
Oligosaccharides as Potential Therapeutics against Atherosclerosis
 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Alginate Oligosaccharide
3. Chitosan Oligosaccharide
4. Heparin-Derived Oligosaccharides
5. Galacto-Oligosaccharides
6. Cyclodextrins
6.1. Effects on Lesion and Plaque Size and Cholesterol Transport
6.2. LDL Oxidation
6.3. Adhesion to Endothelial Cells
6.4. Complement Activation
7. Oligosaccharides Which Stimulate Atherosclerosis
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fan, J.; Watanabe, T. Atherosclerosis: Known and Unknown. Pathol. Int. 2022, 72, 151–160. [Google Scholar] [CrossRef]
- WHO. CVDs Fact Sheets Cardiovascular Diseases (CVDs). Available online: https://www.who.int/health-topics/cardiovascular-diseases#tab=tab_1 (accessed on 11 June 2023).
- Borén, J.; Chapman, M.J.; Krauss, R.M.; Packard, C.J.; Bentzon, J.F.; Binder, C.J.; Daemen, M.J.; Demer, L.L.; Hegele, R.A.; Nicholls, S.J.; et al. Low-Density Lipoproteins Cause Atherosclerotic Cardiovascular Disease: Pathophysiological, Genetic, and Therapeutic Insights: A Consensus Statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2020, 41, 2313–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezentsev, A.; Bezsonov, E.; Kashirskikh, D.; Baig, M.S.; Eid, A.H.; Orekhov, A. Proatherogenic Sialidases and Desialylated Lipoproteins: 35 Years of Research and Current State from Bench to Bedside. Biomedicines 2021, 9, 600. [Google Scholar] [CrossRef] [PubMed]
- Gisterå, A.; Hansson, G.K. The Immunology of Atherosclerosis. Nat. Rev. Nephrol. 2017, 13, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Batty, M.; Bennett, M.R.; Yu, E. The Role of Oxidative Stress in Atherosclerosis. Cells 2022, 11, 3843. [Google Scholar] [CrossRef]
- Petrucci, G.; Rizzi, A.; Hatem, D.; Tosti, G.; Rocca, B.; Pitocco, D. Role of Oxidative Stress in the Pathogenesis of Atherothrombotic Diseases. Antioxidants 2022, 11, 1408. [Google Scholar] [CrossRef] [PubMed]
- Yurdagul, A.; Doran, A.C.; Cai, B.; Fredman, G.; Tabas, I.A. Mechanisms and Consequences of Defective Efferocytosis in Atherosclerosis. Front. Cardiovasc. Med. 2018, 4, 86. [Google Scholar] [CrossRef] [Green Version]
- Leszczynska, A.; O’Doherty, A.; Farrell, E.; Pindjakova, J.; O’Brien, F.J.; O’Brien, T.; Barry, F.; Murphy, M. Differentiation of Vascular Stem Cells Contributes to Ectopic Calcification of Atherosclerotic Plaque. Stem Cells 2016, 34, 913–923. [Google Scholar] [CrossRef] [Green Version]
- Hutcheson, J.D.; Goettsch, C.; Bertazzo, S.; Maldonado, N.; Ruiz, J.L.; Goh, W.; Yabusaki, K.; Faits, T.; Bouten, C.; Franck, G.; et al. Genesis and Growth of Extracellular-Vesicle-Derived Microcalcification in Atherosclerotic Plaques. Nat. Mater. 2016, 15, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Libby, P. The Changing Landscape of Atherosclerosis. Nature 2021, 592, 524–533. [Google Scholar] [CrossRef]
- Yu, Y.; Delbianco, M. Conformational Studies of Oligosaccharides. Chem. Eur. J. 2020, 26, 9814–9825. [Google Scholar] [CrossRef] [PubMed]
- Hofman, D.L.; Van Buul, V.J.; Brouns, F.J.P.H. Nutrition, Health, and Regulatory Aspects of Digestible Maltodextrins. Crit. Rev. Food Sci. Nutr. 2016, 56, 2091–2100. [Google Scholar] [CrossRef]
- Lordan, C.; Thapa, D.; Ross, R.P.; Cotter, P.D. Potential for Enriching Next-Generation Health-Promoting Gut Bacteria through Prebiotics and Other Dietary Components. Gut Microbes 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyrikos-Ergas, T.; Fittolani, G.; Seeberger, P.H.; Delbianco, M. Structural Studies Using Unnatural Oligosaccharides: Toward Sugar Foldamers. Biomacromolecules 2020, 21, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, N.P.; Le, V.; Sligar, A.D.; Mei, L.; Chavarria, D.; Yang, E.Y.; Baker, A.B. Algal Polysaccharides as Therapeutic Agents for Atherosclerosis. Front. Cardiovasc. Med. 2018, 5, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Wang, H.; Liu, N.; Zhao, K.; Sheng, Y.; Pang, H.; Shao, K.; Zhang, M.; Li, S.; He, N. Algal Polysaccharides and Derivatives as Potential Therapeutics for Obesity and Related Metabolic Diseases. Food Funct. 2022, 13, 11387–11409. [Google Scholar] [CrossRef]
