
Chemical Composition, Antibacterial Properties, and Anti-Enzymatic Effects of Eucalyptus Essential Oils Sourced from Tunisia


 , ,
, ,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Yields and Chemical Composition
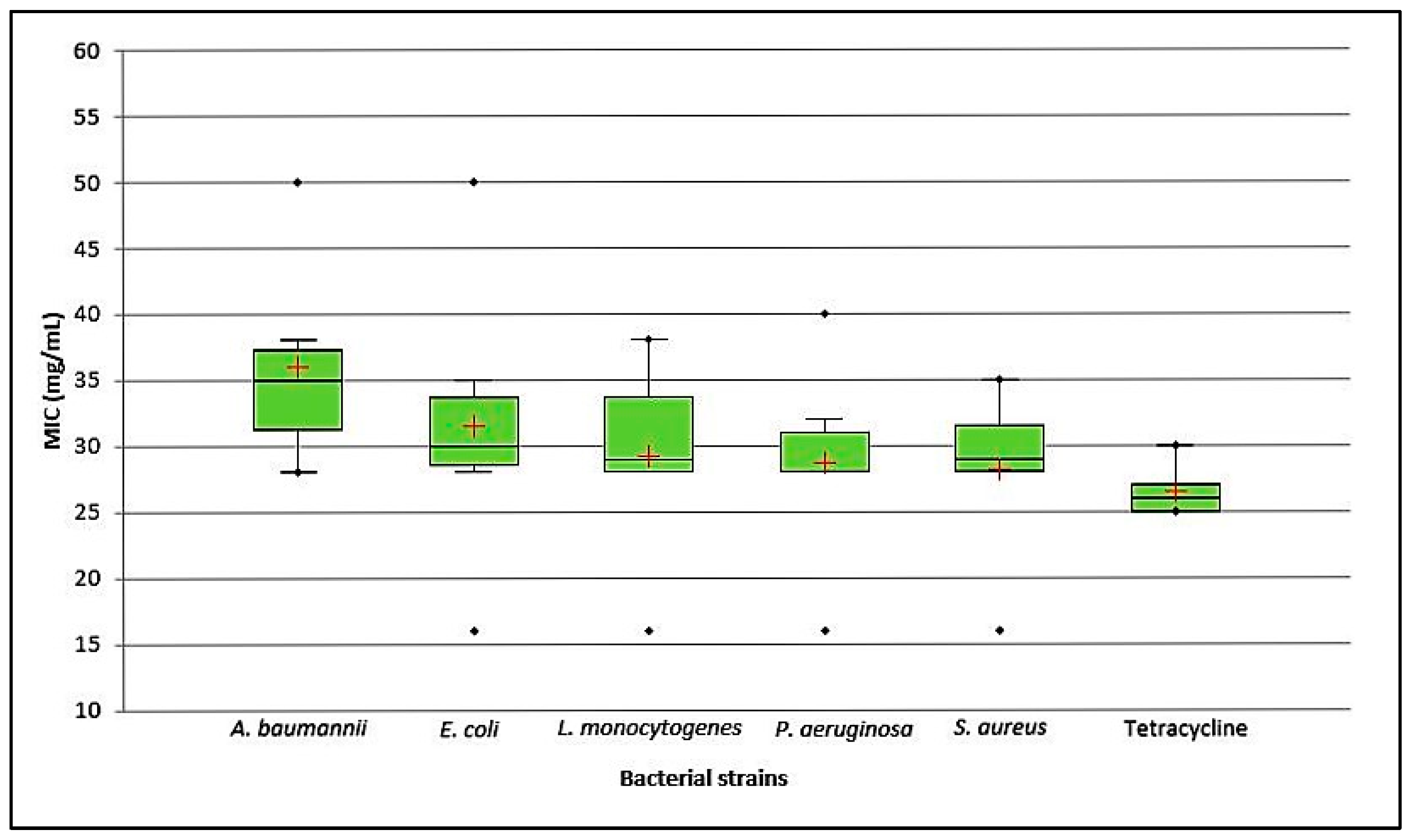
2.2. Antibiofilm Activity
2.3. Anti-Enzymatic Activity
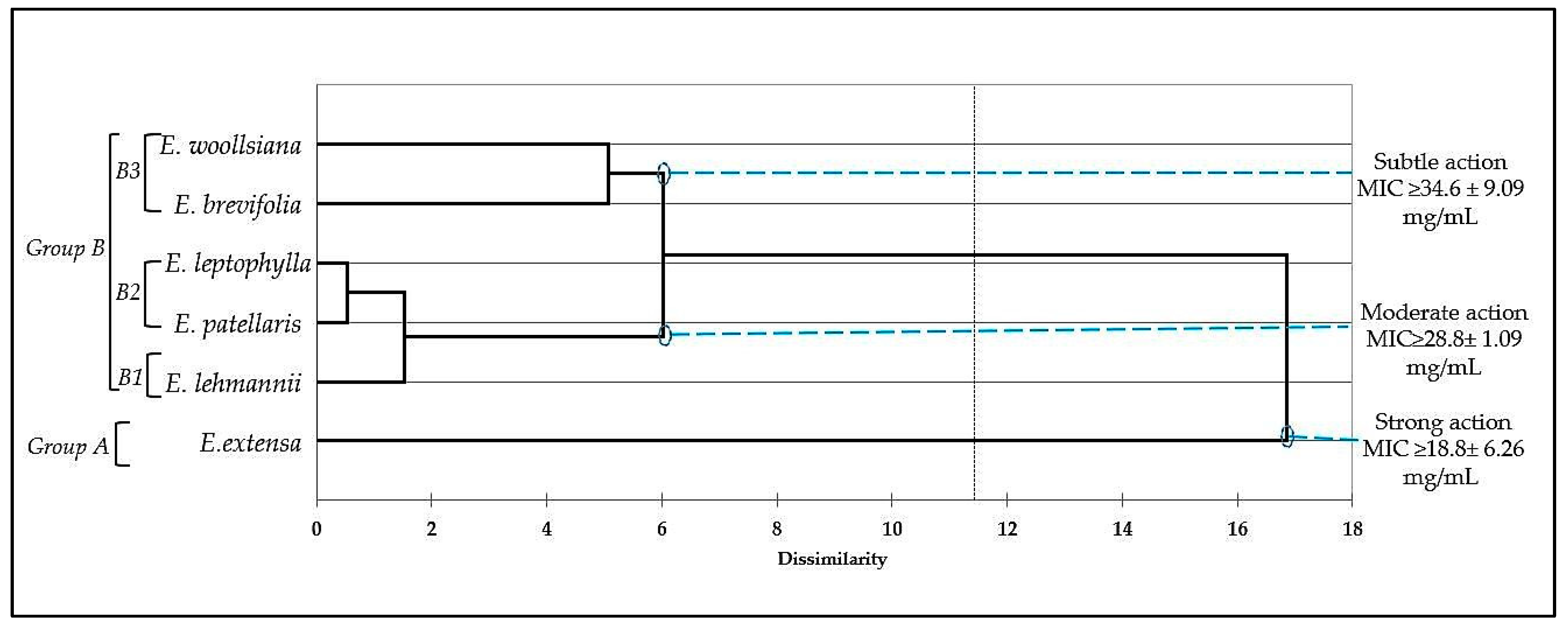
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Isolation and Analysis of the Essential Oils
4.3. Antimicrobial Activity
4.3.1. Microorganisms and Culture Conditions
4.3.2. Minimal Inhibitory Concentration (MIC)
4.3.3. Biofilm Inhibitory Activity
4.3.4. Effects of EOs on Cell Metabolic Activity within Biofilm
4.4. Anti-Enzymatic Activity
4.4.1. Cholinesterases Inhibition
4.4.2. α-Amylase Inhibition Assay
4.4.3. α-Glucosidase Inhibition Assay
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Moloney, M.G. Natural Products as a Source for Novel Antibiotics. Trends Pharmacol. Sci. 2016, 37, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Ghalib, R.M.; Sasikala, P.; Ahmed, K.K.M. Cholinesterase inhibitors from botanicals. Pharmacogn. Rev. 2013, 7, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Savelev, S.U.; Okello, E.J.; Perry, E.K. Butyryl- and acetyl-cholinesterase inhibitory activities in essential oils of Salvia species and their constituents. Phytother. Res. 2004, 18, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Loizzo, M.R.; Menichini, F. Natural products as alpha-amylase and alpha-glucosidase inhibitors and their hypoglycaemic potential in the treatment of diabetes: An update. Mini Rev. Med. Chem. 2010, 10, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Dhakad, A.K.; Pandey, V.V.; Beg, S.; Rawat, J.M.; Singh, A. Biological, medicinal and toxicological significance of Eucalyptus leaf essential oil: A review. J. Sci. Food Agric. 2018, 98, 833–848. [Google Scholar] [CrossRef]
- Polito, F.; Fratianni, F.; Nazzaro, F.; Amri, I.; Kouki, H.; Khammssi, M.; Hamrouni, L.; Malaspina, P.; Cornara, L.; Khedhri, S.; et al. Essential Oil Composition, Antioxidant Activity and Leaf Micromorphology of Five Tunisian Eucalyptus Species. Antioxidants 2023, 12, 867. [Google Scholar] [CrossRef]
- Dey, B.; Mitra, A. Chemo-profiling of eucalyptus and study of its hypoglycemic potential. World J. Diabetes 2013, 4, 170–176. [Google Scholar] [CrossRef]
