
Edible Insects an Alternative Nutritional Source of Bioactive Compounds: A Review

 ,
,  ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Identification and Characterization of Phenolic Compounds in Insects
3. Insects’ Bioactive Compounds as a Function of Diet
4. Biological Activity of Polyphenols Extracts Obtained from Insects
4.1. Antioxidant Bioactivity
4.2. Other Biological Effects
4.3. Phenols’ Internal Synthesis and Immune Defences
5. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Tang, C.; Yang, D.; Liao, H.; Sun, H.; Liu, C.; Wei, L.; Li, F. Edible insects as a food source: A review. Food Prod. Process. Nutr. 2019, 1, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wendin, K.; Olsson, V.; Langton, M. Mealworms as food ingredient sensory investigation of a model system. Foods 2019, 8, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongema, Y. List of Edible Insects of the World (1 April 2017); Wageningen UR: Wageningen, The Netherlands, 2017. Available online: https://www.wur.nl/en/Research-Results/Chair-groups/Plant-Sciences/Laboratory-of-Entomology/Edible-insects/Worldwide-species-list.htm (accessed on 3 November 2021).
- Żuk-Gołaszewska, K.; Gałęcki, R.; Obremski, K.; Smetana, S.; Figiel, S.; Gołaszewski, J. Edible Insect Farming in the Context of the EU Regulations and Marketing An Overview. Insects 2022, 13, 446. [Google Scholar] [CrossRef] [PubMed]
- El Hajj, R.; Mhemdi, H.; Besombes, C.; Allaf, K.; Lefrançois, V.; Vorobiev, E. Edible Insects’ Transformation for Feed and Food Uses An Overview of Current Insights and Future Developments in the Field. Processes 2022, 10, 970. [Google Scholar] [CrossRef]
- Sun-Waterhouse, D.; Waterhouse, G.I.; You, L.; Zhang, J.; Liu, Y.; Ma, L.; Gao, J.; Dong, Y. Transforming insect biomass into consumer wellness foods: A review. Food Res. Int. 2016, 89, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Lucas, A.J.; de Oliveira, L.M.; Da Rocha, M.; Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chem. 2020, 311, 126022. [Google Scholar] [CrossRef]
- Ordoñez-Araque, R.; Egas-Montenegro, E. Edible insects: A food alternative for the sustainable development of the planet. Int. J. Gastron. Food Sci. 2021, 23, 100304. [Google Scholar] [CrossRef]
- Hanboonsong, Y.; Jamjanya, T.; Durst, P.B. Six-legged livestock: Edible insect farming, collection and marketing in Thailand. RAP Publ. 2013, 3, 8–21. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Looking at Edible Insects from a Food Safety Perspective; Challenges and Opportunities for the Sector; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Dobermann, D.; Swift, J.A.; Field, L.M. Opportunities and hurdles of edible insects for food and feed. Nutr. Bull. 2017, 42, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, T.K.; Jeong, C.H.; Yong, H.I.; Cha, J.Y.; Kim, B.K.; Choi, Y.S. Biological activity and processing technologies of edible insects: A review. Food Sci. Biotechnol. 2021, 30, 1003–1023. [Google Scholar] [CrossRef]
- Escobar-Ortiz, A.; Hernández-Saavedra, D.; Lizardi-Mendoza, J.; Pérez-Ramírez, I.F.; Mora-Izaguirre, O.; Ramos-Gómez, M.; Reynoso-Camacho, R. Consumption of cricket Acheta domesticus flour decreases insulin resistance and fat accumulation in rats fed with high-fat and-fructose diet. J. Food Biochem. 2022, 46, e14269. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sun, J.; Yu, L.; Zhang, C.; Bi, J.; Zhu, F.; Qu, M.; Yang, Q. Antioxidant activity and phenolic compounds of Holotrichia parallela Motschulsky extracts. Food Chem. 2012, 134, 1885–1891. [Google Scholar] [CrossRef] [PubMed]
- Ssepuuya, G.; Kagulire, J.; Katongole, J.; Kabbo, D.; Claes, J.; Nakimbugwe, D. Suitable extraction conditions for determination of total anti-oxidant capacity and phenolic compounds in Ruspolia differens Serville. J. Insects Food Feed 2021, 7, 205–214. [Google Scholar] [CrossRef]
- Nino, M.C.; Reddivari, L.; Ferruzzi, M.G.; Liceaga, A.M. Targeted phenolic characterization and antioxidant bioactivity of extracts from edible Acheta domesticus. Foods 2021, 10, 2295. [Google Scholar] [CrossRef]
- Nino, M.C.; Reddivari, L.; Osorio, C.; Kaplan, I.; Liceaga, A.M. Insects as a source of phenolic compounds and potential health benefits. J. Insects Food Feed 2021, 7, 1077–1087. [Google Scholar] [CrossRef]
- Zielinska, E.; Baraniak, B.; Karaś, M. Antioxidant and anti-inflammatory activities of hydrolysates and peptide fractions obtained by enzymatic hydrolysis of selected heat-treated edible insects. Nutrients 2017, 9, 970. [Google Scholar] [CrossRef] [Green Version]
- David-Birman, T.; Raften, G.; Lesmes, U. Effects of thermal treatments on the colloidal properties, antioxidant capacity and in-vitro proteolytic degradation of cricket flour. Food Hydrocoll. 2018, 79, 48–54. [Google Scholar] [CrossRef]
- Issaoui, M.; Delgado, A.M.; Caruso, G.; Micali, M.; Barbera, M.; Atrous, H.; Ouslati, A.; Chammem, N. Phenols, flavors, and the Mediterranean diet. J. AOAC Int. 2020, 103, 915–924. [Google Scholar] [CrossRef]
- Nardini, M. Phenolic compounds in food: Characterization and health benefits. Molecules 2022, 27, 783. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, S.; Wei, X.; Xiao, J.; Huang, D. Characterization, Antioxidant Activities, and Pancreatic Lipase Inhibitory Effect of Extract From the Edible Insect Polyrhachis vicina. Front. Nutr. 2022, 9, 860174. [Google Scholar] [CrossRef]