- Vourakis, M.; Mayer, G.; Rousseau, G. The Role of Gut Microbiota on Cholesterol Metabolism in Atherosclerosis. Int. J. Mol. Sci. 2021, 22, 8074. [Google Scholar] [CrossRef]
- Shen, X.; Li, L.; Sun, Z.; Zang, G.; Zhang, L.; Shao, C.; Wang, Z. Gut Microbiota and Atherosclerosis—Focusing on the Plaque Stability. Front. Cardiovasc. Med. 2021, 8, 668532. [Google Scholar] [CrossRef]
- Zhao, X.; Oduro, P.K.; Tong, W.; Wang, Y.; Gao, X.; Wang, Q. Therapeutic Potential of Natural Products against Atherosclerosis: Targeting on Gut Microbiota. Pharmacol. Res. 2021, 163, 105362. [Google Scholar] [CrossRef]
- Xing, M.; Cao, Q.; Wang, Y.; Xiao, H.; Zhao, J.; Zhang, Q.; Ji, A.; Song, S. Advances in Research on the Bioactivity of Alginate Oligosaccharides. Mar. Drugs 2020, 18, 144. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yang, S.; Li, X.; Yan, Q.; Reaney, M.J.T.; Jiang, Z. Alginate Oligosaccharides: Production, Biological Activities, and Potential Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1859–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huwait, E.; Al-Saedi, D.A.; Mirza, Z. Anti-Inflammatory Potential of Fucoidan for Atherosclerosis: In Silico and in vitro Studies in THP-1 Cells. Molecules 2022, 27, 3197. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-J.; Ma, L.-L.; Shi, H.-T.; Zhu, J.-B.; Wu, J.; Ding, Z.-W.; An, Y.; Zou, Y.-Z.; Ge, J.-B. Alginate Oligosaccharide Prevents Acute Doxorubicin Cardiotoxicity by Suppressing Oxidative Stress and Endoplasmic Reticulum-Mediated Apoptosis. Mar. Drugs 2016, 14, 231. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.Y.; Sanada, S.; Matsuzaki, T.; Liao, Y.; Okuda, K.; Yamato, M.; Tsuchida, S.; Araki, R.; Asano, Y.; Asanuma, H.; et al. Chemical Endoplasmic Reticulum Chaperone Alleviates Doxorubicin-Induced Cardiac Dysfunction. Circ. Res. 2016, 118, 798–809. [Google Scholar] [CrossRef]
- Guo, J.-J.; Xu, F.-Q.; Li, Y.-H.; Li, J.; Liu, X.; Wang, X.-F.; Hu, L.-G.; An, Y. Alginate Oligosaccharide Alleviates Myocardial Reperfusion Injury by Inhibiting Nitrative and Oxidative Stress and Endoplasmic Reticulum Stress-Mediated Apoptosis. Drug Des. Dev. Ther. 2017, 11, 2387–2397. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Han, Y.; Wang, Z.; Zhang, R.; Wang, G.; Mao, Y. Alginate Oligosaccharide Protects Endothelial Cells against Oxidative Stress Injury via Integrin-α/FAK/PI3K Signaling. Biotechnol. Lett. 2020, 42, 2749–2758. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, X.; Wu, L.; Li, H.; Chen, Y.; Li, L.; Ni, H.; Li, Q.; Zhu, Y. Exolytic Products of Alginate by the Immobilized Alginate Lyase Confer Antioxidant and Antiapoptotic Bioactivities in Human Umbilical Vein Endothelial Cells. Carbohydr. Polym. 2021, 251, 116976. [Google Scholar] [CrossRef]
- Feng, W.; Liu, J.; Wang, S.; Hu, Y.; Pan, H.; Hu, T.; Guan, H.; Zhang, D.; Mao, Y. Alginate Oligosaccharide Alleviates D-galactose-induced Cardiac Ageing via Regulating Myocardial Mitochondria Function and Integrity in Mice. J. Cell Mol. Med. 2021, 25, 7157–7168. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Luo, T.; Liu, S.; Song, G.; Han, J.; Wang, Y.; Yao, S.; Feng, L.; Qin, S. Chitosan Oligosaccharides Attenuate Atherosclerosis and Decrease Non-HDL in ApoE−/− Mice. J. Atheroscler. Thromb. 2015, 22, 926–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Zhang, J.; Chen, L.; Wu, Q.; Yu, C. Chitosan Oligosaccharides Enhance Lipid Droplets via Down-Regulation of PCSK9 Gene Expression in HepG2 Cells. Exp. Cell Res. 2018, 366, 152–160. [Google Scholar] [CrossRef]