- Khedhri, S.; Polito, F.; Caputo, L.; Manna, F.; Khammassi, N.; Hamrouni, M.; Amri, I.; Nazzaro, F.; De Feo, V.; Fratianni, F. Chemical Composition, Phytotoxic and Antibiofilm Activity of Seven Eucalyptus Species from Tunisia. Molecules 2022, 27, 8227. [Google Scholar] [CrossRef]
- Elaissi, A.; Salah, K.H.; Mabrouk, S.; Larbi, K.M.; Chemi, R.; Harzallah-Skhiri, F. Antibacterial activity and chemical composition of 20 Eucalyptus species’ essential oils. Food Chem. 2011, 129, 1427–1434. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Kaur, S. Eucalyptus essential oil as a natural pesticide. For. Ecol. Manag. 2008, 256, 2166–2174. [Google Scholar] [CrossRef]
- Caputo, L.; Smeriglio, A.; Trombetta, D.; Cornara, L.; Trevena, G.; Valussi, M.; Fratianni, F.; De Feo, V.; Nazzaro, F. Chemical Composition and Biological Activities of the Essential Oils of Leptospermum petersonii and Eucalyptus gunnii. Front. Microbiol. 2020, 11, 409. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.G.; Gago, C.; Antunes, M.D.; Lagoas, S.; Faleiro, M.L.; Megías, C.; Figueiredo, A.C. Antibacterial, antioxidant, and antiproliferative activities of Corymbia citriodora and the essential oils of eight Eucalyptus species. Medicines 2018, 5, 61. [Google Scholar] [CrossRef]
- Polito, F.; Kouki, H.; Khedhri, S.; Hamrouni, L.; Mabrouk, Y.; Amri, I.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical Composition and Phytotoxic and Antibiofilm Activity of the Essential Oils of Eucalyptus bicostata, E. gigantea, E. intertexta, E. obliqua, E. pauciflora and E. tereticornis. Plants 2022, 11, 3017. [Google Scholar] [CrossRef] [PubMed]
- Basak, S.S.; Candan, F. Chemical composition and in vitro antioxidant and antidiabetic activities of Eucalyptus camaldulensis Dehnh. Essential oil. J. Iran. Chem. Soc. 2010, 7, 216–226. [Google Scholar] [CrossRef]
- Aazza, S.; Lyoussi, B.; Miguel, M.G. Antioxidant and Antiacetylcholinesterase Activities of Some Commercial Essential Oils and Their Major Compounds. Molecules 2011, 16, 7672–7690. [Google Scholar] [CrossRef]
- Kobenan, K.C.; Ochou, G.E.C.; Kouadio, I.S.; Kouakou, M.; Bini, K.K.N.; Ceylan, R.; Zrngn, G.; Kosso Boka, N.R.; Ochou, O.G. Chemical Composition, Antioxidant Activity, Cholinesterase Inhibitor and in Vitro Insecticidal Potentiality of Essential Oils of Lippia multiflora Moldenke and Eucalyptus globulus Labill. on the Main Carpophagous Pests of Cotton Plant in Ivory Coast. Chem. Biodivers. 2022, 19, e202100993. [Google Scholar] [CrossRef]
- Barra, A. Factors Affecting Chemical Variability of Essential Oils: A Review of Recent Developments. Nat. Prod. Commun. 2009, 4, 1147–1154. [Google Scholar] [CrossRef]