- Sadat, A.; Biswas, T.; Cardoso, M.H.; Mondal, R.; Ghosh, A.; Dam, P.; Nesa, J.; Chakraborty, J.; Bhattacharjya, D.; Franco, O.L.; et al. Silkworm pupae as a future food with nutritional and medicinal benefits. Curr. Opin. Food Sci. 2022, 44, 100818. [Google Scholar] [CrossRef]
- Del Hierro, J.N.; Gutiérrez-Docio, A.; Otero, P.; Reglero, G.; Martin, D. Characterization, antioxidant activity, and inhibitory effect on pancreatic lipase of extracts from the edible insects Acheta domesticus and Tenebrio molitor. Food Chem. 2020, 309, 125742. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Song, J.L.; Yi, R.; Li, G.; Sun, P.; Park, K.Y.; Suo, H. Comparison of antioxidative effects of Insect tea and its raw tea Kuding tea polyphenols in Kunming mice. Molecules 2018, 23, 204. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.Q.; Wu, X.M.; Wang, Q.; Meng, D.; Huang, Y.M.; Xu, J.L.; Li, Y.; Liu, H.; Xiao, H.; Ding, Z.T. Hydroxytyrosol Dimers from Medicinal Insect Blaps Rynchopetera and the in Vitro Cytotoxic Activity. Nat. Prod. Commun. 2022, 17, 1–8. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.M.; Machì, M.; Cianciosi, D.; Navarro-Hortal, M.D.; Battino, M. Edible insects: A novel nutritious, functional, and safe food alternative. Food Front. 2022, 3, 358–365. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; FAO Forestry Paper No. 171; FAO: Rome, Italy, 2013. [Google Scholar]
- Pimentel, D.; Berger, B.; Filiberto, D.; Newton, M.; Wolfe, B.; Karabinakis, E.; Clark, S.; Poon, E.; Abbet, E.; Nandagopal, S. Water resources: Agricultural and environmental issues. BioScience 2004, 54, 909–918. [Google Scholar] [CrossRef] [Green Version]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Oonincx, D.G.; De Boer, I.J. Environmental impact of the production of mealworms as a protein source for humans—A life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef] [Green Version]
- Oonincx, D.G.; Van Itterbeeck, J.; Heetkamp, M.J.; Van Den Brand, H.; Van Loon, J.J.; Van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [Green Version]
- Mason, J.B.; Black, R.; Booth, S.L.; Brentano, A.B.; Broadbent, B.C.; Connolly, P.; Finley, J.; Goldin, J.; Griffin, T.; Hagen, K.; et al. Fostering strategies to expand the consumption of edible insects: The value of a tripartite coalition between academia, industry, and government. Curr. Dev. Nutr. 2018, 2, nzy056. [Google Scholar] [CrossRef] [Green Version]
- Halloran, A.; Hanboonsong, Y.; Roos, N.; Bruun, S. Life cycle assessment of cricket farming in north-eastern Thailand. J. Clean. Prod. 2017, 156, 83–94. [Google Scholar] [CrossRef]
- Halloran, A.; Roos, N.; Hanboonsong, Y. Cricket farming as a livelihood strategy in Thailand. Geogr. J. 2017, 183, 112–124. [Google Scholar] [CrossRef]
- Kozanayi, W.; Frost, P. Marketing of Mopane Worm in Southern Zimbabwe; Institute of Environmental Studies: Harare, Zimbabwe, 2002; pp. 1–31. [Google Scholar]
- Imathiu, S. Benefits and food safety concerns associated with consumption of edible insects. NFS J. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Derrien, C.; Boccuni, A. Current status of the insect producing industry in Europe. In Edible Insects in Sustainable Food Systems; Halloran, A., Flore, R., Vantomme, P., Roos, N., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 471–479. [Google Scholar]
- Gezondheidsraad, H. Common Advice SciCom 14-2014 and SHC Nr. 9160—Subject: Food Safety Aspects of Insects Intended for Human Consumption; FASFC: Brussels, Belgium, 2014. [Google Scholar]
- Poma, G.; Cuykx, M.; Amato, E.; Calaprice, C.; Focant, J.F.; Covaci, A. Evaluation of hazardous chemicals in edible insects and insect-based food intended for human consumption. Food Chem. Toxicol. 2017, 100, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Kunatsa, Y.; Chidewe, C.; Zvidzai, C.J. Phytochemical and anti-nutrient composite from selected marginalized Zimbabwean edible insects and vegetables. J. Agric. Food Res. 2020, 2, 100027. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Reyes, A.; Rosell, C.M.; Rodrigo, D.; Ibarra-Herrera, C.C. Benefits and challenges in the incorporation of insects in food products. Front. Nutr. 2021, 8, 687712. [Google Scholar] [CrossRef]
- Stoops, J.; Crauwels, S.; Waud, M.; Claes, J.; Lievens, B.; Van Campenhout, L. Microbial community assessment of mealworm larvae Tenebrio molitor and grasshoppers Locusta migratoria migratorioides sold for human consumption. Food Microbiol. 2016, 53, 122–127. [Google Scholar] [CrossRef]
- Leoni, C.; Volpicella, M.; Dileo, M.C.; Gattulli, B.A.; Ceci, L.R. Chitinases as food allergens. Molecules 2019, 24, 2087. [Google Scholar] [CrossRef] [Green Version]
- De Gier, S.; Verhoeckx, K. Insect food allergy and allergens. Mol. Immunol. 2018, 100, 82–106. [Google Scholar] [CrossRef]
- Jeong, K.Y.; Park, J.W. Insect allergens on the dining table. Curr. Protein Pept. Sci. 2020, 21, 159–169. [Google Scholar] [CrossRef]
- Caparros Megido, R.; Desmedt, S.; Blecker, C.; Béra, F.; Haubruge, É.; Alabi, T.; Francis, F. Microbiological load of edible insects found in Belgium. Insects 2017, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, A.; Vecchio, R.; Borrello, M.; Caracciolo, F.; Cembalo, L. Willingness to pay for insect-based food: The role of information and carrier. Food Qual. Prefer. 2019, 72, 177–187. [Google Scholar] [CrossRef]
