


Recent Developments and Applications of Microbial Levan, A Versatile Polysaccharide-Based Biopolymer

 , and
, and
Abstract
: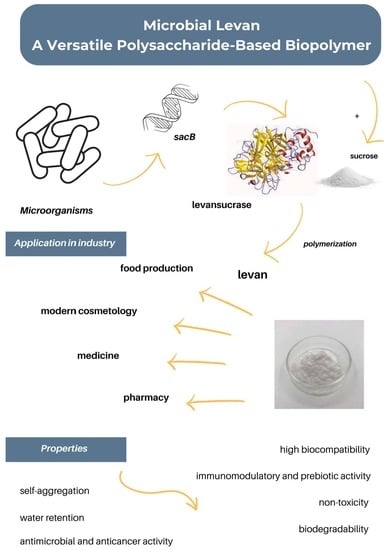
1. Introduction
2. Biosynthesis of Levan
3. Microbial Preparation of Levan

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scale | Microorganism | Sucrose Concentration [g/L] | Cultivation Conditions | Levan Concentration [g/L] | Literature |
|---|---|---|---|---|---|
| Laboratory | Leuconostoc citreum BD1707 | 172.0 | 112 h, 26 °C, 200 rpm, pH 6.12 | 34.86 | [65] |
| Tanticharoenia sakaeratensis | 200.0 | 35 h, 37 °C, 250 rpm | 24.70 | [66] | |
| B. subtilis AF17 | 162.5 | 20 h, 30 °C, 150 rpm, pH 7.0 | 7.90 | [67] | |
| B.subtilis MT453867 | 80.0 | 54 h, 37 °C, 150 rpm, pH 5.0 | 33.00 | [68] | |
| Brenneria goodwinii | 500.0 | 12 h, 35 °C, 8000 rpm, pH 6.0 | 185.00 | [50] | |
| Clostridium acetobutylicum | 28.00 | 72 h, 20 °C, - *, pH 6.0 | - * | [51] | |
| Leuconostoc mesenteroides | 90.0 | 24 h, 30 °C, - *, pH 5.0 | - * | [52] | |
| Sphingobium chungbukense | 10.00 | 24 h, 37 °C, - *, pH 5–10 | - * | [54] | |
| Gluconobacter japonicus LMG 1417 | 720.00 | 24 h, 28 °C, 180 rpm, pH 6.8 | 157.90 | [55] | |
| Peanibacillus polymyxa | 124.00 | 96 h, 30 °C, 250 rpm, pH 7.2 | 68.00 | [56] | |
| Azotobacter chroococcum DSM 2286 + Gluconobacter japonicus LMG 1417 | 809.6 | 48 h, 45 °C, - *, pH 6.2 | 387.40 | [57] | |
| Saccharomyces cereviasie | 50.00 | 48 h, 30 °C, pH 6.5 | 15.00 | [58] | |
| Bioreactor | B. subtilis M | 100.0 | 150 L bioreactor, 24 h, 30 ℃, 200 rpm, pH 7.0, 0.5 vvm | 47.00 | [69] |
| B. subtlis B58 | 130.0 | 16 L bioreactor, 16 h, 37 ℃, 400 rpm, pH 7.0, 1 vvm | 26.65 | [70] | |
| Z. mobilis B-14023 | 299.1 | 42.3 h, 28 ℃, pH 6.0 | 40.20 | [59] | |
| E. herbicola | 50.0 | 5 L bioreactor, 48 h, 25 ℃, 200 rpm, 0.2 vvm | 4.50 | [60] | |
| B. subtilis AF17 | 162.5 | 30 °C, 72 h, 150 rpm, pH 7.0 | 7.90 | [67] | |
| Halomonas smyrnensis AAD6T | 20.00 | 10 L bioreactor 37 °C, 200 rpm, 0.1 vvm, | 18.06 | [70] | |
| Bacillus amyloliquefaciens | 20.00 | 5 L bioreactor, 37 °C, 48 h, 300 rpm, pH 6.0, 6 vvm | 102.00 | [71] | |
| Pichia pastoris | 160.00 | 1,5 L bioreactor, 59 h, 28 °C, 200–1200 rpm | 72.90 | [72] | |
| Saccharomyces cerevisiae | 191.00 | 50 L bioreactor, 48 h, 30 °C, 300 rpm, pH 5.5, 0.5 vvm | 76.00 | [73] |
| Substrate/By-Product used | Microorganism | Sucrose Concentration [%] | Levan Concentration [g/L] | Literature |
|---|---|---|---|---|
| Molasses | Halomonas sp. AAD6 | 48–51 | 12.40 | [61] |
| B. licheniformis NS032 | 49.40 | 53.20 | [78] | |
| B. lentus V8 | 54 | 45.34 | [75] | |
| Sugar beet juice and syrup | Z. mobilis 113 | 65 | 9.60 | [79] |
| The pulp after squeezing orange juice | B. atrophaeus | 12–27 | 24.20 | [63] |
| Distillers’ rye stock | Z. mobilis | * | 25.17 | [80] |
| Date syrup | Microbacterium laevaniformans | * | 10.48 | [81] |
| B. phenoliresistens | * | 8.12 | [76] | |
| Sugar cane syrup | Z. mobilis | * | 15.46 | [82] |
| Baklava syrup | Z. mobilis | * | 8.90 | [83] |
| Buckwheat sourdough bread | G. albidus TMW 2.1191; Kozakia baliensis NBRC 16680 | 8.86 ± 21.71; 50.26 ± 25.21 | 13.91; 12.65 | [64] |
4. Properties and Application of Levan
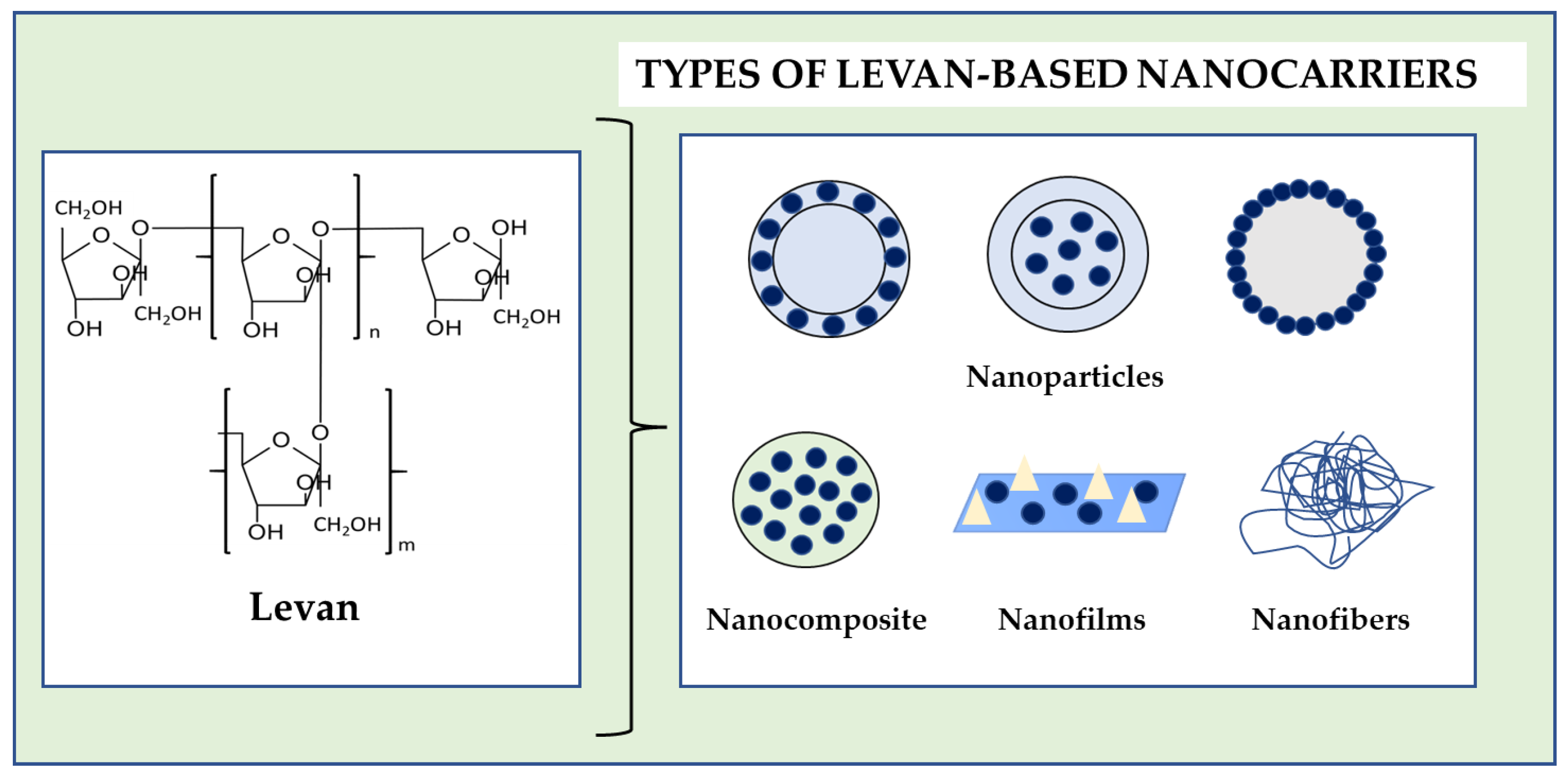
5. Levan Nanocarriers Development
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
References
- Gupta, S.; Sharma, S.; Kumar Nadda, A.; Saad Bala Husain, M.; Gupta, A. Biopolymers from Waste Biomass and Its Applications in the Cosmetic Industry: A Review. Mater. Today Proc. 2022, 68, 873–879. [Google Scholar] [CrossRef]
- Koyyada, A.; Orsu, P. Natural Gum Polysaccharides as Efficient Tissue Engineering and Drug Delivery Biopolymers. J. Drug Deliv. Sci. Technol. 2021, 63, 102431. [Google Scholar] [CrossRef]
- Kartik, A.; Akhil, D.; Lakshmi, D.; Panchamoorthy Gopinath, K.; Arun, J.; Sivaramakrishnan, R.; Pugazhendhi, A. A Critical Review on Production of Biopolymers from Algae Biomass and Their Applications. Bioresour. Technol. 2021, 329, 124868. [Google Scholar] [CrossRef] [PubMed]
- Wankhade, V. Chapter 6—Animal-Derived Biopolymers in Food and Biomedical Technology. In Biopolymer-Based Formulations: Biomedical and Food Applications; Pal, K., Banerjee, I., Sarkar, P., Kim, D., Deng, W.-P., Dubey, N.K., Majumder, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 139–152. ISBN 978-0-12-816897-4. [Google Scholar]
- Mohammed, A.S.A.; Naveed, M.; Jost, N. Polysaccharides; Classification, Chemical Properties, and Future Perspective Applications in Fields of Pharmacology and Biological Medicine (A Review of Current Applications and Upcoming Potentialities). J. Polym. Environ. 2021, 29, 2359–2371. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.-H.; Qi, C.; Ma, M.-G.; Wan, P. Multifunctional Cellulose-Based Hydrogels for Biomedical Applications. J. Mater. Chem. B 2019, 7, 1541–1562. [Google Scholar] [CrossRef]
- Li, Z.; Wei, C. Morphology, Structure, Properties and Applications of Starch Ghost: A Review. Int. J. Biol. Macromol. 2020, 163, 2084–2096. [Google Scholar] [CrossRef]