- Elaissi, A.; Chraif, I.; Bannour, F.; Farhat, F.; Ben Salah, M.; ChemLi, R.; Khouja, M.L. Contribution to the Qualitative and Quantitative Study of Seven Eucalyptus Species Essential Oil Harvested of Hajeb’s Layoun Arboreta (Tunisia). J. Essent. Oil Bear. Plants 2007, 10, 15–25. [Google Scholar] [CrossRef]
- Ben Slimane, B.; Ezzine, O.; Dhahri, S.; Mohamed Lahbib, B.J. Essential oils from two Eucalyptus from Tunisia and their insecticidal action on Orgyia trigotephras (Lepidotera, Lymantriidae). Biol. Res. 2014, 47, 29. [Google Scholar] [CrossRef]
- Mediouni, J.; Mediouni Ben Jemâa, J.; Haouel, S.; Bouaziz, M.; Khouja, M.L. Seasonal variations in chemical composition and fumigant activity of five Eucalyptus essential oils against three moth pests of stored dates in Tunisia. J. Stored Prod. Res. 2011, 48, 61–67. [Google Scholar] [CrossRef]
- Ben Hassine, D.; Rahmani, R.; Souchard, J.; Abderrabba, M.; Bouajila, J. Eucalyptus brevifolia F. Muell and Eucalyptus stricklandii Maiden leaves extracts: HPLC-DAD, GC-MS analysis and in vitro biological activities, combined with the principal component analysis. S. Afr. J. Bot. 2022, 147, 826–839. [Google Scholar] [CrossRef]
- Ben Hassine, D.; Kammoun El Euch, S.; Souchard, J.P.; Cazaux, S.; Abderrabba, M.; Bouajila, J. Phytochemical study and pharmaceutical properties of essential oils and organic extracts of two Eucalyptus species: E. stricklandii Maiden and E. brevifolia F. Muell. J. Essent. Oil Res. 2022, 34, 339–351. [Google Scholar] [CrossRef]
- Elaissi, A.; Rouis, Z.; Ben Salem, N.A.; Mabrouk, S.; ben Salem, Y.; Bel haj Salah, K.; Aouni, M.; Farhat, F.; ChemLi, R.; Harzallah-Skhiri, F.; et al. Chemical composition of 8 eucalyptus species’ essential oils and the evaluation of their antibacterial, antifungal and antiviral activities. BMC Complement. Altern. Med. 2012, 12, 81. [Google Scholar] [CrossRef]
- Yangui, I.; Boutiti, M.; Boussaid, M.; Messaoud, C. Essential Oils of Myrtaceae Species Growing Wild in Tunisia: Chemical Variability and Antifungal Activity Against Biscogniauxia mediterranea, the Causative Agent of Charcoal Canker. Chem. Biodivers. 2017, 14, e1700058. [Google Scholar] [CrossRef]
- Bignell, C.M.; Dunlop, P.J.; Brophy, J.J. Volatile leaf oils of some south-western and southern Australian species of the genus Eucalyptus (series I). Part XVI: Subgenus Symphyomyrtus, Section Bisectaria, SeriesCneorifoliae, Series Porantherae and Series Falcatae. Flavour Fragr. J. 1997, 12, 261–267. [Google Scholar] [CrossRef]