- Grabowski, N.T.; Tchibozo, S.; Abdulmawjood, A.; Acheuk, F.; M’Saad Guerfali, M.; Sayed, W.A.; Plötz, M. Edible insects in Africa in terms of food, wildlife resource, and pest management legislation. Foods 2020, 9, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, L.H.; de Benoist, B.; Dary, O.; Hurrell, R. Guidelines on Food Fortification with Micronutrients; World Health Organization, Department of Nutrition for Health and Development: Geneva, Switzerland, 2006. [Google Scholar]
- Alfonzo, A.; Martorana, A.; Guarrasi, V.; Barbera, M.; Gaglio, R.; Santulli, A.; Settanni, L.; Galati, A.; Moschetti, G.; Francesca, N. Effect of the lemon essential oils on the safety and sensory quality of salted sardines (Sardina pilchardus Walbaum 1792). Food Control 2017, 73, 1265–1274. [Google Scholar] [CrossRef] [Green Version]
- Alfonzo, A.; Gaglio, R.; Barbera, M.; Francesca, N.; Moschetti, G.; Settanni, L. Evaluation of the fermentation dynamics of commercial baker’s yeast in presence of pistachio powder to produce lysine-enriched breads. Ferment 2019, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Barbera, M. Reuse of food waste and wastewater as a source of polyphenolic compounds to use as food additives. J. AOAC Int. 2020, 103, 906–914. [Google Scholar] [CrossRef]
- Gaglio, R.; Alfonzo, A.; Barbera, M.; Franciosi, E.; Francesca, N.; Moschetti, G.; Settanni, L. Persistence of a mixed lactic acid bacterial starter culture during lysine fortification of sourdough breads by addition of pistachio powder. Food Microbiol. 2020, 86, 103349. [Google Scholar] [CrossRef]
- Gaglio, R.; Restivo, I.; Barbera, M.; Barbaccia, P.; Ponte, M.; Tesoriere, L.; Bonanno, A.; Attanzio, A.; Di Grigoli, A.; Francesca, N.; et al. Effect on the antioxidant, lipoperoxyl radical scavenger capacity, nutritional, sensory and microbiological traits of an ovine stretched cheese produced with grape pomace powder addition. Antioxidants 2021, 10, 306. [Google Scholar] [CrossRef]
- Gaglio, R.; Barbaccia, P.; Barbera, M.; Restivo, I.; Attanzio, A.; Maniaci, G.; Di Grigoli, A.; Francesca, N.; Tesoriere, L.; Bonanno, A.; et al. The use of winery by-products to enhance the functional aspects of the fresh ovine “primosale” cheese. Foods 2021, 10, 461. [Google Scholar] [CrossRef]
- Barbaccia, P.; Busetta, G.; Barbera, M.; Alfonzo, A.; Garofalo, G.; Francesca, N.; Moschetti, G.; Settanni, L.; Gaglio, R. Effect of grape pomace from red cultivar ‘Nero d’Avola’ on the microbiological, physicochemical, phenolic profile and sensory aspects of ovine Vastedda-like stretched cheese. J. Appl. Microbiol. 2022, 133, 130–144. [Google Scholar] [CrossRef]
- Busetta, G.; Ponte, M.; Barbera, M.; Alfonzo, A.; Ioppolo, A.; Maniaci, G.; Guarcella, R.; Francesca, N.; Palazzolo, E.; Bonanno, A.; et al. Influence of Citrus Essential Oils on the Microbiological, Physicochemical and Antioxidant Properties of Primosale Cheese. Antioxidants 2022, 11, 2004. [Google Scholar] [CrossRef] [PubMed]
- Adeboye, A.O.; Bolaji, T.A.; Fatola, O.L. Nutritional composition and sensory evaluation of cookies made from wheat and palm weevil larvae flour blends. Annu. Food Sci. Technol. 2016, 17, 543–547. [Google Scholar]
- Kim, H.W.; Setyabrata, D.; Lee, Y.J.; Jones, O.G.; Kim, Y.H.B. Pre-treated mealworm larvae and silkworm pupae as a novel protein ingredient in emulsion sausages. Innov. Food Sci. Emerg. Technol. 2016, 38, 116–123. [Google Scholar] [CrossRef]
- De Oliveira, L.M.; da Silva Lucas, A.J.; Cadaval, C.L.; Mellado, M.S. Bread enriched with flour from cinereous cockroach Nauphoeta cinerea. Innov. Food Sci. Emerg. Technol. 2017, 44, 30–35. [Google Scholar] [CrossRef]
- González, C.M.; Garzón, R.; Rosell, C.M. Insects as ingredients for bakery goods. A comparison study of H. illucens, A. domestica and T. molitor flours. Innov. Food Sci. Emerg. Technol. 2019, 51, 205–210. [Google Scholar] [CrossRef]
- Vasilica, B.T.B.; Chiș, M.S.; Alexa, E.; Pop, C.; Păucean, A.; Man, S.; Igual, M.; Haydee, K.M.; Dalma, K.E.; Stanila, S. The Impact of Insect Flour on Sourdough Fermentation-Fatty Acids, Amino-Acids, Minerals and Volatile Profile. Insects 2022, 13, 576. [Google Scholar] [CrossRef] [PubMed]
- Kowalczewski, P.Ł.; Gumienna, M.; Rybicka, I.; Górna, B.; Sarbak, P.; Dziedzic, K.; Kmiecik, D. Nutritional value and biological activity of gluten-free bread enriched with cricket powder. Molecules 2021, 26, 1184. [Google Scholar] [CrossRef]
- Zielinska, E.; Pankiewicz, U.; Sujka, M. Nutritional, physiochemical, and biological value of muffins enriched with edible insects flour. Antioxidants 2021, 10, 1122. [Google Scholar] [CrossRef]
- Mendoza-Salazar, A.; Santiago-López, L.; Torres-Llanez, M.J.; Hernández-Mendoza, A.; Vallejo-Cordoba, B.; Liceaga, A.M.; González-Córdova, A.F. In Vitro Antioxidant and Antihypertensive Activity of Edible Insects Flours Mealworm and Grasshopper Fermented with Lactococcus lactis Strains. Ferment 2021, 7, 153. [Google Scholar] [CrossRef]
- Gaglio, R.; Barbera, M.; Tesoriere, L.; Osimani, A.; Busetta, G.; Matraxia, M.; Attanzio, A.; Restivo, I.; Aquilanti, L.; Settanni, L. Sourdough “ciabatta” bread enriched with powdered insects: Physicochemical, microbiological, and simulated intestinal digesta functional properties. Innov. Food Sci. Emerg. Technol. 2021, 72, 102755. [Google Scholar] [CrossRef]
- Szawara-Nowak, D.; Bączek, N.; Zieliński, H. Antioxidant capacity and bioaccessibility of buckwheat-enhanced wheat bread phenolics. J. Food Sci. Technol. 2016, 53, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Hayta, M.; Özuğur, G.; Etgü, H.; Şeker, İ.T. Effect of Grape Vitis vinifera L. Pomace on the Quality, Total Phenolic Content and Anti-Radical Activity of Bread. J. Food Process. Preserv. 2014, 38, 980–986. [Google Scholar] [CrossRef]