- Bilal, M.; Gul, I.; Basharat, A.; Qamar, S.A. Polysaccharides-Based Bio-Nanostructures and Their Potential Food Applications. Int. J. Biol. Macromol. 2021, 176, 540–557. [Google Scholar] [CrossRef]
- Prajapati, D.; Pal, A.; Dimkpa, C.; Harish; Singh, U.; Devi, K.A.; Choudhary, J.L.; Saharan, V. Chitosan Nanomaterials: A Prelim of next-Generation Fertilizers; Existing and Future Prospects. Carbohydr. Polym. 2022, 288, 119356. [Google Scholar] [CrossRef]
- Cadenas, R.; Caballero, I.; Nimubona, D.; Blanco, C.A. Brewing with Starchy Adjuncts: Its Influence on the Sensory and Nutritional Properties of Beer. Foods 2021, 10, 1726. [Google Scholar] [CrossRef]
- Šuchová, K.; Fehér, C.; Ravn, J.L.; Bedő, S.; Biely, P.; Geijer, C. Cellulose- and Xylan-Degrading Yeasts: Enzymes, Applications and Biotechnological Potential. Biotechnol. Adv. 2022, 59, 107981. [Google Scholar] [CrossRef]
- Ganjoo, R.; Sharma, S.; Verma, C.; Quraishi, M.A.; Kumar, A. Heteropolysaccharides in Sustainable Corrosion Inhibition: 4E (Energy, Economy, Ecology, and Effectivity) Dimensions. Int. J. Biol. Macromol. 2023, 235, 123571. [Google Scholar] [CrossRef] [PubMed]
- Salwowska, N.M.; Bebenek, K.A.; Żądło, D.A.; Wcisło-Dziadecka, D.L. Physiochemical Properties and Application of Hyaluronic Acid: A Systematic Review. J. Cosmet. Dermatol. 2016, 15, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Wickramaarachchi, K.; Sundaram, M.M.; Henry, D.J.; Gao, X. Alginate Biopolymer Effect on the Electrodeposition of Manganese Dioxide on Electrodes for Supercapacitors. ACS Appl. Energy Mater. 2021, 4, 7040–7051. [Google Scholar] [CrossRef]
- Warren, R.A. Microbial Hydrolysis of Polysaccharides. Annu. Rev. Microbiol. 1996, 50, 183–212. [Google Scholar] [CrossRef]
- Then, C.; Othman, Z.; Azha, W.; Mustapha, W.; Sarmidi, M.R.; Aziz, R.; El Enshasy, H.A. Production of Alginate by Azotobacter Vinelandii in Semi-Industrial Scale Using Batch and Fed-Batch Cultivation Systems. J. Adv. Sci. Res. 2012, 3, 45–50. [Google Scholar]
- Senni, K.; Pereira, J.; Gueniche, F.; Delbarre-Ladrat, C.; Sinquin, C.; Ratiskol, J.; Godeau, G.; Fischer, A.M.; Helley, D.; Colliec-Jouault, S. Marine Polysaccharides: A Source of Bioactive Molecules for Cell Therapy and Tissue Engineering. Mar. Drugs 2011, 9, 1664–1681. [Google Scholar] [CrossRef] [Green Version]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial Extracellular Polysaccharides Involved in Biofilm Formation. Molecules 2009, 14, 2535–2554. [Google Scholar] [CrossRef] [Green Version]
- Nwodo, U.U.; Green, E.; Okoh, A.I. Bacterial Exopolysaccharides: Functionality and Prospects. Int. J. Mol. Sci. 2012, 13, 14002–14015. [Google Scholar] [CrossRef] [Green Version]
- Prakash, U.; Singh, L.; Sharma, V. Role of Xanthan Gum (Xanthomonas Compestris) in Gastroretentive Drug Delivery System: An Overview. Int. Res. J. Pharm. 2016, 4, 35–38. [Google Scholar] [CrossRef]
- Ruissen, A.L.A.; van der Reijden, W.A.; van’t Hof, W.; Veerman, E.C.I.; Nieuw Amerongen, A. V Evaluation of the Use of Xanthan as Vehicle for Cationic Antifungal Peptides. J. Control. Release 1999, 60, 49–56. [Google Scholar] [CrossRef]
- Yuan, M.; Fu, G.; Sun, Y.; Zhang, D. Biosynthesis and Applications of Curdlan. Carbohydr. Polym. 2021, 273, 118597. [Google Scholar] [CrossRef]
- Srikanth, R.; Reddy, C.H.S.S.S.; Siddartha, G.; Ramaiah, M.J.; Uppuluri, K.B. Review on Production, Characterization and Applications of Microbial Levan. Carbohydr. Polym. 2015, 120, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Verlinden, R.A.J.; Hill, D.J.; Kenward, M.A.; Williams, C.D.; Radecka, I. Bacterial Synthesis of Biodegradable Polyhydroxyalkanoates. J. Appl. Microbiol. 2007, 102, 1437–1449. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.S.; Kaur, N.; Rana, V.; Kennedy, J.F. Pullulan: A Novel Molecule for Biomedical Applications. Carbohydr. Polym. 2017, 171, 102–121. [Google Scholar] [CrossRef]
- Greig-Smith, R.; Steel, T. Levan: A New Bacterial Gum from Sugar. J. Soc. Chem. Ind. Scottish Sect. 1902, 21, 1381. [Google Scholar]
- Gupta, S.K.; Das, P.; Singh, S.K.; Akhtar, M.S.; Meena, D.K.; Mandal, S.C. Microbial Levan, an Ideal Prebiotic and Immunonutrient in Aquaculture. World Aquac. 1990, 42, 61–64. [Google Scholar]
- Combie, J. Properties of Levan and Potential Medical Uses. Polysaccharides Drug Deliv. Pharm. Appl. 2006, 934, 13–263. [Google Scholar] [CrossRef]
- Domżał-Kędzia, M.; Lewińska, A.; Jaromin, A.; Weselski, M.; Pluskota, R.; Łukaszewicz, M. Fermentation Parameters and Conditions Affecting Levan Production and Its Potential Applications in Cosmetics. Bioorg. Chem. 2019, 93, 102787. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, B.C.M.; Baldo, C.; Celligoi, M.A.P.C. Levan and Levansucrase—A Mini Review. Int. J. Sci. Technol. Res. 2015, 4, 100–104. [Google Scholar]
- Mardo, K.; Visnapuu, T.; Gromkova, M.; Aasamets, A.; Viigand, K.; Vija, H.; Alamäe, T. High-Throughput Assay of Levansucrase Variants in Search of Feasible Catalysts for the Synthesis of Fructooligosaccharides and Levan. Molecules 2014, 19, 8434–8455. [Google Scholar] [CrossRef] [PubMed]