- Bignell, C.M.; Dunlop, P.J.; Brophy, J.J.; Jackson, J.F. Volatile leaf oils of some Queensland and northern australian species of the Genus Eucalyptus (series ii). Set off. Subgenus Symphyomyrtus, section Adnataria:(a) series Pliganthae,(b) series Ochrophloiae,(c) series Moluccanae,(d) series Polyanthemae,(e) series Paniculatae,(f) series Melliodorae and (g) series Porantheroideae. Flavour Fragr. J. 1997, 12, 19–27. [Google Scholar]
- Angioni, A.; Barra, A.; Cereti, E.; Barile, D.; Coisson, D.; Arlorio, M.; Dessi, M.; Coroneo, V.; Cabras, P. Chemical Composition, Plant Genetic Differences, Antimicrobial and Antifungal Activity Investigation of the Essential Oil of Rosmarinus officinalis L. J. Agric. Food Chem. 2004, 52, 3530–3535. [Google Scholar] [CrossRef] [PubMed]
- Echeverrigaray, S.; Agostini, G.; Atti-Serfini, N.; Paroul, L.; Pauletti, G.; Santos, A. Correlation between the Chemical and Genetic Relationships among Commercial Thyme Cultivars. J. Agric. Food Chem. 2001, 49, 4220–4223. [Google Scholar] [CrossRef]
- Adnan, M.; Siddiqui, A.J.; Hamadou, W.S.; Ashraf, S.A.; Hassan, M.I.; Snoussi, M.; Badraoui, R.; Jamal, A.; Bardacki, F.; Awadelkareem, A.M.; et al. Functional and Structural Characterization of Pediococcus pentosaceus-Derived Biosurfactant and Its Biomedical Potential against Bacterial Adhesion, Quorum Sensing, and Biofilm Formation. Antibiotics 2021, 10, 1371. [Google Scholar] [CrossRef]
- Khammassi, M.; Polito, F.; Amri, I.; Khedhri, S.; Hamrouni, L.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical Composition and Phytotoxic, Antibacterial and Antibiofilm Activity of the Essential Oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii. Molecules 2022, 27, 5820. [Google Scholar] [CrossRef]
- Limam, H.; Jemaa, M.B.; Tammar, S.; Ksibi, N.; Khammassi, S.; Jallouli, S.; Del Re, G.; Msaada, K. Variation in chemical profile of leaves essential oils from thirteen Tunisian Eucalyptus species and evaluation of their antioxidant and antibacterial properties. Ind. Crops Prod. 2020, 158, 112964. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Bassolé, I.H.N.; Juliani, H.R. Essential Oils in Combination and Their Antimicrobial Properties. Molecules 2012, 17, 3989–4006. [Google Scholar] [CrossRef] [PubMed]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial activity of some essential oils—Present status and future perspectives. Medicines 2017, 4, 58–79. [Google Scholar] [CrossRef]
