
Chronotoxicity of Acrylamide in Mice Fed a High-Fat Diet: The Involvement of Liver CYP2E1 Upregulation and Gut Leakage

Abstract
:1. Introduction
2. Results
2.1. The Effects of the HFD on the Survival Rate in ACR-Treated Mice at Treated at ZT0 or ZT12
2.2. The Differential Effects of ACR Administration on Liver Damage at ZT0 and ZT12 in HFD-Fed Mice
2.3. The Differential Effects of ACR Administration on Liver Mitochondrial Function at ZT0 and ZT12 in HFD-Fed Mice
2.4. The Differential Effects of ACR Administration on CYP2E1 Expression at ZT0 and ZT12 in HFD-Fed Mice
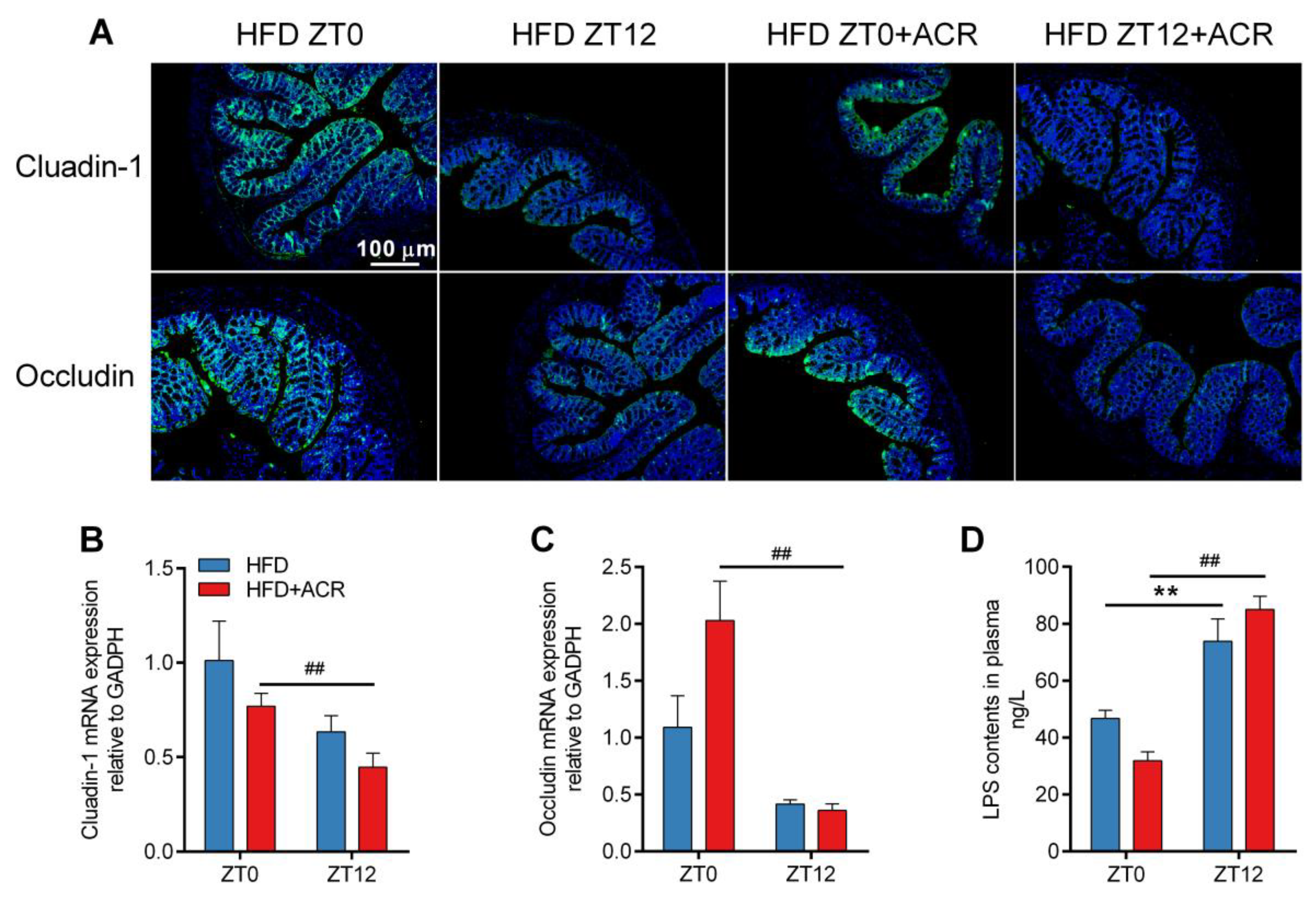
2.5. The Differential Effects of ACR Administration on the Gut Barrier Integrity and Inflammatory Responses at ZT0 and ZT12 in HFD-Fed Mice
3. Discussion
4. Materials and Methods
4.1. Animals and Treatments
4.2. RNA Preparation and RT-qPCR
4.3. Western Blots
4.4. Serum Analysis
4.5. Measurement of Hepatic CYP2E1 Activity
4.6. Hematoxylin and Eosin (H&E), Immunohistochemical (IHC), and Immunofluorescence Staining
4.7. Structural Analysis of the Liver Samples by Transmission Electron Microscopy (TEM)
4.8. Comet Assay
4.9. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Tan, X.; Zhao, T.; Wang, Z.; Wang, J.; Wang, Y.; Liu, Z.; Liu, X. Acrylamide Defects the Expression Pattern of the Circadian Clock and Mitochondrial Dynamics in C57BL/6J Mice Liver and HepG2 Cells. J. Agric. Food Chem. 2018, 66, 10252–10266. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Ye, J.; Liu, W.; Zhao, B.; Shi, X.; Zhang, C.; Liu, Z.; Liu, X. Acrylamide aggravates cognitive deficits at night period via the gut–brain axis by reprogramming the brain circadian clock. Arch. Toxicol. 2019, 93, 467–486. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Song, G.; Zou, C.; Liu, G.; Wu, W.; Yuan, T.; Liu, X. Acrylamide induces mitochondrial dysfunction and apoptosis in BV-2 microglial cells. Free Radic. Biol. Med. 2015, 84, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Bhattacharya, B.; Agarwal, T.; Paul, V.; Chakkaravarthi, S. Integrated approach towards acrylamide reduction in potato-based snacks: A critical review. Food Res. Int. 2022, 156, 111172. [Google Scholar] [CrossRef]
- Song, G.; Liu, Z.; Liu, Q.; Liu, X. Lipoic acid prevents acrylamide-induced neurotoxicity in CD-1 mice and BV2 microglial cells via maintaining redox homeostasis. J. Funct. Foods 2017, 35, 363–375. [Google Scholar] [CrossRef]
- Varga, N. Acrylamide from Maillard reaction products. Nature 2002, 419, 449–450. [Google Scholar]
- Mottram, D.S.; Wedzicha, B.L.; Dodson, A.T. Acrylamide is formed in the Maillard reaction. Nature 2002, 419, 448–449. [Google Scholar] [CrossRef]
- Gamboa da Costa, G.; Churchwell, M.I.; Hamilton, L.P.; Von Tungeln, L.S.; Beland, F.A.; Marques, M.M.; Doerge, D.R. DNA adduct formation from acrylamide via conversion to glycidamide in adult and neonatal mice. Chem. Res. Toxicol. 2003, 16, 1328–1337. [Google Scholar] [CrossRef]