- González-Garcinuño, Á.; Tabernero, A.; Domínguez, Á.; Galán, M.A.; Martin del Valle, E.M. Levan and Levansucrases: Polymer, Enzyme, Micro-Organisms and Biomedical Applications. Biocatal. Biotransformation 2017, 36, 233–244. [Google Scholar] [CrossRef]
- Freitas, F.; Reis, M.A.M.; De Lisboa, U.N.; Lisboa, U. De Polysaccharides; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–24. [Google Scholar] [CrossRef]
- Leloup, L.; Driessen, A.J.M.; Freudl, R.; Chambert, R.; Petit-Glatron, M.F. Differential Dependence of Levansucrase and α-Amylase Secretion on SecA (Div) during the Exponential Phase of Growth of Bacillus Subtilis. J. Bacteriol. 1999, 181, 1820–1826. [Google Scholar] [CrossRef] [PubMed]
- Daguer, J.P.; Geissmann, T.; Petit-Glatron, M.F.; Chambert, R. Autogenous Modulation of the Bacillus Subtilis SacB-LevB-YveA Levansucrase Operon by the LevB Transcript. Microbiology 2004, 150, 3669–3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey, M.W.; Ramaiya, P.; Nelson, B.A.; Brody-Karpin, S.D.; Zaretsky, E.J.; Tang, M.; Lopez de Leon, A.; Xiang, H.; Gusti, V.; Clausen, I.G.; et al. Complete Genome Sequence of the Industrial Bacterium Bacillus Licheniformis and Comparisons with Closely Related Bacillus Species. Genome Biol. 2004, 5, r77. [Google Scholar] [CrossRef] [Green Version]
- Pereira, Y.; Petit-Glatron, M.F.; Chambert, R. YveB, Encoding Endolevanase LevB, Is Part of the SacB-YveB-YveA Levansucrase Tricistronic Operon in Bacillus Subtilis. Microbiology 2001, 147, 3413–3419. [Google Scholar] [CrossRef] [Green Version]
- Jensen, S.L.; Diemer, M.B.; Lundmark, M.; Larsen, F.H.; Blennow, A.; Mogensen, H.K.; Nielsen, T.H. Levanase from Bacillus Subtilis Hydrolyses β-2,6 Fructosyl Bonds in Bacterial Levans and in Grass Fructans. Int. J. Biol. Macromol. 2016, 85, 514–521. [Google Scholar] [CrossRef]
- Méndez-Lorenzo, L.; Porras-Domínguez, J.R.; Raga-Carbajal, E.; Olvera, C.; Rodríguez-Alegría, M.E.; Carrillo-Nava, E.; Costas, M.; Munguía, A.L. Intrinsic Levanase Activity of Bacillus Subtilis 168 Levansucrase (SacB). PLoS ONE 2015, 10, e0143394. [Google Scholar] [CrossRef] [Green Version]
- Shih, I.L.; Chen, L.D.; Wang, T.C.; Wu, J.Y.; Liaw, K.S. Tandem Production of Levan and Ethanol by Microbial Fermentation. Green Chem. 2010, 12, 1242–1247. [Google Scholar] [CrossRef]
- Marvasi, M.; Visscher, P.T.; Casillas Martinez, L. Exopolymeric Substances (EPS) from Bacillus Subtilis: Polymers and Genes Encoding Their Synthesis. FEMS Microbiol. Lett. 2010, 313, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Martin, I.; Debarbouille, M.; Klier, A.; Rapoport, G. Induction and Metabolite Regulation of Levanase Synthesis in Bacillus Subtilis. J. Bacteriol. 1989, 171, 1885–1892. [Google Scholar] [CrossRef] [Green Version]
- Martin, I.; Débarbouillé, M.; Ferrari, E.; Klier, A.; Rapoport, G. Characterization of the Levanase Gene of Bacillus Subtilis Which Shows Homology to Yeast Invertase. MGG Mol. Gen. Genet. 1987, 208, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Martin-Verstraete, I.; Débarbouillé, M.; Klier, A.; Rapoport, G. Levanase Operon of Bacillus Subtilis Includes a Fructose-Specific Phosphotransferase System Regulating the Expression of the Operon. J. Mol. Biol. 1990, 214, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Débarbouillé, M.; Martin-Verstraete, I.; Arnaud, M.; Klier, A.; Rapoport, G. Positive and Negative Regulation Controlling Expression of the Sac Genes in Bacillus Subtilis. Res. Microbiol. 1991, 142, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Wanker, E.; Huber, A.; Schwab, H. Purification and Characterization of the Bacillus Subtilis Levanase Produced in Escherichia coli. Appl. Environ. Microbiol. 1995, 61, 1953–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raga-Carbajal, E.; Carrillo-Nava, E.; Costas, M.; Porras-Dominguez, J.; López-Munguía, A.; Olvera, C. Size Product Modulation by Enzyme Concentration Reveals Two Distinct Levan Elongation Mechanisms in Bacillus Subtilis Levansucrase. Glycobiology 2016, 26, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, I.-L.; Yu, Y.-T.; Shieh, C.-J.; Hsieh, C.-Y. Selective Production and Characterization of Levan by Bacillus Subtilis (Natto) Takahashi. J. Agric. Food Chem. 2005, 53, 82118215. [Google Scholar] [CrossRef] [PubMed]
- Euzenat, O.; Guibert, A.; Combes, D. Production of Fructo-Oligosaccharides by Levansucrase from Bacillus Subtilis C4. Process Biochem. 1997, 32, 237–243. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, S.; Zhang, T.; Jiang, B.; Mu, W. Efficient Biosynthesis of Levan from Sucrose by a Novel Levansucrase from Brenneria Goodwinii. Carbohydr. Polym. 2017, 157, 1732–1740. [Google Scholar] [CrossRef]
- Gao, S.; Qi, X.; Hart, D.J.; Gao, H.; An, Y. Expression and Characterization of Levansucrase from Clostridium Acetobutylicum. J. Agric. Food Chem. 2017, 65, 867–871. [Google Scholar] [CrossRef]
- Ishida, R.; Sakaguchi, K.; Matsuzaki, C.; Katoh, T.; Ishida, N.; Yamamoto, K.; Hisa, K. Levansucrase from Leuconostoc Mesenteroides NTM048 Produces a Levan Exopolysaccharide with Immunomodulating Activity. Biotechnol. Lett. 2016, 38, 681–687. [Google Scholar] [CrossRef]
