen, X. (2023). A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids. Molecules, 28(13), 4982. https://doi.org/10.3390/molecules28134982