- Doerge, D.R.; da Costa, G.G.; McDaniel, L.P.; Churchwell, M.I.; Twaddle, N.C.; Beland, F.A. DNA adducts derived from administration of acrylamide and glycidamide to mice and rats. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2005, 580, 131–141. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, P.; Zhu, Y.; Liu, X.; Hu, X.; Chen, F. The chemoprotection of a blueberry anthocyanin extract against the acrylamide-induced oxidative stress in mitochondria: Unequivocal evidence in mice liver. Food Funct. 2015, 6, 3006–3012. [Google Scholar] [CrossRef]
- Panda, S. Circadian physiology of metabolism. Science 2016, 354, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Lu, B. The effects of phytochemicals on circadian rhythm and related diseases. Crit. Rev. Food Sci. Nutr. 2019, 59, 882–892. [Google Scholar] [CrossRef]
- Claudel, T.; Cretenet, G.; Saumet, A.; Gachon, F. Crosstalk between xenobiotics metabolism and circadian clock. FEBS Lett. 2007, 581, 3626–3633. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.P.; Walisser, J.A.; Yan, L.; Shen, A.L.; Mcdearmon, E.L.; Moran, S.M.; Mcintosh, B.E.; Vollrath, A.L.; Schook, A.C.; Takahashi, J.S. Hepatocyte circadian clock controls acetaminophen bioactivation through NADPH-cytochrome P450 oxidoreductase. Proc. Natl. Acad. Sci. USA 2014, 111, 18757–18762. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Qiao, Q.; Sun, Y.; Chen, Y.; Ren, B.; Liu, X. Sesamol ameliorates diet induced obesity in C57BL/6J mice and suppresses adipogenesis in 3T3-L1 cells via regulating mitochondria-lipid metabolism. Mol. Nutr. Food Res. 2017, 61, 1600717. [Google Scholar] [CrossRef]
- Popkin, B.M.; Adair, L.S.; Wen, N.S. Global nutrition transition and the pandemic of obesity in developing countries. Nutr. Rev. 2012, 70, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Mi, Y.; Qi, G.; Fan, R.; Ji, X.; Liu, Z.; Liu, X. EGCG ameliorates diet-induced metabolic syndrome associating with the circadian clock. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2017, 1863, 1575–1589. [Google Scholar] [CrossRef]
- Mi, Y.; Qi, G.; Gao, Y.; Li, R.; Wang, Y.; Li, X.; Huang, S.; Liu, X. (-)-Epigallocatechin-3-gallate Ameliorates Insulin Resistance and Mitochondrial Dysfunction in HepG2 Cells: Involvement of Bmal1. Mol. Nutr. Food Res. 2017, 61, 1700440. [Google Scholar] [CrossRef]
- Ghanayem, B.I.; Bai, R.; Kissling, G.E.; Travlos, G.; Hoffler, U. Diet-induced obesity in male mice is associated with reduced fertility and potentiation of acrylamide-induced reproductive toxicity. Biol. Reprod. 2010, 82, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Khemawoot, P.; Yokogawa, K.; Shimada, T.; Miyamoto, K.-I. Obesity-induced increase of CYP2E1 activity and its effect on disposition kinetics of chlorzoxazone in Zucker rats. Biochem. Pharmacol. 2007, 73, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Abdelmegeed, M.A.; Banerjee, A.; Yoo, S.-H.; Jang, S.; Gonzalez, F.J.; Song, B.-J. Critical role of cytochrome P450 2E1 (CYP2E1) in the development of high fat-induced non-alcoholic steatohepatitis. J. Hepatol. 