- Kirtel, O.; Menéndez, C.; Versluys, M.; Van den Ende, W.; Hernández, L.; Toksoy Öner, E. Levansucrase from Halomonas Smyrnensis AAD6T: First Halophilic GH-J Clan Enzyme Recombinantly Expressed, Purified, and Characterized. Appl. Microbiol. Biotechnol. 2018, 102, 9207–9220. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Shin, W.-R.; Sekhon, S.S.; Lee, J.-P.; Kim, Y.-C.; Ahn, J.-Y.; Kim, Y.-H. Molecular Docking Analysis and Biochemical Evaluation of Levansucrase from Sphingobium Chungbukense DJ77. ACS Comb. Sci. 2018, 20, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Hövels, M.; Kosciow, K.; Kniewel, J.; Jakob, F.; Deppenmeier, U. High Yield Production of Levan-Type Fructans by Gluconobacter Japonicus LMG 1417. Int. J. Biol. Macromol. 2020, 164, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Liyaskina, E.V.; Rakova, N.A.; Kitykina, A.A.; Rusyaeva, V.V.; Toukach, P.V.; Fomenkov, A.; Vainauskas, S.; Roberts, R.J.; Revin, V.V. Production and Characterization of the Exopolysaccharide from Strain Paenibacillus Polymyxa 2020. PLoS ONE 2021, 16, e0253482. [Google Scholar] [CrossRef]
- Hövels, M.; Kosciow, K.; Deppenmeier, U. Characterization of a Novel Endo-Levanase from Azotobacter Chroococcum DSM 2286 and Its Application for the Production of Prebiotic Fructooligosaccharides. Carbohydr. Polym. 2021, 255, 117384. [Google Scholar] [CrossRef]
- Franken, J.; Brandt, B.A.; Tai, S.L.; Bauer, F.F. Biosynthesis of Levan, a Bacterial Extracellular Polysaccharide, in the Yeast Saccharomyces Cerevisiae. PLoS ONE 2013, 8, e77499. [Google Scholar] [CrossRef] [Green Version]
- Silbir, S.; Dagbagli, S.; Yegin, S.; Baysal, T.; Goksungur, Y. Levan Production by Zymomonas Mobilis in Batch and Continuous Fermentation Systems. Carbohydr. Polym. 2014, 99, 454–461. [Google Scholar] [CrossRef]
- Keith, J.; Wiley, B.; Ball, D.; Arcidiacono, S.; Zorfass, D.; Mayer, J.; Kaplan, D. Continuous Culture System for Production of Biopolymer Levan Using Erwinia Herbicola. Biotechnol. Bioeng. 1991, 38, 557–560. [Google Scholar] [CrossRef]
- Küçükaşik, F.; Kazak, H.; Güney, D.; Finore, I.; Poli, A.; Yenigün, O.; Nicolaus, B.; Öner, E.T. Molasses as Fermentation Substrate for Levan Production by Halomonas sp. Appl. Microbiol. Biotechnol. 2011, 89, 1729–1740. [Google Scholar] [CrossRef]
- Seibel, J.; Moraru, R.; Götze, S.; Buchholz, K.; Na’amnieh, S.; Pawlowski, A.; Hecht, H.J.; Secchi, M.; Castellani, V.; Collina, E.; et al. Selenium Enriched Green Tea Increase Stability of Lactobacillus Casei and Lactobacillus Plantarum in Chitosan Coated Alginate Microcapsules during Exposure to Simulated Gastrointestinal and Refrigerated Conditions. Int. J. Biol. Macromol. 2020, 21, 406–411. [Google Scholar] [CrossRef]
- González-Garcinuño, Á.; Tabernero, A.; Sánchez-Álvarez, J.M.; Galán, M.A.; Martin del Valle, E.M. Effect of Bacteria Type and Sucrose Concentration on Levan Yield and Its Molecular Weight. Microb. Cell Fact. 2017, 16, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ua-Arak, T.; Jakob, F.; Vogel, R.F. Influence of Levan-Producing Acetic Acid Bacteria on Buckwheat-Sourdough Breads. Food Microbiol. 2017, 65, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Feng, H.; Wang, X.; Liu, Z.; Wu, Z. Levan from Leuconostoc Citreum BD1707: Production Optimization and Changes in Molecular Weight Distribution during Cultivation. BMC Biotechnol. 2021, 21, 14. [Google Scholar] [CrossRef]
- Aramsangtienchai, P.; Kongmon, T.; Pechroj, S.; Srisook, K. Enhanced Production and Immunomodulatory Activity of Levan from the Acetic Acid Bacterium, Tanticharoenia Sakaeratensis. Int. J. Biol. Macromol. 2020, 163, 574–581. [Google Scholar] [CrossRef]
- Bouallegue, A.; Casillo, A.; Chaari, F.; Cimini, D.; Corsaro, M.M.; Bachoual, R.; Ellouz-Chaabouni, S. Statistical Optimization of Levan: Influence of the Parameter on Levan Structure and Angiotensin I-Converting Enzyme Inhibitory. Int. J. Biol. Macromol. 2020, 158, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Gamal, A.A.; Abbas, H.Y.; Abdelwahed, N.A.M.; Kashef, M.T.; Mahmoud, K.; Esawy, M.A.; Ramadan, M.A. Optimization Strategy of Bacillus Subtilis MT453867 Levansucrase and Evaluation of Levan Role in Pancreatic Cancer Treatment. Int. J. Biol. Macromol. 2021, 182, 1590–1601. [Google Scholar] [CrossRef] [PubMed]
- Ragab, T.I.M.; Malek, R.A.; Elsehemy, I.A.; Farag, M.M.S.; Salama, B.M.; Abd EL-Baseer, M.A.; Gamal-Eldeen, A.M.; El Enshasy, H.A.; Esawy, M.A. Scaling up of Levan Yield in Bacillus Subtilis M and Cytotoxicity Study on Levan and Its Derivatives. J. Biosci. Bioeng. 2019, 127, 655–662. [Google Scholar] [CrossRef]
- Hamid, K.R.A.; Elsayed, E.A.; Enshasy, H.A.E.; Esawy, M.; Malek, R.A. Bioprocess Optimization for Levan Production by Bacillus Subtilis B58. J. Sci. Ind. Res. (India) 2018, 77, 386–393. [Google Scholar]
- Gu, Y.; Zheng, J.; Feng, J.; Cao, M.; Gao, W.; Quan, Y.; Dang, Y.; Wang, Y.; Wang, S.; Song, C. Improvement of Levan Production in Bacillus Amyloliquefaciens through Metabolic Optimization of Regulatory Elements. Appl. Microbiol. Biotechnol. 2017, 101, 4163–4174. [Google Scholar] [CrossRef]