- LaSarre, B.; Federle, M.J. Exploiting Quorum Sensing to Confuse Bacterial Pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef]
- Mączka, W.; Duda-Madej, A.; Górny, A.; Grabarczyk, M.; Wińska, K. Can Eucalyptol Replace Antibiotics? Molecules 2021, 26, 4933. [Google Scholar] [CrossRef]
- Masyita, A.; Sari, R.M.; Astuti, A.D.; Yasir, B.; Rumata, N.R.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef]
- Martins, C.D.M.; do Nascimento, E.A.; de Morais, S.A.; de Oliveira, A.; Chang, R.; Cunha, L.C.; Martins, M.M.; Martins, C.H.; Moraes, T.D.S.; Rodrigues, P.V.; et al. Chemical Constituents and Evaluation of Antimicrobial and Cytotoxic Activities of Kielmeyera coriacea Mart. & Zucc. Essential Oils. Evid. Based Complement. Alternat. Med. 2015, 2015, 842047. [Google Scholar] [CrossRef]
- Sareedenchai, V. Screening of acetylcholinesterase inhibitory activity in essential oil from Myrtaceae. Thai J. Pharm. Sci. 2019, 43, 63–68. [Google Scholar]
- Kiendrebeogo, M.; Coulibaly, A.Y.; Nebie, R.C.H.; Zeba, B.; Lamien, C.E.; Lamien-Meda, A.; Nacoulma, O.G. Antiacetylcholinesterase and antioxidant activity of essential oils from six medicinal plants from Burkina Faso. Rev. Bras. Farmacogn. 2011, 21, 63–69. [Google Scholar] [CrossRef]
- Chamali, S.; Bouajila, J.; Ben Daoud, H.; Camy, S.; Saadaoui, E.; Condoret, J.-S.; Romdhane, M. Variation in chemical composition and biological properties of two Tunisian Eucalyptus essential oils under three eco-friendly extraction techniques. J. Essent. Oil. Res. 2022, 34, 36–53. [Google Scholar] [CrossRef]
- Caputo, L.; Capozzolo, F.; Amato, G.; De Feo, V.; Fratianni, F.; Vivenzio, G.; Nazzaro, F. Chemical composition, antibiofilm, cytotoxic, and anti-acetylcholinesterase activities of Myrtus communis L. leaves essential oil. BMC Complement. Med. Ther. 2022, 22, 142. [Google Scholar] [CrossRef] [PubMed]
- Goodner, K.L. Practical retention index models of OV-101, DB-1, DB-5, and DB-Wax for flavor and fragrance compounds. LWT-Food Sci. Technol. 2008, 41, 951–958. [Google Scholar] [CrossRef]
- Jennings, W.; Shibamoto, T. Qualitative Analyisis of Flavour and Fragrance Volatiles by Glass Capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980; ISBN 9780323141055. [Google Scholar]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Polito, F.; Amato, G.; Caputo, L.; Francolino, R.; D’Acierno, A.; Fratianni, F.; Candido, V.; Coppola, R.; De Feo, V. Chemical composition of essential oils of bulbs and aerial parts of two cultivars of Allium sativum and their antibiofilm activity against food and nosocomial pathogens. Antibiotics 2022, 11, 724. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylases α and β. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1955; Volume 1, pp. 149–158. [Google Scholar] [CrossRef]