- Stefan, J.; Kus, K.; Wisniewska, A.; Lorkowska-Zawicka, B.; Kaminski, K.; Szczubialka, K.; Nowakowska, M.; Korbut, R. The Antiatherogenic Effect of New Biocompatible Cationically Modified Polysaccharides: Chitosan and Pullulan-the Comparison Study. J. Physiol. Pharmacol. 2018, 69, 26402. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Y.; Ma, X.; Zhang, B.; Huang, Y.; Zhao, J.; Wang, S.; Li, Y.; Zhu, Y.; Xiong, J.; et al. Synthesis and Characterization of Fucoidan-Chitosan Nanoparticles Targeting P-Selectin for Effective Atherosclerosis Therapy. Oxidative Med. Cell. Longev. 2022, 2022, 8006642. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-R.; Wu, J.; Yu, X.-C.; Qian, X.; Xiong, R.; Wang, H.-F.; Yu, D.-F.; Liu, F.-F.; He, S.-Y. Heparin-Derived Oligosaccharide Inhibits Vascular Intimal Hyperplasia in Balloon-Injured Carotid Artery. Chin. J. Nat. Med. 2017, 15, 442–450. [Google Scholar] [CrossRef]
- Liu, J.; Wang, H.; Yu, D.; Chen, X.; He, S. Modulation of Binding to Vascular Endothelial Growth Factor and Receptor by Heparin Derived Oligosaccharide. Carbohydr. Polym. 2017, 174, 558–564. [Google Scholar] [CrossRef]
- Hashmi, A.; Naeem, N.; Farooq, Z.; Masood, S.; Iqbal, S.; Naseer, R. Effect of Prebiotic Galacto-Oligosaccharides on Serum Lipid Profile of Hypercholesterolemics. Probiotics Antimicro. Prot. 2016, 8, 19–30. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Sandhu, Y.K.; Korzun, W.J.; Ghosh, S. Dietary Supplementation with Galactooligosaccharides Attenuates High-Fat, High-Cholesterol Diet-Induced Glucose Intolerance and Disruption of Colonic Mucin Layer in C57BL/6 Mice and Reduces Atherosclerosis in Ldlr−/− Mice. J. Nutr. 2020, 150, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.; Huang, X.; Cao, H.; Bai, Y.; Che, Q.; Nie, H.; Su, Z. Anti-Obesity Effects of Galacto-Oligosaccharides in Obese Rats. Eur. J. Pharmacol. 2022, 917, 174728. [Google Scholar] [CrossRef]
- Boyanov, K.O.; Choneva, M.A.; Dimov, I.; Dimitrov, I.V.; Vlaykova, T.I.; Gerginska, F.D.; Delchev, S.D.; Hrischev, P.I.; Georgieva, K.N.; Bivolarska, A.V. Effect of Oligosaccharides on the Antioxidant, Lipid and Inflammatory Profiles of Rats with Streptozotocin-Induced Diabetes Mellitus. Z. Naturforschung C 2022, 77, 379–386. [Google Scholar] [CrossRef]
- Sakurai, T.; Sakurai, A.; Chen, Y.; Vaisman, B.L.; Amar, M.J.; Pryor, M.; Thacker, S.G.; Zhang, X.; Wang, X.; Zhang, Y.; et al. Dietary α-Cyclodextrin Reduces Atherosclerosis and Modifies Gut Flora in Apolipoprotein E-Deficient Mice. Mol. Nutr. Food Res. 2017, 61, 1600804. [Google Scholar] [CrossRef]
- Coisne, C.; Hallier-Vanuxeem, D.; Boucau, M.-C.; Hachani, J.; Tilloy, S.; Bricout, H.; Monflier, E.; Wils, D.; Serpelloni, M.; Parissaux, X.; et al. β-Cyclodextrins Decrease Cholesterol Release and ABC-Associated Transporter Expression in Smooth Muscle Cells and Aortic Endothelial Cells. Front. Physiol. 2016, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Han, J.; Park, J.-H. Cyclodextrin Polymer Improves Atherosclerosis Therapy and Reduces Ototoxicity. J. Control. Release 2020, 319, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X.; Yu, B.; Peng, X.; Liu, Y.; Wang, A.; Zhao, D.; Pang, D.; OuYang, H.; Tang, X. Cyclodextrin Ameliorates the Progression of Atherosclerosis via Increasing High-Density Lipoprotein Cholesterol Plasma Levels and Anti-Inflammatory Effects in Rabbits. J. Cardiovasc. Pharmacol. 2019, 73, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, S.; Grebe, A.; Bakke, S.S.; Bode, N.; Halvorsen, B.; Ulas, T.; Skjelland, M.; De Nardo, D.; Labzin, L.I.; Kerksiek, A.; et al. Cyclodextrin Promotes Atherosclerosis Regression via Macrophage Reprogramming. Sci. Transl. Med. 2016, 8, 333ra50. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Lund, F.W.; Haka, A.S.; Maxfield, F.R. High-Density Lipoprotein or Cyclodextrin Extraction of Cholesterol from Aggregated LDL Reduces Foam Cell Formation. J. Cell Sci. 2019, 132, jcs.237271. [Google Scholar] [CrossRef]
- Ao, M.; Gan, C.; Shao, W.; Zhou, X.; Chen, Y. Effects of Cyclodextrins on the Structure of LDL and Its Susceptibility to Copper-Induced Oxidation. Eur. J. Pharm. Sci. 2016, 91, 183–189. [Google Scholar] [CrossRef]