- Ávila-Fernández, Á.; Montiel, S.; Rodríguez-Alegría, M.E.; Caspeta, L.; López Munguía, A. Simultaneous Enzyme Production, Levan-Type FOS Synthesis and Sugar by-Products Elimination Using a Recombinant Pichia Pastoris Strain Expressing a Levansucrase-Endolevanase Fusion Enzyme. Microb. Cell Fact. 2023, 22, 18. [Google Scholar] [CrossRef]
- Ko, H.; Bae, J.H.; Sung, B.H.; Kim, M.J.; Kim, C.H.; Oh, B.R.; Sohn, J.H. Efficient Production of Levan Using a Recombinant Yeast Saccharomyces Cerevisiae Hypersecreting a Bacterial Levansucrase. J. Ind. Microbiol. Biotechnol. 2019, 46, 1611–1620. [Google Scholar] [CrossRef] [PubMed]
- Sarilmiser, H.K.; Ates, O.; Ozdemir, G.; Arga, K.Y.; Oner, E.T. Effective Stimulating Factors for Microbial Levan Production by Halomonas Smyrnensis AAD6T. J. Biosci. Bioeng. 2015, 119, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Abou-taleb, K.A.; Abdel-Monem, M.O.; Yassin, M.H.; Draz, A.A.; Blaiotta, G. Production, Purification and Characterization of Levan Polymer from Bacillus Lentus V8 Strain. Br. Microbiol. Res. J. 2015, 5, 22–32. [Google Scholar] [CrossRef]
- Moussa, T.A.A.; Al-Qaysi, S.A.S.; Thabit, Z.A.; Kadhem, S.B. Microbial Levan from Brachybacterium Phenoliresistens: Characterization and Enhancement of Production. Process Biochem. 2017, 57, 9–15. [Google Scholar] [CrossRef]
- Erkorkmaz, B.A.; Kırtel, O.; Ateş Duru, Ö.; Toksoy Öner, E. Development of a Cost-Effective Production Process for Halomonas Levan. Bioprocess Biosyst. Eng. 2018, 41, 1247–1259. [Google Scholar] [CrossRef]
- Gojgic-Cvijovic, G.D.; Jakovljevic, D.M.; Loncarevic, B.D.; Todorovic, N.M.; Pergal, M.V.; Ciric, J.; Loos, K.; Beskoski, V.P.; Vrvic, M.M. Production of Levan by Bacillus Licheniformis NS032 in Sugar Beet Molasses-Based Medium. Int. J. Biol. Macromol. 2019, 121, 142–151. [Google Scholar] [CrossRef] [Green Version]
- Bekers, M.; Linde, R.; Danilevich, A.; Kaminska, E.; Upite, D.; Vigants, A.; Scherbaka, R. Sugar Beet Diffusion Juice and Syrup as Media for Ethanol and Levan Production by Zymomonas Mobilis. Food Biotechnol. 1999, 13, 107–119. [Google Scholar] [CrossRef]
- Bekers, M.; Grube, M.; Vulfa, L.; Upite, D.; Kaminska, E.; Scherbaka, R.; Vigants, A.; Danilevich, A. Stillage as a Source of Growth Promoting Biofactors and a Stimulator of Levan and Extracellular Levansucrase Synthesis for Zymomonas Mobilis. Food Technol. Biotechnol. 2002, 40, 305–310. [Google Scholar]
- Bekers, M.; Grube, M.; Vulfa, L.; Upite, D.; Kaminska, E.E.; Scherbaka, R.; Vigants, A.; Danilevich, A.; de Oliveira, M.R.; da Silva, R.S.S.F.; et al. Microbial Production of Levan Using Date Syrup and Investigation of Its Properties. Int. J. Nutr. Food Eng. World Acad. Sci. Eng. Technol. 2007, 13, 107–119. [Google Scholar]
- de Oliveira, M.R.; da Silva, R.S.S.F.; Buzato, J.B.; Celligoi, M.A.P.C. Study of Levan Production by Zymomonas Mobilis Using Regional Low-Cost Carbohydrate Sources. Biochem. Eng. J. 2007, 37, 177–183. [Google Scholar] [CrossRef]
- Omeroglu, M.A.; Gonul-Baltaci, N.; Arslan, N.P.; Adiguzel, A.; Taskin, M. Microbial Conversion of Waste Baklava Syrup to Biofuels and Bioproducts. Biocatal. Agric. Biotechnol. 2022, 42, 102364. [Google Scholar] [CrossRef]
- Mummaleti, G.; Sarma, C.; Kalakandan, S.K.; Gazula, H.; Sivanandham, V.; Anandharaj, A. Characterization of Levan Produced from Coconut Inflorescence Sap Using Bacillus Subtilis and Its Application as a Sweetener. LWT 2022, 154, 112697. [Google Scholar] [CrossRef]
- Bersaneti, G.T.; Pan, N.C.; Baldo, C.; Celligoi, M.A.P.C. Co-Production of Fructooligosaccharides and Levan by Levansucrase from Bacillus Subtilis Natto with Potential Application in the Food Industry. Appl. Biochem. Biotechnol. 2018, 184, 838–851. [Google Scholar] [CrossRef]
- de Paula, V.C.; Pinheiro, I.O.; Lopes, C.E.; Calazans, G.M.T. Microwave-Assisted Hydrolysis of Zymomonas Mobilis Levan Envisaging Oligofructan Production. Bioresour. Technol. 2008, 99, 2466–2470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, W.; Ni, D.; Dai, Q.; Guang, C.; Zhang, T.; Mu, W. An Overview of Levan-Degrading Enzyme from Microbes. Appl. Microbiol. Biotechnol. 2019, 103, 7891–7902. [Google Scholar] [CrossRef]
- Cai, G.; Wu, D.; Li, X.; Lu, J. Levan from Bacillus Amyloliquefaciens JN4 Acts as a Prebiotic for Enhancing the Intestinal Adhesion Capacity of Lactobacillus Reuteri JN101. Int. J. Biol. Macromol. 2020, 146, 482–487. [Google Scholar] [CrossRef]
- Cheng, R.; Cheng, L.; Zhao, Y.; Wang, L.; Wang, S.; Zhang, J. Biosynthesis and Prebiotic Activity of a Linear Levan from a New Paenibacillus Isolate. Appl. Microbiol. Biotechnol. 2021, 105, 769–787. [Google Scholar] [CrossRef]
- Shimizu, N.; Abea, A.; Ushiyama, T.; Toksoy Öner, E. Effect of Temperature on the Hydrolysis of Levan Treated with Compressed Hot Water Fluids. Food Sci. Nutr. 2020, 8, 2004–2014. [Google Scholar] [CrossRef]
- Xu, M.; Pan, L.; Zhou, Z.; Han, Y. Structural Characterization of Levan Synthesized by a Recombinant Levansucrase and Its Application as Yogurt Stabilizers. Carbohydr. Polym. 2022, 291, 119519. [Google Scholar] [CrossRef]