- Lachowicz, S.; Świeca, M.; Pejcz, E. Biological activity, phytochemical parameters, and potential bioaccessibility of wheat bread enriched with powder and microcapsules made from Saskatoon berry. Food Chem. 2021, 338, 128026. [Google Scholar] [CrossRef]
- Parkar, S.G.; Trower, T.M.; Stevenson, D.E. Fecal microbial metabolism of polyphenols and its effects on human gut microbiota. Anaerobe 2013, 23, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Shah, N.P. Lactic acid bacterial fermentation modified phenolic composition in tea extracts and enhanced their antioxidant activity and cellular uptake of phenolic compounds following in vitro digestion. J. Funct. Foods 2016, 20, 182–194. [Google Scholar] [CrossRef]
- Ikawa, M.; Schaper, T.D.; Dollard, C.A.; Sasner, J.J. Utilization of Folin−Ciocalteu phenol reagent for the detection of certain nitrogen compounds. J. Agric. Food Chem. 2003, 51, 1811–1815. [Google Scholar] [CrossRef]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. Thorough study of reactivity of various compound classes toward the Folin−Ciocalteu reagent. J. Agric. Food Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef] [Green Version]
- Musundire, R. Bio-active compounds composition in edible stinkbugs consumed in South-Eastern districts of Zimbabwe. Int. J. Biol. 2014, 6, 36–45. [Google Scholar] [CrossRef]
- Musundire, R.; Zvidzai, C.J.; Chidewe, C.; Ngadze, R.T.; Macheka, L.; Manditsera, F.A.; Mubaiwa, J.; Masheka, A. Nutritional and bioactive compounds composition of Eulepida mashona, an edible beetle in Zimbabwe. J. Insects Food Feed 2016, 2, 179–187. [Google Scholar] [CrossRef]
- Musundire, R.; Zvidzai, C.J.; Chidewe, C.; Samende, B.K.; Manditsera, F.A. Nutrient and anti-nutrient composition of Henicus whellani Orthoptera: Stenopelmatidae, an edible ground cricket, in south-eastern Zimbabwe. Int. J. Trop. Insect Sci. 2014, 34, 223–231. [Google Scholar] [CrossRef]
- Pyo, S.J.; Kang, D.G.; Jung, C.; Sohn, H.Y. Anti-thrombotic, anti-oxidant and haemolysis activities of six edible insect species. Foods 2020, 9, 401. [Google Scholar] [CrossRef]
- Hopkins, T.L.; Ahmad, S.A. Flavonoid wing pigments in grasshoppers. Experientia 1991, 47, 1089–1091. [Google Scholar] [CrossRef]
- Bernays, E.A.; Howard, J.J.; Champagne, D.; Estesen, B.J. Rutin: A phagostimulant for the polyphagous acridid Schistocerca americana. Entomol. Exp. Appl. 1991, 60, 19–28. [Google Scholar] [CrossRef]
- Hirayama, C.; Ono, H.; Meng, Y.; Shimada, T.; Daimon, T. Flavonoids from the cocoon of Rondotia menciana. Phytochem 2013, 94, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Kurioka, A.; Yamazaki, M. Purification and identification of flavonoids from the yellow green cocoon shell Sasamayu of the silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2002, 66, 1396–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, Y.; Nakajima, K.I.; Nagayasu, K.I.; Takabayashi, C. Flavonoid 5-glucosides from the cocoon shell of the silkworm, Bombyx mori. Phytochemistry 2002, 59, 275–278. [Google Scholar] [CrossRef]
- Fu, X.; Chai, C.L.; Li, Y.P.; Li, P.; Luo, S.H.; Li, Q.; Li, M.W.; Liu, Y.Q. Metabolomics reveals abundant flavonoids in edible insect Antheraea pernyi. J. Asia Pac. Entomol. 2021, 24, 711–715. [Google Scholar] [CrossRef]
- Wilson, A. Flavonoid pigments in marbled white butterfly Melanargia galathea are dependent on flavonoid content of larval diet. J. Chem. Ecol. 1985, 11, 1161–1179. [Google Scholar] [CrossRef]
- Morris, S.J.; Thomson, R.H. The flavonoid pigments of the small heath butterfly, Coenonympha pamphilus L. J. Insect Physiol. 1964, 10, 377–383. [Google Scholar] [CrossRef]
- Wilson, A. Flavonoid pigments in chalkhill blue Lysandra coridon Poda and other lycaenid butterflies. J. Chem. Ecol. 1987, 13, 473–493. [Google Scholar] [CrossRef]
- Burghardt, F.; Fiedlert, K.; Proksch, P. Uptake of flavonoids from Vicia villosa Fabaceae by the lycaenid butterfly, Polyommatus icarus Lepidoptera: Lycaenidae. Biochem. Syst. Ecol. 1997, 25, 527–536. [Google Scholar] [CrossRef]
- Morris, S.J.; Thomson, R.H. Flavonoid pigments in the marbled white butterfly (Melanargia Galathea Seitz). Tetrahedron Lett. 1963, 4, 101–104. [Google Scholar] [CrossRef]
- Vihakas, M.A.; Kapari, L.; Salminen, J.P. New types of flavonol oligoglycosides accumulate in the hemolymph of birch-feeding sawfly larvae. J. Chem. Ecol. 2010, 36, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Vihakas, M.; Tähtinen, P.; Ossipov, V.; Salminen, J.P. Flavonoid metabolites in the hemolymph of European pine sawfly Neodiprion sertifer larvae. J. Chem. Ecol. 2012, 38, 538–546. [Google Scholar] [CrossRef]
- Yoon, C.H.; Jeon, S.H.; Ha, Y.J.; Kim, S.W.; Bang, W.Y.; Bang, K.H.; Gal, S.W.; Kim, I.S.; Cho, Y.S. Functional chemical components in Protaetia brevitarsis larvae: Impact of supplementary feeds. Food Sci. Anim. Resour. 2020, 40, 461–473. [Google Scholar] [CrossRef]
- EU Regulation 2015/2283 on Novel Foods, Amending Regulation (EU) No 1169/2011 of the European Parliament and the Council and Repealing Regulation (EC) No 258/97 of the European Parliament and the Council and Commission Regulation (EC) No 1852/2001. Available online: https://eur-lex.europa.eu/legal-content/IT/TXT/?uri=CELEX:32015R2283 (accessed on 3 November 2021).
- Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Knutsen, H.K. Safety of partially defatted house cricket (Acheta domesticus) powder as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2022, 20, e07258. [Google Scholar]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Knutsen, H.K. Safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06343. [Google Scholar]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Knutsen, H.K. Safety of frozen and dried formulations from migratory locust (Locusta migratoria) as a Novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06667. [Google Scholar]
- EU Implementing Regulation 2017/2470. Commission Implementing Regulation (EU) 2017/2470 of 20 December 2017 Establishing the Union List of Novel Foods in Accordance with Regulation (EU) 2015/2283 of the European Parliament and the Council on Novel Foods. Available online: https://eur-lex.europa.eu/eli/reg_impl/2017/2470/oj (accessed on 3 November 2021).
- Tata, A.; Massaro, A.; Marzoli, F.; Miano, B.; Bragolusi, M.; Piro, R.; Belluco, S. Authentication of Edible Insects’ Powders by the Combination of DART-HRMS Signatures: The First Application of Ambient Mass Spectrometry to Screening of Novel Food. Foods 2022, 11, 2264. [Google Scholar] [CrossRef]
- Haber, M.; Mishyna, M.; Itzhak Martinez, J.J.; Benjamin, O. Edible larvae and pupae of honey bee (Apis mellifera): Odor and nutritional characterization as a function of diet. Food Chem. 2019, 292, 197–203. [Google Scholar] [CrossRef]
- Schiel, L.; Wind, C.; Krueger, S.; Braun, P.G.; Koethe, M. Applicability of analytical methods for determining the composition of edible insects in German Food Control. J. Food Compos. Anal. 2022, 112, 104676. [Google Scholar] [CrossRef]
- Di Mattia, C.; Battista, N.; Sacchetti, G.; Serafini, M. Antioxidant activities in vitro of water and Liposoluble Extracts obtained by different species of edible insects and invertebrates. Front. Nutr. 2019, 6, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasconcelos dos Santos, O.; Sodré Dias, P.C.; Dias Soares, S.; Vieira da Conceição, L.R.; Teixeira-Costa, B.E. Artisanal oil obtained from insects’ larvae (Speciomerus ruficornis): Fatty acids composition, physicochemical, nutritional and antioxidant properties for application in food. Eur. Food Res. Technol. 2021, 247, 1803–1813. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Giacintucci, V.; Di Mattia, C.; Sacchetti, G.; Neri, L.; Pittia, P. Role of olive oil phenolics in physical properties and stability of mayonnaise-like emulsions. Food Chem. 2016, 213, 369–377. [Google Scholar] [CrossRef]
- Hall, F.; Johnson, P.E.; Liceaga, A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. 2018, 262, 39–47. [Google Scholar] [CrossRef]
- Aiello, D.; Giglio, A.; Talarico, F.; Vommaro, M.L.; Tagarelli, A.; Napoli, A. Mass Spectrometry-Based Peptide Profiling of Haemolymph from Pterostichus melas Exposed to Pendimethalin Herbicide. Molecules 2022, 27, 4645. [Google Scholar] [CrossRef]
- Bongiorno, D.; Di Stefano, V.; Indelicato, S.; Avellone, G.; Ceraulo, L. Bio-phenols determination in olive oils: Recent mass spectrometry approaches. Mass Spectrom. Rev. 2021, e21744. [Google Scholar] [CrossRef]
- Indelicato, S.; Bongiorno, D.; Indelicato, S.; Drahos, L.; Turco Liveri, V.; Turiák, L.; Vekey, K.; Ceraulo, L. Degrees of freedom effect on fragmentation in tandem mass spectrometry of singly charged supramolecular aggregates of sodium sulfonates. J. Mass Spectrom. 2013, 48, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Bongiorno, D.; Indelicato, S.; Giorgi, G.; Scarpella, S.; Liveri, V.T.; Ceraulo, L. Electrospray ion mobility mass spectrometry of positively charged sodium bis(2-ethylhexyl)sulfosuccinate aggregates. Eur. J. Mass Spectrom. 2014, 20, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Francesca, N.; Barbera, M.; Martorana, A.; Saiano, F.; Gaglio, R.; Aponte, M.; Moschetti, G.; Settanni, L. Optimised method for the analysis of phenolic compounds from caper (Capparis spinosa L.) berries and monitoring of their changes during fermentation. Food Chem. 2016, 196, 1172–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bongiorno, D.; Ceraulo, L.; Indelicato, S.; Turco Liveri, V.; Indelicato, S. Charged supramolecular assemblies of surfactant molecules in gas phase. Mass Spectrom. Rev. 2016, 35, 170–187. [Google Scholar] [CrossRef] [PubMed]