- Ao, M.; Chen, Y. Effects of MβCD on Lipoxygenase-Induced LDL Oxidation. Chem. Pharm. Bull. 2017, 65, 200–203. [Google Scholar] [CrossRef] [Green Version]
- Ao, M.; Wu, L.; Zhou, X.; Chen, Y. Methyl-β-Cyclodextrin Impairs the Monocyte-Adhering Ability of Endothelial Cells by Down-Regulating Adhesion Molecules and Caveolae and Reorganizing the Actin Cytoskeleton. Biol. Pharm. Bull. 2016, 39, 1029–1034. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.; Lee, S.; Park, H. β-Cyclodextrin Inhibits Monocytic Adhesion to Endothelial Cells through Nitric Oxide-Mediated Depletion of Cell Adhesion Molecules. Molecules 2020, 25, 3575. [Google Scholar] [CrossRef]
- Chen, G.; Zhou, Y.; Zhang, W.; Qin, Y.; Wei, B.; Sun, Y.; Chen, Y. Methyl-β-Cyclodextrin Suppresses the Monocyte-Endothelial Adhesion Triggered by Lipopolysaccharide (LPS) or Oxidized Low-Density Lipoprotein (OxLDL). Pharm. Biol. 2021, 59, 1034–1042. [Google Scholar] [CrossRef]
- Guo, J.; Li, D.; Tao, H.; Li, G.; Liu, R.; Dou, Y.; Jin, T.; Li, L.; Huang, J.; Hu, H.; et al. Cyclodextrin-Derived Intrinsically Bioactive Nanoparticles for Treatment of Acute and Chronic Inflammatory Diseases. Adv. Mater. 2019, 31, 1904607. [Google Scholar] [CrossRef] [PubMed]
- Bakke, S.S.; Aune, M.H.; Niyonzima, N.; Pilely, K.; Ryan, L.; Skjelland, M.; Garred, P.; Aukrust, P.; Halvorsen, B.; Latz, E.; et al. Cyclodextrin Reduces Cholesterol Crystal–Induced Inflammation by Modulating Complement Activation. J. Immunol. 2017, 199, 2910–2920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilely, K.; Bakke, S.S.; Palarasah, Y.; Skjoedt, M.-O.; Bartels, E.D.; Espevik, T.; Garred, P. Alpha-Cyclodextrin Inhibits Cholesterol Crystal-Induced Complement-Mediated Inflammation: A Potential New Compound for Treatment of Atherosclerosis. Atherosclerosis 2019, 283, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, T.; Yashiro, M.; Nishioka, N.; Hirakawa, K.; Olden, K.; Roberts, J.D. PI3K/Akt Signalling Is Required for the Attachment and Spreading, and Growth in vivo of Metastatic Scirrhous Gastric Carcinoma. Br. J. Cancer 2012, 106, 1535–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-R.; Zhao, J.-T.; Xie, Z.-Z. Integrin-Mediated Cancer Progression as a Specific Target in Clinical Therapy. Biomed. Pharmacother. 2022, 155, 113745. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; You, J.; Fu, J.; Wang, X.; Zhang, Y. Phosphatidylinositol 3-Kinase/Akt Mediates Integrin Signaling to Control RNA Polymerase I Transcriptional Activity. Mol. Cell. Biol. 2016, 36, 1555–1568. [Google Scholar] [CrossRef] [Green Version]
- Van Der Gronde, T.; Hartog, A.; Van Hees, C.; Pellikaan, H.; Pieters, T. Systematic Review of the Mechanisms and Evidence behind the Hypocholesterolaemic Effects of HPMC, Pectin and Chitosan in Animal Trials. Food Chem. 2016, 199, 746–759. [Google Scholar] [CrossRef]
- Tao, W.; Wang, G.; Wei, J. The Role of Chitosan Oligosaccharide in Metabolic Syndrome: A Review of Possible Mechanisms. Mar. Drugs 2021, 19, 501. [Google Scholar] [CrossRef]
- Li, L.; Li, W.; Ren, Z.; He, S.; Xu, G.; Yang, L. Effect of Heparin-Derived Oligosaccharide on Vascular Smooth Muscle Cell Proliferation. Vasc. Endovasc. Surg. 2012, 46, 393–400. [Google Scholar] [CrossRef]
- Yu, D.; Rui, X.; He, S. Effect of Heparin-Derived Oligosaccharide on BFGFR1 and BFGFR2 in Vascular Smooth Muscle Cells. Vasc. Endovasc. Surg. 2014, 48, 289–296. [Google Scholar] [CrossRef]
- Abdel-Razek, O.; Sadananda, S.N.; Li, X.; Cermakova, L.; Frohlich, J.; Brunham, L.R. Increased Prevalence of Clinical and Subclinical Atherosclerosis in Patients with Damaging Mutations in ABCA1 or APOA1. J. Clin. Lipidol. 2018, 12, 116–121. [Google Scholar] [CrossRef]
- Ogura, M. HDL, Cholesterol Efflux, and ABCA1: Free from Good and Evil Dualism. J. Pharmacol. Sci. 2022, 150, 81–89. [Google Scholar] [CrossRef]
- Hellesøy, M.; Lorens, J.B. Cellular Context–Mediated Akt Dynamics Regulates MAP Kinase Signaling Thresholds during Angiogenesis. Mol. Biol. Cell 2015, 26, 2698–2711. [Google Scholar] [CrossRef] [PubMed]