- Kovanda, L.; Zhang, W.; Wei, X.; Luo, J.; Wu, X.; Atwill, E.R.; Vaessen, S.; Li, X.; Liu, Y. In Vitro Antimicrobial Activities of Organic Acids and Their Derivatives on Several Species of Gram-Negative and Gram-Positive Bacteria. Molecules 2019, 24, 3770. [Google Scholar] [CrossRef] [Green Version]
- Gan, L.; Jiang, G.; Yang, Y.; Zheng, B.; Zhang, S.; Li, X.; Tian, Y.; Peng, B. Development and Characterization of Levan/Pullulan/Chitosan Edible Films Enriched with ε-Polylysine for Active Food Packaging. Food Chem. 2022, 388, 132989. [Google Scholar] [CrossRef] [PubMed]
- Koşarsoy Ağçeli, G. A New Approach to Nanocomposite Carbohydrate Polymer Films: Levan and Chia Seed Mucilage. Int. J. Biol. Macromol. 2022, 218, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.H.; Chung, C.B.; Kim, Y.H.; Kim, K.S.H.; Han, C.S.; Kim, C.H. Cosmeceutical Properties of Levan Produced by Zymomonas Mobilis. Int. J. Cosmet. Sci. 2006, 28, 231. [Google Scholar] [CrossRef]
- Lewińska, A.; Domżał-Kędzia, M.; Kierul, K.; Bochynek, M.; Pannert, D.; Nowaczyk, P.; Łukaszewicz, M. Targeted Hybrid Nanocarriers as a System Enhancing the Skin Structure. Molecules 2021, 26, 1063. [Google Scholar] [CrossRef]
- Kim, T.; Kim, K.; Ryo, K.; Lee, O.; Kim, T. Cosmetic Composition Containing Levan Having Cell Proliferation, Skin-Moisturizing and Irritation-Alleviating Effects. Patent JP2003277225A, 2 October 2003. [Google Scholar]
- Masayo, F.; Takayuki, T. Beautifully Whitening Agent 2006. Patent JP2006052146A, 23 February 2006. [Google Scholar]
- Pantelić, I.; Lukić, M.; Gojgić-Cvijović, G.; Jakovljević, D.; Nikolić, I.; Lunter, D.J.; Daniels, R.; Savić, S. Bacillus Licheniformis Levan as a Functional Biopolymer in Topical Drug Dosage Forms: From Basic Colloidal Considerations to Actual Pharmaceutical Application. Eur. J. Pharm. Sci. 2020, 142, 105109. [Google Scholar] [CrossRef]
- Sezer, A.D.; Kazak Sarılmışer, H.; Rayaman, E.; Çevikbaş, A.; Öner, E.T.; Akbuğa, J. Development and Characterization of Vancomycin-Loaded Levan-Based Microparticular System for Drug Delivery. Pharm. Dev. Technol. 2017, 22, 627–634. [Google Scholar] [CrossRef]
- Cinan, E.; Cesur, S.; Erginer Haskoylu, M.; Gunduz, O.; Toksoy Oner, E. Resveratrol-Loaded Levan Nanoparticles Produced by Electrohydrodynamic Atomization Technique. Nanomaterials 2021, 11, 2582. [Google Scholar] [CrossRef]
- Kim, S.J.; Bae, P.K.; Chung, B.H. Self-Assembled Levan Nanoparticles for Targeted Breast Cancer Imaging. Chem. Commun. 2015, 51, 107–110. [Google Scholar] [CrossRef]
- Richa, R.; Roy Choudhury, A. Exploration of Polysaccharide Based Nanoemulsions for Stabilization and Entrapment of Curcumin. Int. J. Biol. Macromol. 2020, 156, 1287–1296. [Google Scholar] [CrossRef]
- Patel, P.; Agrawal, Y. Preparation and In-Vitro Evaluation of Levan Micelles: A Polyfructan Based Nano-Carrier for Breast Cancer Targeted Delivery. Drug Deliv. Lett. 2019, 9, 97–107. [Google Scholar] [CrossRef]
- Xu, W.; Peng, J.; Ni, D.; Zhang, W.; Wu, H.; Mu, W. Preparation, Characterization and Application of Levan/Montmorillonite Biocomposite and Levan/BSA Nanoparticle. Carbohydr. Polym. 2020, 234, 115921. [Google Scholar] [CrossRef] [PubMed]
- Demirci, T.; Hasköylü, M.E.; Eroğlu, M.S.; Hemberger, J.; Toksoy Öner, E. Levan-Based Hydrogels for Controlled Release of Amphotericin B for Dermal Local Antifungal Therapy of Candidiasis. Eur. J. Pharm. Sci. 2020, 145, 105255. [Google Scholar] [CrossRef] [PubMed]
- Hamada, M.A.; Hassan, R.A.; Abdou, A.M.; Elsaba, Y.M.; Aloufi, A.S.; Sonbol, H.; Korany, S.M. Bio_Fabricated Levan Polymer from Bacillus Subtilis MZ292983.1 with Antibacterial, Antibiofilm, and Burn Healing Properties. Appl. Sci. 2022, 12, 6413. [Google Scholar] [CrossRef]
- Osman, A.; Lin, E.; Hwang, D.S. A Sticky Carbohydrate Meets a Mussel Adhesive: Catechol-Conjugated Levan for Hemostatic and Wound Healing Applications. Carbohydr. Polym. 2023, 299, 120172. [Google Scholar] [CrossRef]
- Hundschell, C.S.; Jakob, F.; Wagemans, A.M. Molecular Weight Dependent Structure and Polymer Density of the Exopolysaccharide Levan. arXiv 2019, arXiv:1909.07737. [Google Scholar]
- de Siqueira, E.C.; Rebouças, J.d.S.; Pinheiro, I.O.; Formiga, F.R. Levan-Based Nanostructured Systems: An Overview. Int. J. Pharm. 2020, 580, 119242. [Google Scholar] [CrossRef]
- Makino Tadashi, I.; Masayuki Yamada, K.; Jun-ichi, K. Fast Dissolving Tablet and Production. Patent US005501861A, 5 March 1996. [Google Scholar]
- Kang, S.A.; Jang, K.-H.; Seo, J.-W.; Kim, K.H.; Kim, Y.H.; Rairakhwada, D.; Seo, M.Y.; Lee, J.O.; Do Ha, S.; Kim, C.-H.; et al. Levan: Applications and Perspectives. In Microbial Production of Biopolymers and Polymer Precursors; Caister Academic Press: Norwich, UK, 2009; pp. 145–161. [Google Scholar] [CrossRef]
- Elvassore, N.; Bertucco, A.; Caliceti, P. Production of Insulin-Loaded Poly(Ethylene Glycol)/Poly(L-Lactide) (PEG/PLA) Nanoparticles by Gas Antisolvent Techniques. J. Pharm. Sci. 2001, 90, 1628–1636. [Google Scholar] [CrossRef] [PubMed]