- Si, M.M.; Lou, J.S.; Zhou, C.X.; Shen, J.N.; Wu, H.H.; Yang, B.; He, Q.J.; Wu, H.S. Insulin releasing and alpha-glucosidase inhibitory activity of ethyl acetate fraction of Acorus calamus in vitro and in vivo. J. Ethnopharmacol. 2010, 128, 154–159. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| A | B | C | D | E | F | KI a | KI b | Identification c | |
|---|---|---|---|---|---|---|---|---|---|
| 3-Ethyl-2-pentanone | - | - | - | 0.2 | - | - | 783 | - | 1, 2 |
| 3-Methylbutyl acetate | - | - | - | 0.2 | - | - | 815 | 1132 | 1, 2 |
| 4-Methyl-2-pentyl acetate | - | - | - | - | - | 0.2 | 831 | - | 1, 2 |
| 2-Methylbutanoic anhydride | - | - | - | 31.0 | - | - | 833 | - | 1, 2 |
| α-Pinene | 16.6 | 12.0 | 21.4 | 0.7 | 1.3 | 7.5 | 857 | 1012 | 1, 2, 3 |
| 2,4-Dimethyl-3-ethylpentane | - | - | - | 0.3 | - | - | 859 | - | 1, 2 |
| Camphene | 1.0 | 0.2 | 0.9 | 0.1 | - | 0.3 | 868 | 1075 | 1, 2, 3 |
| β-Pinene | 0.2 | 0.3 | - | - | 0.3 | 0.7 | 891 | 1110 | 1, 2, 3 |
| Myrcene | - | 0.2 | - | - | 0.6 | 0.1 | 893 | 1166 | 1, 2 |
| α-Phellandrene | 0.1 | 1.0 | - | - | 1.5 | 0.1 | 918 | 1177 | 1, 2, 3 |
| 2-Ethyl-1-pentanol | - | - | - | 0.1 | - | - | 921 | - | 1, 2 |
| α-Terpinene | - | 0.1 | 0.2 | - | 0.4 | - | 930 | 1170 | 1, 2, 3 |
| p-Cymene | - | - | - | - | 1.8 | - | 938 | 1250 | 1, 2 |
| Eucalyptol | 57.0 | 50.0 | 70.5 | 32.2 | 26.3 | 53.6 | 943 | 1210 | 1, 2, 3 |
| cis-β-Ocimene | 0.3 | 0.1 | - | - | - | 0.1 | 955 | 1225 | 1, 2 |
| γ-Terpinene | 1.1 | 0.1 | - | - | 1.0 | 0.3 | 971 | 1221 | 1, 2, 3 |
| trans-p-Mentha-2,8-dienol | 0.1 | - | - | - | - | - | 981 | - | |
| Fenchone | 0.2 | - | - | - | - | - | 994 | - | |
| Terpinolene | - | 0.2 | - | - | 0.5 | - | 996 | 1267 | 1, 2, 3 |
| 6-Camphenol | 0.3 | - | - | 0.1 | - | 0.2 | 998 | - | 1, 2 |
| p-Cymenene | - | 0.1 | - | - | - | - | 999 | 1269 | 1, 2 |
| 3-Methylbutyl 2-methylbutanoate | - | - | - | 0.1 | - | - | 1008 | - | 1, 2 |
| Linalool | - | - | - | - | 0.4 | - | 1009 | 1506 | 1, 2, 3 |
| α-Pinene oxide | 0.1 | - | - | - | 0.1 | 0.1 | 1012 | 1384 | 1, 2 |
| Butanoic acid, 3-methyl-, 3-methylbutyl ester | 0.2 | - | - | 0.5 | 0.2 | 0.7 | 1015 | 1285 | 1, 2 |