- Mei, Z.; Yuan, J.; Li, D. Biological Activity of Galacto-Oligosaccharides: A Review. Front. Microbiol. 2022, 13, 993052. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, M.; Mao, H. Cross-linked Enzyme Aggregates of Recombinant Cyclodextrin Glycosyltransferase for High-purity Β-cyclodextrin Production. J. Chem. Technol. Biotechnol. 2019, 94, 1528–1533. [Google Scholar] [CrossRef]
- Fang, F.; Huang, X.-J.; Guo, Y.-Z.; Hong, X.; Wu, H.-M.; Liu, R.; Chen, D.-J. Selective and Regenerable Surface Based on β-Cyclodextrin for Low-Density Lipoprotein Adsorption. Langmuir 2018, 34, 8163–8169. [Google Scholar] [CrossRef] [PubMed]
- Fenyvesi, É.; Puskás, I.; Szente, L. Applications of Steroid Drugs Entrapped in Cyclodextrins. Environ. Chem. Lett. 2019, 17, 375–391. [Google Scholar] [CrossRef]
- Saokham, P.; Muankaew, C.; Jansook, P.; Loftsson, T. Solubility of Cyclodextrins and Drug/Cyclodextrin Complexes. Molecules 2018, 23, 1161. [Google Scholar] [CrossRef] [Green Version]
- Komiyama, M. Cyclodextrins as Eminent Constituents in Nanoarchitectonics for Drug Delivery Systems. Beilstein J. Nanotechnol. 2023, 14, 218–232. [Google Scholar] [CrossRef]
- Liu, J.; Ding, X.; Fu, Y.; Xiang, C.; Yuan, Y.; Zhang, Y.; Yu, P. Cyclodextrins Based Delivery Systems for Macro Biomolecules. Eur. J. Med. Chem. 2021, 212, 113105. [Google Scholar] [CrossRef]
- Topuz, F.; Uyar, T. Advances in the Development of Cyclodextrin-Based Nanogels/Microgels for Biomedical Applications: Drug Delivery and Beyond. Carbohydr. Polym. 2022, 297, 120033. [Google Scholar] [CrossRef]
- Zhang, L. Cyclodextrin Related Drug Delivery System to Promote Atherosclerosis Regression. Pharmazie 2020, 75, 619–625. [Google Scholar]
- Snip, O.S.C.; Hoekstra, M.; Zhang, Y.; Geerling, J.J.; Van Eck, M. 2-Hydroxypropyl-Beta-Cyclodextrin Treatment Does Not Induce Atherosclerotic Lesion Regression in Western-Type Diet-Fed Apolipoprotein E Knockout Mice. Biomolecules 2022, 12, 1205. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, Y.; Nabavi, S.M.; Sahebkar, A.; Little, P.J.; Xu, S.; Weng, J.; Ge, J. Mechanisms of Oxidized LDL-Mediated Endothelial Dysfunction and Its Consequences for the Development of Atherosclerosis. Front. Cardiovasc. Med. 2022, 9, 925923. [Google Scholar] [CrossRef]
- Pouw, R.B.; Ricklin, D. Tipping the Balance: Intricate Roles of the Complement System in Disease and Therapy. Semin. Immunopathol. 2021, 43, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wu, Q.; Shuai, Z.-W. Emerging Role of Ficolins in Autoimmune Diseases. Pharmacol. Res. 2021, 163, 105266. [Google Scholar] [CrossRef]
- Dobó, J.; Kocsis, A.; Dani, R.; Gál, P. Proprotein Convertases and the Complement System. Front. Immunol. 2022, 13, 958121. [Google Scholar] [CrossRef] [PubMed]
- Barratt-Due, A.; Pischke, S.E.; Nilsson, P.H.; Espevik, T.; Mollnes, T.E. Dual Inhibition of Complement and Toll-like Receptors as a Novel Approach to Treat Inflammatory Diseases—C3 or C5 Emerge Together with CD14 as Promising Targets. J. Leukoc. Biol. 2017, 101, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Samstad, E.O.; Niyonzima, N.; Nymo, S.; Aune, M.H.; Ryan, L.; Bakke, S.S.; Lappegård, K.T.; Brekke, O.-L.; Lambris, J.D.; Damås, J.K.; et al. Cholesterol Crystals Induce Complement-Dependent Inflammasome Activation and Cytokine Release. J. Immunol. 2014, 192, 2837–2845. [Google Scholar] [CrossRef] [Green Version]
- Niyonzima, N.; Bakke, S.S.; Gregersen, I.; Holm, S.; Sandanger, Ø.; Orrem, H.L.; Sporsheim, B.; Ryan, L.; Kong, X.Y.; Dahl, T.B.; et al. Cholesterol Crystals Use Complement to Increase NLRP3 Signaling Pathways in Coronary and Carotid Atherosclerosis. EBioMedicine 2020, 60, 102985. [Google Scholar] [CrossRef] [PubMed]