- Bahadori, F.; Eskandari, Z.; Ebrahimi, N.; Bostan, M.S.; Eroğlu, M.S.; Oner, E.T. Development and Optimization of a Novel PLGA-Levan Based Drug Delivery System for Curcumin, Using a Quality-by-Design Approach. Eur. J. Pharm. Sci. 2019, 138, 105037. [Google Scholar] [CrossRef] [PubMed]
- González-Garcinuño, Á.; Tabernero, A.; Marcelo, G.; Sebastián, V.; Arruebo, M.; Santamaría, J.; Martín del Valle, E. Differences in Levan Nanoparticles Depending on Their Synthesis Route: Microbial vs Cell-Free Systems. Int. J. Biol. Macromol. 2019, 137, 62–68. [Google Scholar] [CrossRef]
- Charoenwongpaiboon, T.; Wangpaiboon, K.; Septham, P.; Jiamvoraphong, N.; Issaragrisil, S.; Pichyangkura, R.; Lorthongpanich, C. Production and Bioactivities of Nanoparticulated and Ultrasonic-Degraded Levan Generated by Erwinia Tasmaniensis Levansucrase in Human Osteosarcoma Cells. Int. J. Biol. Macromol. 2022, 221, 1121–1129. [Google Scholar] [CrossRef]
- Sima, F.; Mutlu, E.C.; Eroglu, M.S.; Sima, L.E.; Serban, N.; Ristoscu, C.; Petrescu, S.M.; Oner, E.T.; Mihailescu, I.N. Levan Nanostructured Thin Films by MAPLE Assembling. Biomacromolecules 2011, 12, 2251–2256. [Google Scholar] [CrossRef] [PubMed]
- Bostan, M.S.; Mutlu, E.C.; Kazak, H.; Sinan Keskin, S.; Oner, E.T.; Eroglu, M.S. Comprehensive Characterization of Chitosan/PEO/Levan Ternary Blend Films. Carbohydr. Polym. 2014, 102, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Hasirci, V.; Vrana, E.; Zorlutuna, P.; Ndreu, A.; Yilgor, P.; Basmanav, F.B.; Aydin, E. Nanobiomaterials: A Review of the Existing Science and Technology, and New Approaches. J. Biomater. Sci. Polym. Ed. 2006, 17, 1241–1268. [Google Scholar] [CrossRef] [PubMed]
- Koşarsoy Ağçeli, G.; Hammamchi, H.; Cihangir, N. Novel Levan/Bentonite/Essential Oil Films: Characterization and Antimicrobial Activity. J. Food Sci. Technol. 2022, 59, 249–256. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, Z.; Han, Y. Levan-Chitosan Blend Films: Preparation, Structural, Physical Properties and Application in Pork Packaging. Int. J. Biol. Macromol. 2022, 217, 624–632. [Google Scholar] [CrossRef]
- Song, Y.; Joo, K.; Seo, J.H. Evaluation of Mechanical and Thermal Properties of Hydroxyapatite-Levan Composite Bone Graft. Biotechnol. Bioprocess Eng. 2021, 26, 201–207. [Google Scholar] [CrossRef]
- Mujtaba, M.; Ali, Q.; Yilmaz, B.A.; Seckin Kurubas, M.; Ustun, H.; Erkan, M.; Kaya, M.; Cicek, M.; Oner, E.T. Understanding the Effects of Chitosan, Chia Mucilage, Levan Based Composite Coatings on the Shelf Life of Sweet Cherry. Food Chem. 2023, 416, 135816. [Google Scholar] [CrossRef]
- Choi, W.I.; Hwang, Y.; Sahu, A.; Min, K.; Sung, D.; Tae, G.; Chang, J.H. An Injectable and Physical Levan-Based Hydrogel as a Dermal Filler for Soft Tissue Augmentation. Biomater. Sci. 2018, 6, 2627–2638. [Google Scholar] [CrossRef]
- Hwang, Y.; Lee, J.S.; An, H.; Oh, H.; Sung, D.; Tae, G.; Choi, W. Il Hydroxyapatite-Embedded Levan Composite Hydrogel as an Injectable Dermal Filler for Considerable Enhancement of Biological Efficacy. J. Ind. Eng. Chem. 2021, 104, 491–499. [Google Scholar] [CrossRef]
- Selvi, S.S.; Hasköylü, M.E.; Genç, S.; Toksoy Öner, E. Synthesis and Characterization of Levan Hydrogels and Their Use for Resveratrol Release. J. Bioact. Compat. Polym. 2021, 36, 464–480. [Google Scholar] [CrossRef]
- Sezer, A.D.; Kazak, H.; Öner, E.T.; Akbua, J. Levan-Based Nanocarrier System for Peptide and Protein Drug Delivery: Optimization and Influence of Experimental Parameters on the Nanoparticle Characteristics. Carbohydr. Polym. 2011, 84, 358–363. [Google Scholar] [CrossRef]
- Nakapong, S.; Pichyangkura, R.; Ito, K.; Iizuka, M.; Pongsawasdi, P. High Expression Level of Levansucrase from Bacillus Licheniformis RN-01 and Synthesis of Levan Nanoparticles. Int. J. Biol. Macromol. 2013, 54, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.B.A.; Kalla, D.; Uppuluri, K.B.; Anbazhagan, V. Green Synthesis of Silver and Gold Nanoparticles Employing Levan, a Biopolymer from Acetobacter Xylinum NCIM 2526, as a Reducing Agent and Capping Agent. Carbohydr. Polym. 2014, 112, 539–545. [Google Scholar] [CrossRef]
- Kim, S.J.; Chung, B.H. Antioxidant Activity of Levan Coated Cerium Oxide Nanoparticles. Carbohydr. Polym. 2016, 150, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Akturk, O. The Anticancer Activity of Doxorubicin-Loaded Levan-Functionalized Gold Nanoparticles Synthesized by Laser Ablation. Int. J. Biol. Macromol. 2022, 196, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yue, X.; Zeng, Y.; Hua, E.; Wang, M.; Sun, Y. Bacillus Amyloliquefaciens Levan and Its Silver Nanoparticles with Antimicrobial Properties. Biotechnol. Biotechnol. Equip. 2018, 32, 1583–1589. [Google Scholar] [CrossRef] [Green Version]
- Oktavia, I.; Fithriah, A.; Permatasari, N.; Ratnaningsih, E.; Hertadi, R. Levan Produced by the Halophilic Bacterium Bacillus Licheniformis BK1 as a Nanoparticle for Protein Immobilization. Indones. J. Chem. 2020, 20, 493. [Google Scholar] [CrossRef]
- Ko, H.; Sung, B.H.; Kim, M.J.; Park, H.J.; Sohn, J.H.; Bae, J.H. Dual-Functional Carboxymethyl Levan-Based Protein Carrier for Cosmeceutical Application of Human Epidermal Growth Factor. Int. J. Biol. Macromol. 2023, 229, 181–187. [Google Scholar] [CrossRef]