- Di Donna, L.; Bartella, L.; De Vero, L.; Gullo, M.; Giuffrè, A.M.; Zappia, C.; Capocasale, M.; Poiana, M.; D’Urso, S. Vinegar production from Citrus bergamia by-products and preservation of bioactive compounds. Eur. Food Res. Technol. 2020, 246, 1981–1990. [Google Scholar] [CrossRef]
- Mazzotti, F.; Bartella, L.; Talarico, I.R.; Napoli, A.; Di Donna, L. High-throughput determination of flavanone-O-glycosides in citrus beverages by paper spray tandem mass spectrometry. Food Chem. 2021, 360, 130060. [Google Scholar] [CrossRef]
- Bartella, L.; Mazzotti, F.; Talarico, I.R.; De Luca, G.; Santoro, I.; Prejanò, M.; Riccioni, C.; Marino, T.; Di Donna, L. Structural Characterization of Peripolin and Study of Antioxidant Activity of HMG Flavonoids from Bergamot Fruit. Antioxidants 2022, 11, 1847. [Google Scholar] [CrossRef]
- Argo, A.; Bongiorno, D.; Bonifacio, A.; Pernice, V.; Liotta, R.; Indelicato, S.; Zerbo, S.; Fleres, P.; Ceraulo, L.; Procaccianti, P. A fatal case of a paint thinner ingestion: Comparison between toxicological and histological findings. Am. J. Forensic Med. Pathol. 2010, 31, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Trujillo, M.; Gómez-Caravaca, A.M.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Parella, T. Separation and identification of phenolic compounds of extra virgin olive oil from Olea europaea L. by HPLC-DAD-SPE-NMR/MS. Identification of a new diastereoisomer of the aldehydic form of oleuropein aglycone. J. Agric. Food Chem. 2010, 58, 9129–9136. [Google Scholar] [CrossRef]
- Castiglione, F.; Appetecchi, G.B.; Passerini, S.; Panzeri, W.; Indelicato, S.; Mele, A. Multiple points of view of heteronuclear NOE: Long range vs short range contacts in pyrrolidinium based ionic liquids in the presence of Li salts. J. Mol. Liq. 2015, 210, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Rodríguez, G.; Marina, M.L.; Plaza, M. Strategies for the extraction and analysis of non-extractable polyphenols from plants. J. Chromatogr. A 2017, 1514, 1–15. [Google Scholar] [CrossRef]
- Sugiki, T.; Furuita, K.; Fujiwara, T.; Kojima, C. Current NMR techniques for structure-based drug discovery. Molecules 2018, 23, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.M.; Zhu, H.J.; Zhou, F.J.; Tu, Z.C.; Cheng, Y.X. Phenolic compounds from the insect Blaps japanensis with inhibitory activities towards cancer cells, COX-2, ROCK1 and JAK3. Tetrahedron 2019, 75, 1029–1033. [Google Scholar] [CrossRef]
- Harborne, J.B. Flavonoid pigments. In Herbivores: Their Interactions with Secondary Plant Metabolites, 2nd ed.; Rosenthal, G.A., Berenbaum, M.R., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 389–429. [Google Scholar]
- Schittko, U.; Burghardt, F.; Fiedler, K.; Wray, V.; Proksch, P. Sequestration and distribution of flavonoids in the common blue butterfly Polyommatus icarus reared on Trifolium repens. Phytochemistry 1999, 51, 609–614. [Google Scholar] [CrossRef]
- Salminen, J.P.; Lempa, K. Effects of hydrolysable tannins on a herbivorous insect: Fate of individual tannins in insect digestive tract. Chemoecology 2002, 12, 203–211. [Google Scholar] [CrossRef]
- Ferreres, F.; Fernandes, F.; Oliveira, J.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Metabolic profiling and biological capacity of Pieris brassicae fed with kale (Brassica oleracea L. var. acephala). Food Chem. Toxicol. 2009, 47, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, F.; Proksch, P.; Fiedler, K. Flavonoid sequestration by the common blue butterfly Polyommatus icarus: Quantitative intraspecific variation in relation to larval hostplant, sex and body size. Biochem. Syst. Ecol. 2001, 29, 875–889. [Google Scholar] [CrossRef]
- Sang, S.; Lapsley, K.; Jeong, W.-S.; Lachance, P.A.; Ho, C.T.; Rosen, R.T. Antioxidative phenolic compounds isolated from almond skins (Prunus amygdalus Batsch). J. Agric. Food Chem. 2002, 50, 2459–2463. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Ghosh, A.; Ray, M.; Gangopadhyay, D. Evaluation of proximate composition and antioxidant properties in silk-industrial byproduct. Food Sci. Technol. 2020, 132, 109900. [Google Scholar] [CrossRef]
- Wannee, S.; Luchai, B. 1-Deoxynojirimycin and polyphenolic composition and antioxidant activity of different native Thai silkworm (Bombyx mori) larvae. J. King Saud Univ. Sci. 2020, 32, 2762–2766. [Google Scholar] [CrossRef]
- Deori, M.; Boruah, D.C.; Devi, D.; Devi, R. Antioxidant and antigenotoxic effects of pupae of the MUGA silkworm Antheraea assamensis. Food Biosci. 2014, 5, 108–114. [Google Scholar] [CrossRef]