2012, 57, 860–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, E.E.; Hsu, T.M.; Kanoski, S.E. Gut to brain dysbiosis: Mechanisms linking western diet consumption, the microbiome, and cognitive impairment. Front. Behav. Neurosci. 2017, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazier, T.H.; DiBaise, J.K.; McClain, C.J. Gut microbiota, intestinal permeability, obesity-induced inflammation, and liver injury. J. Parenter. Enter. Nutr. 2011, 35 (Suppl. S5), 14S–20S. [Google Scholar] [CrossRef] [PubMed]
- Oh-Oka, K.; Kono, H.; Ishimaru, K.; Miyake, K.; Kubota, T.; Ogawa, H.; Okumura, K.; Shibata, S.; Nakao, A. Expressions of Tight Junction Proteins Occludin and Claudin-1 Are under the Circadian Control in the Mouse Large Intestine: Implications in Intestinal Permeability and Susceptibility to Colitis. PLoS ONE 2014, 9, e98016. [Google Scholar] [CrossRef]
- Erkekoglu, P.; Baydar, T. Acrylamide neurotoxicity. Nutr. Neurosci. 2014, 17, 49–57. [Google Scholar] [CrossRef]
- Jacobi, D.; Liu, S.; Burkewitz, K.; Kory, N.; Knudsen, N.H.; Alexander, R.K.; Unluturk, U.; Li, X.; Kong, X.; Hyde, A.L. Hepatic Bmal1 Regulates Rhythmic Mitochondrial Dynamics and Promotes Metabolic Fitness. Cell Metab. 2015, 22, 709–720. [Google Scholar] [CrossRef] [Green Version]
- Ghanayem, B.I.; Witt, K.L.; Kissling, G.E.; Tice, R.R.; Recio, L. Absence of acrylamide-induced genotoxicity in CYP2E1-null mice: Evidence consistent with a glycidamide-mediated effect. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2005, 578, 284–297. [Google Scholar] [CrossRef]
- Matsunaga, N.; Ikeda, M.; Takiguchi, T.; Koyanagi, S.; Ohdo, S. The molecular mechanism regulating 24-hour rhythm of CYP2E1 expression in the mouse liver. Hepatology 2008, 48, 240–251. [Google Scholar] [CrossRef]
- Lee, S.H. Intestinal permeability regulation by tight junction: Implication on inflammatory bowel diseases. Intest. Res. 2015, 13, 11. [Google Scholar] [CrossRef] [Green Version]
- Timmermann, C.A.G.; Mølck, S.S.; Kadawathagedara, M.; Bjerregaard, A.A.; Törnqvist, M.; Brantsaeter, A.L.; Pedersen, M. A review of dietary intake of acrylamide in humans. Toxics 2021, 9, 155. [Google Scholar] [CrossRef]
- Seiber, J.N. Modern issues in food safety—A perspective. J. Integr. Agric. 2012, 11, 9–13. [Google Scholar] [CrossRef]
- Park, H.R.; Kim, M.-S.; Kim, S.J.; Park, M.; Kong, K.H.; Kim, H.S.; Kwack, S.J.; Kang, T.S.; Kim, S.H.; Kim, H.S. Acrylamide induces cell death in neural progenitor cells and impairs hippocampal neurogenesis. Toxicol. Lett. 2010, 193, 86–93. [Google Scholar] [CrossRef]
- Marković Filipović, J.; Miler, M.; Kojić, D.; Karan, J.; Ivelja, I.; Čukuranović Kokoris, J.; Matavulj, M. Effect of Acrylamide Treatment on Cyp2e1 Expression and Redox Status in Rat Hepatocytes. Int. J. Mol. Sci. 2022, 23, 6062. [Google Scholar] [CrossRef]