- Mutlu, E.C.; Bahadori, F.; Bostan, M.S.; Sarilmiser, H.K.; ToksoyOner, E.; Eroğlu, M.S. Halomonas Levan-Coated Phospholipid Based Nano-Carrier for Active Targeting of A549 Lung Cancer Cells. Eur. Polym. J. 2021, 144, 110239. [Google Scholar] [CrossRef]
- Lee, J.S.; Park, E.; Oh, H.; Choi, W.I.; Koo, H. Levan Nanoparticles with Intrinsic CD44-Targeting Ability for Tumor-Targeted Drug Delivery. Int. J. Biol. Macromol. 2023, 234, 123634. [Google Scholar] [CrossRef]
- Lewińska, A.; Domżał-Kędzia, M.; Łukaszewicz, M. Self-Assembly Levan, as a Natural Active Bio-Nanocarrier. Sustain. Chem. Pharm. 2023, 31, 100936. [Google Scholar] [CrossRef]






| Microbial Polymer | Synthesizing Microorganisms | Functions in Cell | Application in Industries | Literature |
|---|---|---|---|---|
| Alginate | Azotobacter vinelandii, Pseudomonas aeruginosa | Protection against desiccation, starvation, phagocytosis, UV radiation, effective barrier against reactive biocides, and involvement in the formation of a biofilm on a solid substrate. Under adverse conditions caused by changes in the ionic, osmotic, pH environment or toxic–metallic conditions, exopolysaccharides can create a protective buffer zone around the cell, protecting it from unfavorable changes in its structure. Increased ability of microorganisms to colonize host tissues. | For the production of dressings for hard-to-heal wounds, it is used for encapsulation, i.e., coating (immobilization) of various bioactive materials, as a component of face creams, In the food industry, this compound is used as a thickener, stabilizer or gelling agent in the production of jellies. | [16] |
| Bacterial cellulose | Acetobacter spp. | Cellulose dressings used to treat hard-to-heal wounds, e.g., burns. | [17] | |
| Dextran | L. mesenteroides, L. dextranicum | In the food industry, it is used as a thickener and emulsifier, and in cosmetics, it is used as an ingredient in powders and lipsticks. However, the most important is dextran in medicine, where it can be used as a blood plasma substitute. | [18] | |
| Gellan | Sphingomonas elodea | Stabilizing, thickening and gelling agent (E418). Due to its thermostability, gellan can be used as an alternative to microbial agar, in particular in the cultivation of thermophilic bacteria, and as a drug carrier. | [19] | |
| Xanthan | Xanthomonas campestris | Stabilizing, thickening and gelling. Xanthan is used, e.g., for the production of puddings, sauces, drinks, fillings, and yoghurts, as well as in ointments, eye gels and toothpaste, and as an auxiliary substance in tablets, regulating the kinetics of the active substance release, e.g., by using the phenomenon of mucoadhesion—adhesion of the drug form to the membrane mucosa | [20,21] | |
| Curdlan | Alcaligenes faecalis | Used primarily in the pharmaceutical and food industries, e.g., as a stabilizer, texture modifier and water binder. | [22] | |
| Levan | Bacillus spp., Lactobacillus spp., Pseudomonas spp., Streptococcus spp., Xanthomonas spp., Zymomonas spp. | Stabilizer, flavor and fragrance carrier and thickener, and in the case of pharmaceuticals it can be used as a carrier of the active substance. In cosmetology, it is used for moisturizing preparations. | [23] | |
| PHA | Aeromonas hydrophila, Burkholderia sacchari, Escherichia coli, Halomonas baliviensis | For the encapsulation of grains and fertilizers and the production of biodegradable containers and films, as sutures, orthopedic implants, mechanical barriers to prevent post-operative adhesions, internal drug release systems in the form of stents, scaffolding for bone marrow and bandages to support wound healing. | [24] | |
| Pullulan | Aureobasidium pullulans | The production of edible films that are used in various breath fresheners or oral hygiene products, as a vegetarian substitute for medicine capsules, instead of gelatin, as a food additive is known under the number E 1204. | [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domżał-Kędzia, M.; Ostrowska, M.; Lewińska, A.; Łukaszewicz, M. Recent Developments and Applications of Microbial Levan, A Versatile Polysaccharide-Based Biopolymer. Molecules 2023, 28, 5407. https://doi.org/10.3390/molecules28145407
Domżał-Kędzia M, Ostrowska M, Lewińska A, Łukaszewicz M. Recent Developments and Applications of Microbial Levan, A Versatile Polysaccharide-Based Biopolymer. Molecules. 2023; 28(14):5407. https://doi.org/10.3390/molecules28145407
Chicago/Turabian StyleDomżał-Kędzia, Marta, Monika Ostrowska, Agnieszka Lewińska, and Marcin Łukaszewicz. 2023. "Recent Developments and Applications of Microbial Levan, A Versatile Polysaccharide-Based Biopolymer" Molecules 28, no. 14: 5407. https://doi.org/10.3390/molecules28145407
APA StyleDomżał-Kędzia, M., Ostrowska, M., Lewińska, A., & Łukaszewicz, M. (2023). Recent Developments and Applications of Microbial Levan, A Versatile Polysaccharide-Based Biopolymer. Molecules, 28(14), 5407. https://doi.org/10.3390/molecules28145407