- Pilely, K.; Rosbjerg, A.; Genster, N.; Gal, P.; Pál, G.; Halvorsen, B.; Holm, S.; Aukrust, P.; Bakke, S.S.; Sporsheim, B.; et al. Cholesterol Crystals Activate the Lectin Complement Pathway via Ficolin-2 and Mannose-Binding Lectin: Implications for the Progression of Atherosclerosis. J. Immunol. 2016, 196, 5064–5074. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; Yazdi, A.S.; Biedermann, T. Clinical Spectrum of α-Gal Syndrome: From Immediate-Type to Delayed Immediate-Type Reactions to Mammalian Innards and Meat. Allergo J. Int. 2016, 25, 55–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, J.M.; Nguyen, A.T.; Schuyler, A.J.; Commins, S.P.; Taylor, A.M.; Platts-Mills, T.A.E.; McNamara, C.A. IgE to the Mammalian Oligosaccharide Galactose-α-1,3-Galactose Is Associated with Increased Atheroma Volume and Plaques with Unstable Characteristics—Brief Report. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1665–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vernon, S.T.; Kott, K.A.; Hansen, T.; Finemore, M.; Baumgart, K.W.; Bhindi, R.; Yang, J.; Hansen, P.S.; Nicholls, S.J.; Celermajer, D.S.; et al. Immunoglobulin E Sensitization to Mammalian Oligosaccharide Galactose-α-1,3 (α-Gal) Is Associated with Noncalcified Plaque, Obstructive Coronary Artery Disease, and ST-Segment–Elevated Myocardial Infarction. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 352–361. [Google Scholar] [CrossRef]
- Perazza, L.R.; Mitchell, P.L.; Jensen, B.A.H.; Daniel, N.; Boyer, M.; Varin, T.V.; Bouchareb, R.; Nachbar, R.T.; Bouchard, M.; Blais, M.; et al. Dietary Sucrose Induces Metabolic Inflammation and Atherosclerotic Cardiovascular Diseases More than Dietary Fat in LDLr ApoB100/100 Mice. Atherosclerosis 2020, 304, 9–21. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Oligosaccharide | General Effect of Oligosaccharide Treatment | Proposed Molecular Mechanism | Reference |
|---|---|---|---|
| Alginate oligosaccharide (AOS)—2 to 25 monomers of β-1,4-mannuronic acid and α-1,4-guluronic acid boned by 1, 4-glycosidic linkages | increased mouse survival rate and improved DOX-induced cardiac dysfunction, and attenuated myocardial apoptosis | decreased the expression of pro-apoptotic proteins (CHOP and Bax) and increased the expression of anti-apoptotic protein Bcl-2 | [24] |
| decreased myocardial apoptosis and attenuated oxidative stress; decreased the infarct size and ameliorated cardiac dysfunction after I/R injury in mice | decreased generation of superoxide, 4-hydroxynonenal, and 3-nitrotyrosine; down-regulated NADPH oxidase 2 and inducible nitric oxide synthase; up-regulated Blc-2, and down-regulated CHOP, caspase-12, Bax, and glucose-regulated protein 78 | [26] | |
| protected HUVECs against oxidative stress-induced apoptosis | regulated the expression of P21, FAK, CDK2, integrin-α, PTEN, and PI3K; increased the expression of Bcl-2, and decreased levels of caspase 3 and Bax | [27] | |
| protected HUVECs against H2O2-induced oxidative stress and apoptosis | up-regulated antioxidant enzymes (SOD and CAT) and glutathione, and reduced the activities of caspase-3 and caspase-9 | [28] | |
| preserved ejection fraction and fractional shortening; improved mitochondrial biogenesis, turn-over and integrity in mouse model of D-galactose-induced cardiac ageing | inhibited the D-galactose-induced up-regulation of ageing markers p21 and p53, and cardiac peptide hormones BNP and ANP; increased the mtDNA copy number, autophagy rate, sustained mitochondrial membrane potential and up-regulated the expression of PGC-1α and SIRT3 | [29] | |
| Chitosan oligosaccharide (COS), a linear oligomer composed of β-(1 ➔ 4)-linked N-acetyl-d-glucosamine and d-glucosamine | decreased the lesion and plaque areas in aortic root, increased plaque stability, and reduced plasma triglycerides, cholesterol, apoB100, and apoB48 levels in ApoE−/− mice fed on HFD | increased the expression of SR-BI and ABCA1 in macrophages in vitro, and SR-BI and LDL-R in liver | [30] |
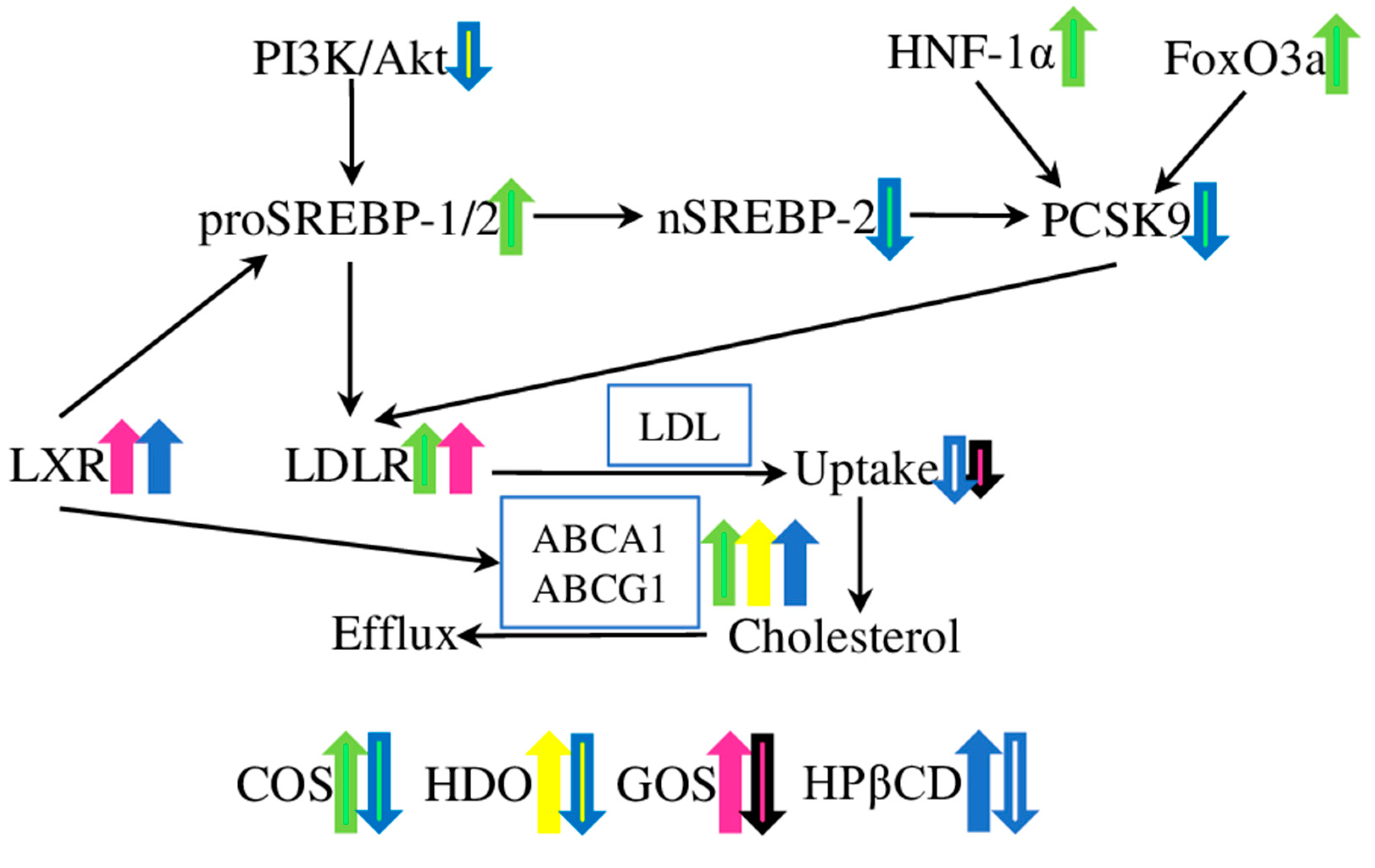
| improved lipid uptake by liver and decreased lipid concentration in the serum | down-regulated the mRNA levels of PCSK9; increased the expression of SREBP-2 and HNF-1α in total cell lysates; decreased the levels of active nSREBP-2, and increased the levels of FOXO3a in nuclear lysates | [31] | |