- Tan, X.; Li, L.; Wang, J.; Zhao, B.; Pan, J.; Wang, L.; Liu, X.; Liu, X.; Liu, Z. Resveratrol prevents acrylamide-induced mitochondrial dysfunction and inflammatory responses via targeting circadian regular Bmal1 and Cry1 in hepatocytes. J. Agric. Food Chem. 2019, 67, 8510–8519. [Google Scholar] [CrossRef]
- Petrov, D.; Pedrós, I.; Artiach, G.; Sureda, F.X.; Barroso, E.; Pallàs, M.; Casadesús, G.; Beas-Zarate, C.; Carro, E.; Ferrer, I. High-fat diet-induced deregulation of hippocampal insulin signaling and mitochondrial homeostasis deficiences contribute to Alzheimer disease pathology in rodents. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2015, 1852, 1687–1699. [Google Scholar] [CrossRef] [Green Version]
- Vial, G.; Dubouchaud, H.; Couturier, K.; Cottet-Rousselle, C.; Taleux, N.; Athias, A.; Galinier, A.; Casteilla, L.; Leverve, X.M. Effects of a high-fat diet on energy metabolism and ROS production in rat liver. J. Hepatol. 2011, 54, 348–356. [Google Scholar] [CrossRef]
- Matsuzawa-Nagata, N.; Takamura, T.; Ando, H.; Nakamura, S.; Kurita, S.; Misu, H.; Ota, T.; Yokoyama, M.; Honda, M.; Miyamoto, K.-i. Increased oxidative stress precedes the onset of high-fat diet–induced insulin resistance and obesity. Metabolism 2008, 57, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Melissa, W.; Mary, N.; Hasan, M.; Nihal, A. Circadian rhythm connections to oxidative stress: Implications for human health. Antioxid. Redox Signal. 2013, 19, 192–208. [Google Scholar]
- Kurebayashi, H.; Ohno, Y. Metabolism of acrylamide to glycidamide and their cytotoxicity in isolated rat hepatocytes: Protective effects of GSH precursors. Arch. Toxicol. 2006, 80, 820. [Google Scholar] [CrossRef]
- Gonzalez, F.J. Role of cytochromes P450 in chemical toxicity and oxidative stress: Studies with CYP2E1. Mutat. Res. 2005, 569, 101–110. [Google Scholar] [CrossRef]
- Leung, T.-M.; Nieto, N. CYP2E1 and oxidant stress in alcoholic and non-alcoholic fatty liver disease. J. Hepatol. 2013, 58, 395–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, S.; Tian, L.; Zeng, L.; Luo, H.; Yang, X.; Na, L.; Chen, X.; Liu, Z.; Rui, L.; Win, S. Gut microbiota mediates diurnal variation of acetaminophen induced acute liver injury in mice. J. Hepatol. 2018, 69, S0168827818301466. [Google Scholar] [CrossRef] [PubMed]
- Compare, D.; Coccoli, P.; Rocco, A.; Nardone, O.; De Maria, S.; Cartenì, M.; Nardone, G. Gut–liver axis: The impact of gut microbiota on non alcoholic fatty liver disease. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006, 26, 1175–1186. [Google Scholar] [CrossRef]
- Wang, H.; Xu, D.-X.; Lv, J.-W.; Ning, H.; Wei, W. Melatonin attenuates lipopolysaccharide (LPS)-induced apoptotic liver damage in D-galactosamine-sensitized mice. Toxicology 2007, 237, 49–57. [Google Scholar] [CrossRef]
- Hamilton, M.K.; Boudry, G.; Lemay, D.G.; Raybould, H.E. Changes in intestinal barrier function and gut microbiota in high-fat diet-fed rats are dynamic and region dependent. Am. J. Physiol.-Gastrointest. Liver Physiol. 2015, 308, G840–G851. [Google Scholar] [CrossRef] [Green Version]