| endo-Fenchol | - | - | - | - | 0.1 | 0.6 | 1016 | - | 1, 2, 3 |
| exo-Fenchol | 1.2 | 0.3 | 1.3 | 0.2 | - | - | 1019 | - | 1, 2, 3 |
| cis-p-Menth-2-en-1-ol | - | - | - | - | 0.3 | - | 1024 | - | 1, 2 |
| α-Campholenal | 0.2 | 0.1 | - | - | - | 0.1 | 1027 | - | 1, 2 |
| 2,4,4-Trimethylcyclopentanol | - | - | - | 0.1 | - | - | 1029 | - | 1, 2 |
| allo-Ocimene | 0.3 | 0.1 | - | - | - | 0.1 | 1036 | 1388 | 1, 2 |
| trans-Pinocarveol | 5.8 | 2.1 | - | 4.6 | 0.1 | 4.6 | 1040 | 1664 | 1, 2 |
| trans-p-Menth-2-en-1-ol | - | - | - | - | 0.3 | 0.1 | 1042 | 1571 | 1, 2 |
| Camphor | 0.2 | 1043 | 1491 | 1, 2, 3 | |||||
| Camphene hydrate | - | 0.2 | - | - | - | - | 1046 | - | 1, 2 |
| trans-Pinocamphone | 0.2 | 1049 | - | 1, 2 | |||||
| Pinocarvone | 1.7 | 0.6 | - | 0.9 | - | - | 1065 | - | 1, 2 |
| Borneol | 1.9 | 0.3 | 0.9 | 0.3 | 0.3 | - | 1067 | 1715 | 1, 2, 3 |
| neo-iso-Isopulegol | 0.1 | 0.2 | - | - | - | - | 1068 | - | 1, 2 |
| Pinocampheol | 0.3 | 1069 | - | 1, 2 | |||||
| Terpinen-4-ol | 0.6 | 0.5 | - | - | 2.1 | - | 1078 | 1636 | 1, 2, 3 |
| Cryptone | - | - | - | - | 0.4 | - | 1084 | 1659 | 1, 2 |
| trans-p-Mentha-1(7),8-dien-2-ol | - | 0.4 | - | 0.2 | - | - | 1088 | 1810 | 1, 2 |
| α-Terpineol | 4.1 | 1.6 | 0.1 | - | 1.6 | 0.6 | 1090 | 1662 | 1, 2, 3 |
| cis-Piperitenol | - | - | - | - | 0.2 | - | 1091 | - | 1, 2 |
| Dihydrocarveol | - | - | - | - | - | 0.4 | 1092 | - | 1, 2, 3 |
| Myrtenol | 0.3 | - | - | 0.2 | - | 0.7 | 1093 | - | 1, 2 |
| Safranal | - | - | - | - | 0.1 | - | 1095 | 1648 | 1, 2 |
| Verbenone | - | - | - | - | 0.2 | - | 1097 | - | 1, 2 |
| trans-Piperitenol | - | - | - | - | 0.3 | - | 1102 | - | 1, 2 |
| Pulegone | - | - | - | - | 0.2 | - | 1111 | 1662 | 1, 2 |
| endo-Fenchyl acetate | 0.1 | - | - | - | - | - | 1112 | - | 1, 2 |
| trans-Chrysantenyl acetate | - | - | - | - | 0.2 | - | 1114 | - | 1, 2 |
| Isobornyl acetate | 0.1 | - | - | - | - | - | 1118 | 1582 | 1, 2 |
| p-Menth-8-en-2-ol | 0.6 | - | - | - | - | - | 1226 | - | 1, 2 |
| Carveol | - | - | - | 0.1 | - | 0.1 | 1131 | - | 1, 2, 3 |
| Cumin aldehyde | - | - | - | - | 0.2 | - | 1138 | 1802 | 1, 2 |
| cis-Ocimenone | - | - | - | - | - | 0.6 | 1139 | 1225 | 1, 2 |
| Piperitone | - | - | - | - | 0.1 | - | 1144 | 1748 | 1, 2 |
| trans-Piperitenone oxide | - | - | - | 0.4 | - | - | 1148 | - | 1, 2 |
| Phellandral | - | - | - | - | 0.3 | - | 1164 | 1720 | 1, 2 |
| Bornyl acetate | 0.2 | 1176 | 1575 | 1, 2 | |||||
| Carvacrol | - | - | - | - | 0.7 | - | 1191 | 2219 | 1, 2, 3 |