| demonstrated lipid-lowering properties, inhibited atherosclerotic plaque development | decreased the plasma LDL cholesterol level by 32%; reduced the area of atherosclerotic plaque by 33% in ApoE–/– mice; reduced the expression of HMGCR in HepG2 cells | [32] | |
| chitosan–fucoidan nanoparticles (CFNs) | exhibited antioxidant and anti-inflammatory activities; intravenously injected CFNs decreased the average plaque area (by 36.5%), and necrotic core area, and enhanced the fibrous cap thickness around the plaques, thus stabilising atherosclerotic plaques in atherosclerotic ApoE–/– mice | - | [33] |
| Heparin-derived oligosaccharides (HDOs); these are structurally diverse, ranging in size from disaccharide to tetradecasaccharide | ameliorated intimal hyperplasia and inhibited the histopathology and restenosis induced by balloon injury in rabbits fed a high-fat diet | decreased expression of SR-BI, MCP-1, VCAM-1, VEGF, and bFGF in the arterial wall; decreased the serum levels of total cholesterol, HDL, LDL, and triglycerides; increased the expression level of ABCA1 | [34] |
| acted like VEGF antagonist in culture of human aortic smooth muscle cells | inhibited the expression of VEGF receptors 1 and 2, and interrupted normal binding between VEGF and VEGF receptors; inhibited the expression of MAPK, PI3K/Akt, and PKC | [35] | |
| Galacto-oligosaccharides (GOS); these consist of β-linked galactose moieties with galactose or glucose at the reducing end, different degrees of polymerisation and configurations | demonstrated lipid-lowering properties in hypercholesterolaemic rats fed on high-fat diet, | decreased levels of total cholesterol, LDL and VLDL cholesterol, and triglycerides | [36] |
| reduced atherosclerotic lesion area; ameliorated metabolic alterations induced by a high-fat Western-type diet in Ldlr−/− mice | decreased levels of circulating macrophages and neutrophils (by 30 and 60%, respectively); improved glucose tolerance and reduced plasma LPS concentrations | [37] | |
| facilitated white-fat browning and thermogenesis, and promoted cholesterol catabolism, thus inhibiting obesity in rats | improved levels of HDL cholesterol and reduced levels of total cholesterol, triglycerides, and LDL cholesterol; increased the expression of fat-browning genes (PRDM16, PPARγ, PGC1α, and UCP1); increased levels of LDL-R, PPARα, cholesterol 7α-hydroxylase proteins and LXRα proteins | [38] | |
| demonstrated antioxidant, lipid-lowering, and anti-inflammatory properties in a rat model of streptozotocin-induced diabetes | reduced levels of MDA, total cholesterol, and CRP | [39] | |
| α- and β- Cyclodextrins (α-CD and β-CD), comprising glucose molecules in the pyranose (six-membered) ring configuration; 6, 8, or 10 glucopyranosides bind with each other to form α-, β-, and γ-cyclodextrin, respectively. | α-CD decreased the aortic atherosclerotic lesions by 65% in ApoE−/− mice fed on high-fat diet | β-CD decreased the intestinal lipid absorption and plasma level of free fatty acids | [40] |
| β-CD | regulated the cellular cholesterol pools in vitro in aortic bovine ECs and SMCs | decreased cholesterol release and the expression levels of ABCA1 and ABCG1 transporters | [41] |
| cyclodextrin polymer (CDP) | inhibited plaque growth in a mouse model of atherosclerosis; demonstrated better pharmacokinetics and reduced ototoxicity at high doses | - | [42] |
| 2-hydroxypropyl-β-cyclodextrin (HPβCD) | reduced the atherosclerosis lesion area and collagen and macrophage content in the lesions in rabbits fed a high-fat diet | reduced inflammatory cytokine and adhesion proteins (TNF-a, IL-6, IL-8, MCP-1, VCAM-1, and ICAM-1) and plasma triglyceride levels; increased plasma HDL cholesterol levels; increased the expression of ABCA1 and ABCG1 transporters in livers and aortic plaques | [43] |