- Moreira, A.P.B.; Texeira, T.F.S.; Ferreira, A.B.; Peluzio, M.d.C.G.; Alfenas, R.d.C.G. Influence of a high-fat diet on gut microbiota, intestinal permeability and metabolic endotoxaemia. Br. J. Nutr. 2012, 108, 801–809. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, H.; Wang, J.; Liu, P.; Tan, X.; Ren, B.; Liu, Z.; Liu, X. Lycopene Supplementation Attenuates Oxidative Stress, Neuroinflammation, and Cognitive Impairment in Aged CD-1 Mice. J. Agric. Food Chem. 2018, 66, 3127–3136. [Google Scholar] [CrossRef]
- Li, M.; Xu, T.; Zhou, F.; Wang, M.; Song, H.; Xiao, X.; Lu, B. Neuroprotective effects of four phenylethanoid glycosides on H2O2-induced apoptosis on Pc12 cells via the Nrf2/ARE pathway. Int. J. Mol. Sci. 2018, 19, 1135. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Liu, X.; Luo, S.; Chu, C.; Wu, D.; Liu, R.; Lei, W.; Wang, J.; Liu, X. Extract of sesame cake and sesamol alleviate chronic unpredictable mild stress-induced depressive-like behaviors and memory deficits. J. Funct. Foods 2018, 42, 237–247. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer | |
|---|---|---|
| Fis1 | AGGCTCTAAAGTATGTGCGAGG | GGCCTTATCAATCAGGCGTTC |
| Drp1 | CGTGACAAATGAAATGGTGC | CATTAGCCCACAGGCATCAG |
| Pink1 | CTGAGATGCCTGAGTCGGTG | CTGAGATGCCTGAGTCGGTG |
| Binp3 | CTCCCAGACACCACAAGATAC | CTTCCTCAGACAGAGTGCTG |
| Mfn1 | CCTACTGCTCCTTCTAACCCA | AGGGACGCCAATCCTGTGA |
| Opa1 | CTGAGGCCCTTCTCTTGTTAGG | CTGACACCTTCCTGTAATGCTTG |
| CYP2E1 | CCTGGTGGAGGAGCTCAAAA | TGTTGAAGAGAATATCCGCAATGA |
| POR | AGGCACATCCTAGCCATTCTCCAA | ACTTCGCTTCATACTCCACAGCCA |
| ZO-1 | TGGGAACAGCACACAGTGAC | GCTGGCCCTCCTTTTAACAC |
| Occludin | ACCCGAAGAAAGATGGATCG | CATAGTCAGATGGGGGTGGA |
| Claudin-1 | CGGGCAGATACAGTGCAAAG | ACTTCATGCCAATGGTGGAC |
| Tnf-α | CCCTCACACTCAGATCATCTTCT | GCTACGACGTGGGCTACAG |
| IL-1β | TGACGGACCCCAAAAGATGA | TCTCCACAGCCACAATGAGT |
| GADPH | TGGAGAAACCTGCCAAGTATGA | TGGAAGAATGGGAGTTGCTGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Liu, Y.; Gao, H.; Ge, S.; Yao, X.; Liu, C.; Tan, X. Chronotoxicity of Acrylamide in Mice Fed a High-Fat Diet: The Involvement of Liver CYP2E1 Upregulation and Gut Leakage. Molecules 2023, 28, 5132. https://doi.org/10.3390/molecules28135132
Wang L, Liu Y, Gao H, Ge S, Yao X, Liu C, Tan X. Chronotoxicity of Acrylamide in Mice Fed a High-Fat Diet: The Involvement of Liver CYP2E1 Upregulation and Gut Leakage. Molecules. 2023; 28(13):5132. https://doi.org/10.3390/molecules28135132
Chicago/Turabian StyleWang, Luanfeng, Yanhong Liu, Huajing Gao, Shuqi Ge, Xinru Yao, Chang Liu, and Xintong Tan. 2023. "Chronotoxicity of Acrylamide in Mice Fed a High-Fat Diet: The Involvement of Liver CYP2E1 Upregulation and Gut Leakage" Molecules 28, no. 13: 5132. https://doi.org/10.3390/molecules28135132
APA StyleWang, L., Liu, Y., Gao, H., Ge, S., Yao, X., Liu, C., & Tan, X. (2023). Chronotoxicity of Acrylamide in Mice Fed a High-Fat Diet: The Involvement of Liver CYP2E1 Upregulation and Gut Leakage. Molecules, 28(13), 5132. https://doi.org/10.3390/molecules28135132