- Sugumaran, M. Chemistry of cuticular sclerotization. Adv. Insect Physiol. 2010, 39, 151–209. [Google Scholar]
- Whitten, M.M.A.; Coates, C.J. Re-evaluation of insect melanogenesis research: Views from the dark side. Pigment Cell Melanoma Res. 2017, 30, 386–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söderhäll, K. Prophenoloxidase activating system and melanization—A recognition mechanism of arthropods? A review. Dev. Comp. Immunol. 1982, 6, 601–611. [Google Scholar] [PubMed]
- Cerenius, L.; Söderhäll, K. Immune properties of invertebrate phenoloxidases. Dev. Comp. Immunol. 2021, 122, 104098. [Google Scholar] [CrossRef]
- Bell, K.; Smith, V.J. In vitro superoxide production by hyaline cells of the shore crab Carcinus maenas (L.). Dev. Comp. Immunol. 1993, 17, 211–219. [Google Scholar] [CrossRef]
- Anderson, R.S. Hemocyte-derived reactive oxygen intermediate production in four bivalve mollusks. Dev. Comp. Immunol. 1994, 18, 89–96. [Google Scholar] [CrossRef]
- Valembois, P.; Lassègues, M. In vitro generation of reactive oxygen species by free coelomic cells of the annelid Eisenia fetida andrei: An analysis by chemiluminescence and nitro blue tetrazolium reduction. Dev. Comp. Immunol. 1995, 19, 195–204. [Google Scholar] [CrossRef]
- Riley, P.A. Radicals in melanin biochemistry. Ann. N. Y. Acad. Sci. 1988, 551, 111–119. [Google Scholar] [CrossRef]
- Salvatore, L.; Gallo, N.; Aiello, D.; Lunetti, P.; Barca, A.; Blasi, L.; Madaghiele, M.; Bettini, S.; Giancane, G.; Hasan, M.; et al. An Insight on Type I Collagen from Horse Tendon for the Manufacture of Implantable Devices. Int. J. Biol. Macromol. 2020, 154, 291–306. [Google Scholar] [CrossRef]
- O’Brien, P.J. Molecular mechanisms of quinone cytotoxicity. Chem. Biol. Interact. 1991, 80, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Smit, N.P.M.; Peters, K.; Menko, W.; Westerhof, W.; Pavel, S.; Riley, P.A. Cytotoxicity of a selected series of substituted phenols towards cultured melanoma cells. Melanoma Res. 1996, 2, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Nappi, A.J.; Vass, E. Melanogenesis and the generation of cytotoxic molecules during insect cellular immune reactions. Pigment Cell Res. 1993, 6, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Cotelle, N.; Moreau, S.; Cotelle, P.; Catteau, J.P.; Bernier, J.L.; Henicart, J.P. Generation of free radicals by simple prenylated hydroquinone derivativs, natural antitumor agents from the marine urochordate Aplidium californicum. Chem. Res. Toxicol. 1991, 4, 300–305. [Google Scholar] [CrossRef] [PubMed]


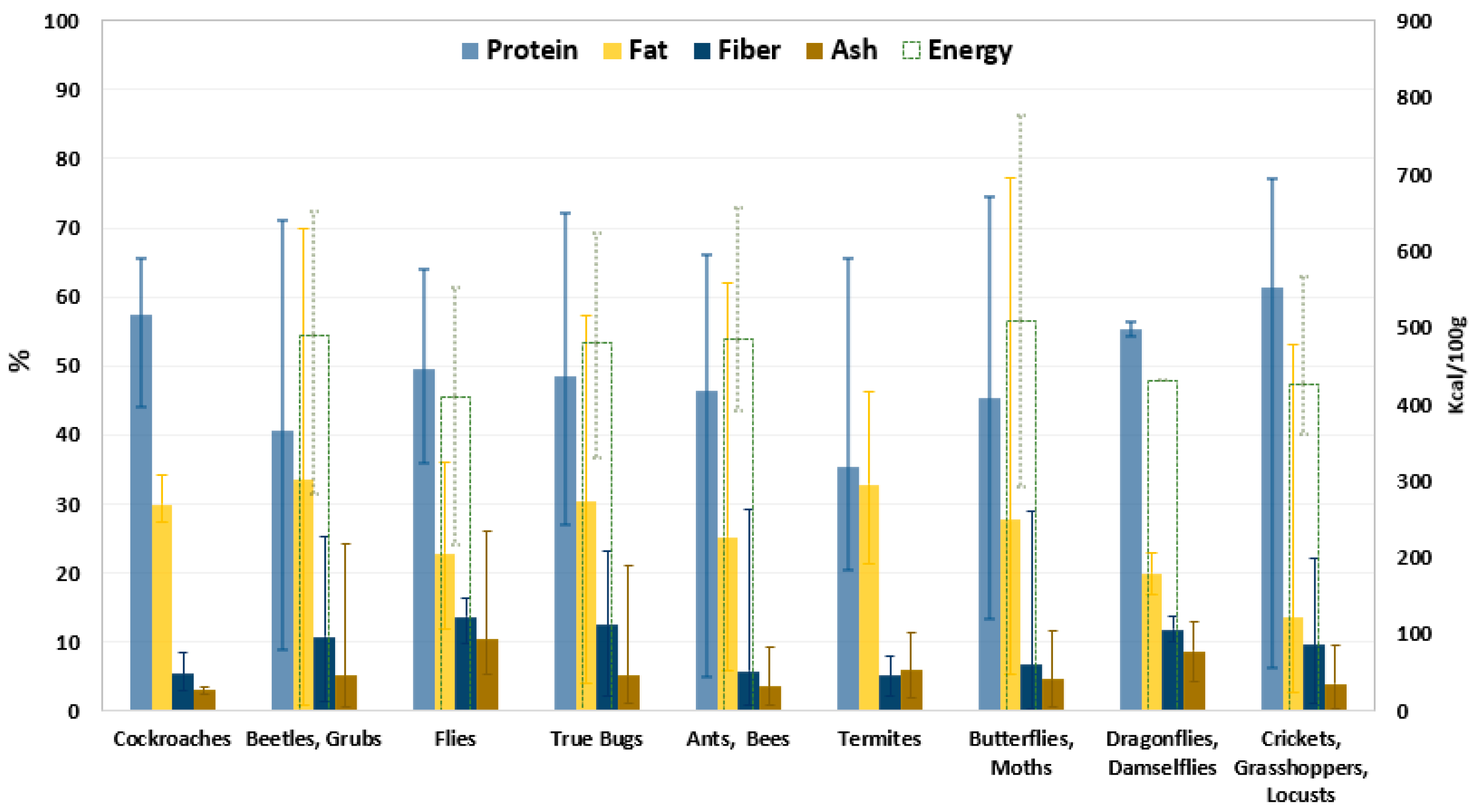{kind=link}
| Chicken | Pork | Beef | Insects | Reference | |
|---|---|---|---|---|---|
| Water L/kg meat | 2300 | 3500–22,000 | 43,000 | 40 | [8,29] |
| Feed kg/kg live animal | 2.5 | 5 | 10 | 1.7 | [30] |
| Land ha/kg protein | 2–3.5 | 2–3.5 | 10 | 1 | [31] |
| CH4 emissions g/kg biomass | n.r 1 | 1.9 | 114 | 0.1 | [32,33] |
| CO2 emissions g/kg biomass | n.r 1 | 79.6 | 285 | 7.6 | [32,33] |
| Insects | gGAE/100 g | Reference |
|---|---|---|
| Stink bugs (Encosternum delegorguei) | 3.6 | [17,75] |
| Cricket (Henicus whellani) | 0.08 | [17] |
| Cricket (Henicus whellani) | 0.77 | [27] |
| Beetle (Eulepida mashona) | 0.08 | [76] |
| Ground cricket (Henicus whellani) | 0.77 | [77] |
| Stinkbugs (Encosternum delegorguei) | 3.6 | [17,75] |
| House cricket (Acheta domesticus) | 0.3–5.0 | [24] |
| Chafer beetles (Holotrichia parallela) | 5 g | [14] |
| Beetle (Eulepida mashona) | 0.08 | [27] |
| Rhinoceros beetle (Allomyrina dichotoma) | 0.13 | [78] |
| Mealworm beetle (Tenebrio molitor) | 0.26 | [78] |
| Scarabaeidae (Protaetia brevitarsis) | 1.18 | [78] |
| Cricket (Gryllus bimaculatus) | 1.56 | [78] |
| Cricket (Teleogryllus emma) | 1.55 | [78] |
| Bee (Apis mellifera) | 1.24 | [78] |
| Family/Species | Polyphenolic Compounds | Reference |
|---|---|---|
| Black ant | ||
| Polyrhachis vicina | Salicylic acid, trans-cinnamic acid, vanillic acid, isoferulic acid, gallic acid, 3,4-dihydroxybenzoic acid, formononetin, liquiritigenin, quercetin, caffeic acid, naringenin, catechin, sakuranetin, and L-epicatechin. | [22] |
| Acrididae | ||
| Acheta domesticus | Quinic acid, gallic acid, 4-hydroxybenzoic acid, chlorogenic acid, caffeic acid, syringic acid, p-coumaric acid, ferulic acid, sinapic acid, 2-hydroxybenzoic acid, daidzein, quercetin, naringenin, and apigenin | [16] |
| Dissoteira carolina | Quercetin (3,3′,4′,5,7-pentahydroxyflavone) and quercetin-β-3-O-glucoside | [79] |