| reduced atherosclerosis plaque size and cholesterol crystal load, and facilitated plaque regression in ApoE−/− mice on a continued cholesterol-rich diet | activated ABCA1 and ABCG1, modulated the expression of the inflammasome sensor NLRP3 and pro-inflammatory cytokines (IL-1β and IL-18); increased cholesterol excretion into the urine in human patients | [44] | |
| extracted free cholesterol from aggregated LDL in macrophages | reduced cholesterol uptake and cholesteryl ester accumulation from aggregated LDL | [45] | |
| α-CD, HPβCD, methyl-βCD (MβCD), and γ-CD | caused a significant separation between lipid and protein components of native LDL in vitro | reduced LDL susceptibility to copper-induced oxidation in a concentration-dependent manner | [46] |
| methyl-βCD and HPβCD | demonstrated anti-oxidant properties in vitro | inhibited lipoxygenase-induced LDL oxidation in a concentration-dependent way, increased the lipoxygenase-induced oxidation of the lipids depleted from LDL | [47] |
| MβCD | impaired the adhesion of THP-1 monocytes to HUVECs | decreased the expression of ICAM-1 and modulated membrane-cytoskeleton coupling; reduced the average length of individual actin filaments and reduced the expression of caveolin-1 and PIP2 levels | [48] |
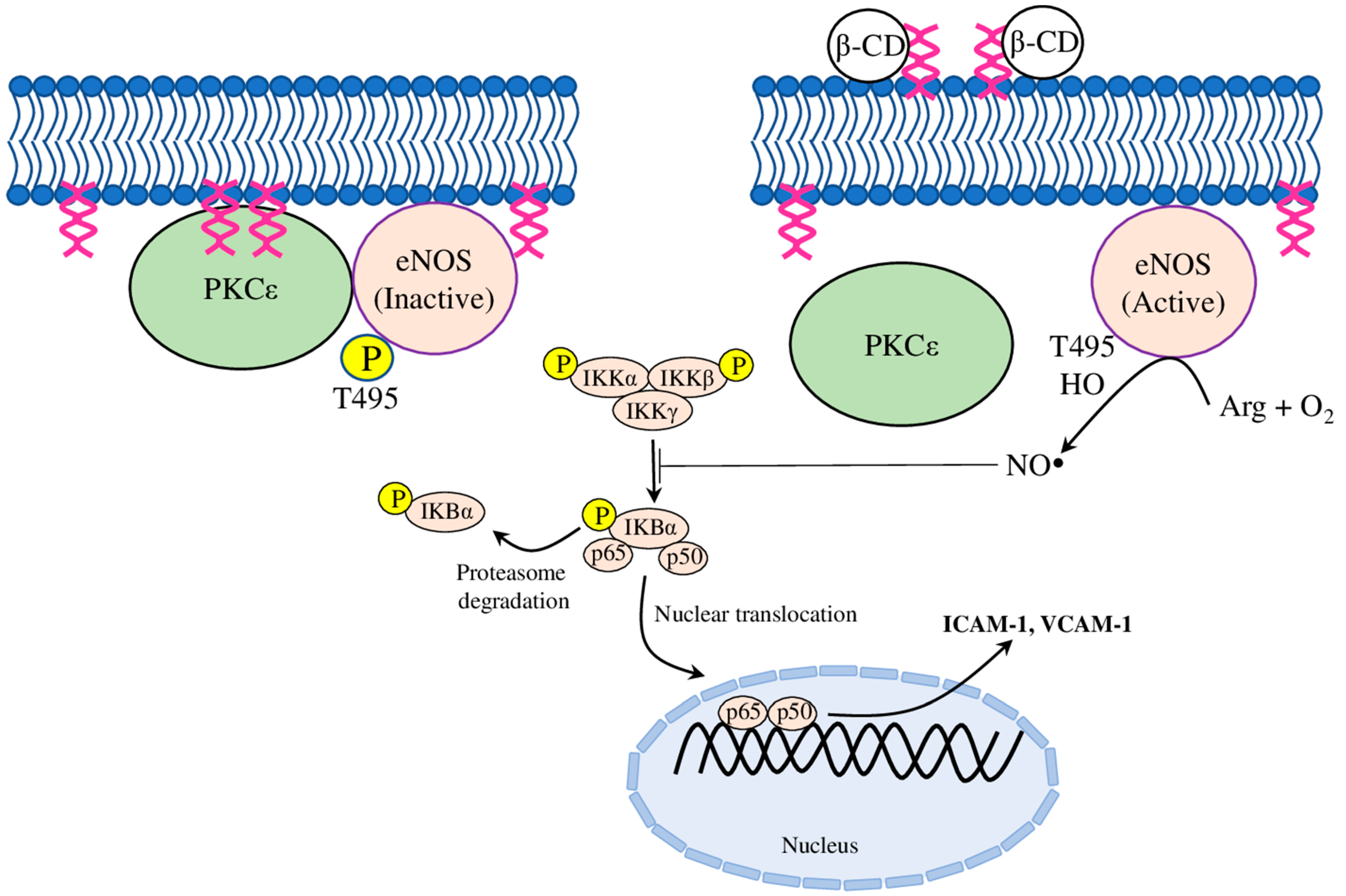
| βCD | regulated NO production in bovine aortic ECs; inhibited monocyte adhesion to ECs | inactivated PKCε, inhibited IκB degradation and subsequent TNFα-induced NFκB activation, thus blocking ICAM-1 and VCAM-1 expression | [49] |
| MβCD | suppressed LPS/oxLDL-induced monocyte-HUVEC adhesion | reduced by 2–3 times the expression of ICAM-1 and VCAM-1 | [50] |
| luminol-conjugated β-cyclodextrin nanoparticles (LCD NP) | inhibited neutrophil and macrophage infiltration and subsequent pro-inflammatory events in both in vitro and in vivo experiments; long-term administration resulted in NP accumulation in atherosclerotic plaques, reduced plaque formation, reduced neutrophils and macrophages number, and increased collagen content around plaques in ApoE−/− mice fed on a high-fat diet | reduced phorbol 12-myristate 13-acetate-induced production of TNFα, IL-1β, and ROS in neutrophils inhibited the MCP-1-stimulated migration of RAW264.7 macrophages | [51] |
| HPβCD | inhibited C3c binding to cholesterol crystals, reduced TCC generation and deposition on the surface of cholesterol crystals through reduced C1q and ficolin-2, and Igs (IgM and IgA) deposition on the surface of cholesterol crystals; increased the phagocytosis of cholesterol crystals in granulocytes | reduced the expression of CR1 and CR3 receptors in monocytes decreased cholesterol crystal-stimulated ROS production in both monocytes and granulocytes; decreased expression of key cytokines (such as TNFα, IL-1α, IL-1β, IL-6 and IL-8), NLRP3 related genes and MIP-1α | [52] |
| αCD | inhibited cholesterol crystal-induced complement activation; reduced phagocytosis in monocytes and granulocytes | prevented binding of C1q and ficolin-2, reduced ROS production in monocytes and granulocytes | [53] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dabravolski, S.A.; Sukhorukov, V.N.; Melnichenko, A.A.; Khotina, V.A.; Orekhov, A.N. Oligosaccharides as Potential Therapeutics against Atherosclerosis. Molecules 2023, 28, 5452. https://doi.org/10.3390/molecules28145452
Dabravolski SA, Sukhorukov VN, Melnichenko AA, Khotina VA, Orekhov AN. Oligosaccharides as Potential Therapeutics against Atherosclerosis. Molecules. 2023; 28(14):5452. https://doi.org/10.3390/molecules28145452
Chicago/Turabian StyleDabravolski, Siarhei A., Vasily N. Sukhorukov, Alexandra A. Melnichenko, Victoria A. Khotina, and Alexander N. Orekhov. 2023. "Oligosaccharides as Potential Therapeutics against Atherosclerosis" Molecules 28, no. 14: 5452. https://doi.org/10.3390/molecules28145452
APA StyleDabravolski, S. A., Sukhorukov, V. N., Melnichenko, A. A., Khotina, V. A., & Orekhov, A. N. (2023). Oligosaccharides as Potential Therapeutics against Atherosclerosis. Molecules, 28(14), 5452. https://doi.org/10.3390/molecules28145452