| Schistocerca americana | Luteolin (3′,4′,5,7- tetrahydroxyflavone) and β-3-O-glucoside | [80] |
| Beetle | ||
| Holotrichia parallela | Resveratrol, 4-hydroxyacetophenone, protocatechualdehyde, ferulic acid, gallic acid, protocatechuic acid, epicatechin, quercetin, and catechin | [14] |
| Caterpillar | ||
| Rondotia menciana | Quercetin-glycosides and kaempferol-glycosides | [81] |
| Silkworm | ||
| Bombyx mori | Quercetin, kaempferol, and quercetin-glycosides | [82,83] |
| Butterfly | ||
| Antheraea pernyi | Hyperoside (quercetin 3-O-glucoside), isoquercitroside, tricin 7-O-hexoside, hesperetin 5-O-glucoside, protocatechuic acid, luteolin 7-O-glucoside (cynaroside), kaempferol 3-O-glucoside (astragalin), C-hexosyl-luteolin O-p-coumaroylhexoside, luteolin 6-C-glucoside, tricin 4′-O-(β-guaiacylglyceryl) ether O-hexoside, orientin, luteolin C-hexoside, kaempferol 3-O-galactoside (trifolin), and tricin 4′-O-(β-guaiacylglyceryl) ether 7-O-hexoside 1 | [84] |
| Melanargia galathea | Tricin (4′,5,7-trihydroxy-3′,5′-dimethoxyflavone), apigenin (4,5,7-trihydroxyflavone), tricin 7-glucoside, orientin (8-glucosylluteolin), luteolin 7-diglucoside, orientin 7-glucoside, vitexin 7-glucoside, isoorientin (luteolin 6-C-glucoside), isovitexin (6-C-glucosylapigenin), and tricin 4′-conjugate | [85] |
| Coenonympha pamphilus | Tricin (4′,5,7-trihydroxy-3′,5′-dimethoxyflavone) | [86] |
| Lysandra coridon Poda | Kaempferol-glycosides | [87] |
| Polyommatus icarus | Quercetin, kaempferol, and quercetin-glycosides and kaempferol-glycosides | [88] |
| Pieris brassicae | Kaempferol glycosides and ferulic and sinapic acids | [79] |
| Melanargia galathea | Glycosides of tricin (tricin-glycosides), lutexin, and tricin (4′,5,7-trihydroxy-3′,5′-dimethoxyflavone) | [89] |
| Hymenoptera | ||
| Amauronematus amplus, Arge sp. , Nematus bre-vivalvis, Nematus pravus, Nematus viridis, Nematus alpestris, Trichiosoma scalesii | Quercetin-glycosides, kaempferol-glycosides | [90] |
| Neodiprion sertifer | (+)-Catechin 7-O-β-glucoside, isorhamnetin 3,7,4′-tri-O-β-glucosid, kaempferol 3,7, 4′-tri-O-β-glucoside, kaempferol 3,7, 4′-tri-O-β-glucoside, and quercetin 3,7,4′-tri-O-β-glucoside | [91] |
| Insect Species | Most Abundant Molecules | Activity | References |
|---|---|---|---|
| P. vicina | salicylic acid, gallic acid, liquiritigenin | PL inhibitory activity | Zhang et al. [22] |
| A. assamensis | not specified | antityrosinase activity | Deori et al. [131] |
| B. mori | not specified | antigenotoxic activity | Deori et al. [131] |
| B. japanensis | blapsins | anticancer activity, inhibitory activities versus the JAK3 family genes; oncological, cardiovascular, and neurological disorders, organ transplantation, and autoimmune diseases | Yan et al. [121] |
| Macrotermes facilger | polyphenols | antioxidant and antimicrobial properties; | Kunatsa et al. [41] |
| Macrotermes facilger | oxalates, tannins, and cyanogenic glycosides | antinutritional properties | Kunatsa et al. [41] |
| Henicus whellani | polyphenols | antioxidant and antimicrobial properties; | Kunatsa et al. [41] |
| Henicus whellani | oxalates, tannins, and cyanogenic glycosides | antinutritional properties | Kunatsa et al. [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aiello, D.; Barbera, M.; Bongiorno, D.; Cammarata, M.; Censi, V.; Indelicato, S.; Mazzotti, F.; Napoli, A.; Piazzese, D.; Saiano, F. Edible Insects an Alternative Nutritional Source of Bioactive Compounds: A Review. Molecules 2023, 28, 699. https://doi.org/10.3390/molecules28020699
Aiello D, Barbera M, Bongiorno D, Cammarata M, Censi V, Indelicato S, Mazzotti F, Napoli A, Piazzese D, Saiano F. Edible Insects an Alternative Nutritional Source of Bioactive Compounds: A Review. Molecules. 2023; 28(2):699. https://doi.org/10.3390/molecules28020699
Chicago/Turabian StyleAiello, Donatella, Marcella Barbera, David Bongiorno, Matteo Cammarata, Valentina Censi, Serena Indelicato, Fabio Mazzotti, Anna Napoli, Daniela Piazzese, and Filippo Saiano. 2023. "Edible Insects an Alternative Nutritional Source of Bioactive Compounds: A Review" Molecules 28, no. 2: 699. https://doi.org/10.3390/molecules28020699
APA StyleAiello, D., Barbera, M., Bongiorno, D., Cammarata, M., Censi, V., Indelicato, S., Mazzotti, F., Napoli, A., Piazzese, D., & Saiano, F. (2023). Edible Insects an Alternative Nutritional Source of Bioactive Compounds: A Review. Molecules, 28(2), 699. https://doi.org/10.3390/molecules28020699