| p-Cymen-7-ol | - | - | - | - | - | 0.5 | 1197 | 2113 | 1, 2 |
| δ-Elemene | - | 0.3 | - | - | 0.2 | 0.2 | 1208 | 1479 | 1, 2 |
| Piperitenone | - | 0.1 | - | - | 0.1 | - | 1215 | - | 1, 2 |
| Elixene | - | - | - | - | 0.9 | - | 1218 | - | 1, 2 |
| exo-2-Hydroxycineole acetate | 0.3 | - | - | - | 0.1 | - | 1226 | - | 1, 2 |
| α-Terpinyl acetate | - | 2.4 | - | - | - | - | 1232 | 1685 | 1, 2 |
| Isoledene | - | - | - | - | 0.1 | - | 1251 | - | 1, 2 |
| α-Copaene | - | 0.3 | - | - | - | - | 1253 | 1477 | 1, 2 |
| β-Elemene | - | - | - | - | 0.3 | - | 1275 | - | 1, 2 |
| Longifolene | - | - | - | 0.1 | 0.3 | - | 1283 | 1575 | 1, 2 |
| trans-Caryophyllene | - | 0.1 | - | - | 0.1 | 0.2 | 1284 | 1617 | 1, 2 |
| α-Gurjunene | - | 0.2 | - | - | - | 0.1 | 1285 | 1535 | 1, 2 |
| β-Gurjunene | - | - | - | - | - | 0.2 | 1298 | 1655 | 1, 2 |
| β-Cedrene | - | - | - | - | 0.9 | - | 1299 | 1613 | 1, 2 |
| β-Copaene | - | - | - | 0.1 | 0.1 | - | 1301 | - | 1, 2 |
| α-Panasinsene | 0.1 | - | - | - | - | - | 1307 | - | 1, 2 |
| α-Guaiene | - | - | - | - | 1.5 | - | 1307 | 1600 | 1, 2 |
| Aromadendrene | - | 2.9 | - | 4.4 | - | 4.7 | 1308 | 1631 | 1, 2 |
| allo-Aromadendrene | - | 0.9 | - | 0.5 | - | 1.1 | 1312 | 1660 | 1, 2 |
| cis-Cadina-1(6).4-diene | - | - | - | - | 0.1 | - | 1322 | - | 1, 2 |
| Dauca-5,8-diene | - | - | - | - | 0.1 | - | 1325 | - | 1, 2 |
| trans-Cadina-1(6),4-diene | - | - | - | - | 0.2 | - | 1332 | - | 1, 2 |
| γ-Gurjunene | - | 0.6 | - | 0.1 | - | 0.6 | 1335 | - | 1, 2 |
| β-Selinene | - | - | - | 0.2 | - | - | 1343 | 1725 | 1, 2 |
| Guaia-1(10),11-diene | - | 0.1 | - | - | - | - | 1344 | - | 1, 2 |
| Viridiflorene | - | 0.8 | - | 0.2 | - | - | 1350 | - | 1, 2 |
| α-Selinene | - | - | - | 0.1 | - | - | 1351 | 1713 | 1, 2 |
| 9-epi-β-Caryophyllene | - | - | - | - | - | 0.4 | 1352 | - | 1, 2 |
| Eudesma-4(14),11-diene | - | - | - | - | 0.1 | - | 1353 | 1708 | 1, 2 |
| 6-[1-(Hydroxymethyl)vinyl]-4,8a-dimethyl-1,2,3,5,6,7,8,8a-octahydro-2-naphthalenol | - | - | - | - | 0.7 | - | 1356 | - | 1, 2 |
| Longifolene | 0.1 | - | - | - | - | - | 1364 | 1574 | 1, 2 |
| dehydro-Aromadendrene | - | 0.4 | - | - | 0.4 | - | 1366 | - | 1, 2 |
| γ-Muurolene | - | 0.1 | - | - | - | 0.1 | 1376 | 1725 | 1, 2 |
| trans-Muurola-4(14),5-diene | - | - | - | - | - | 0.4 | 1381 | - | 1, 2 |
| trans-Cycloisolongifol-5-ol | - | - | - | - | 0.1 | - | 1387 | - | 1, 2 |
| Cadina-3,9-diene | - | 0.2 | - | - | - | - | 1392 | - | 1, 2 |
| γ-Vetivenene | - | - | - | - | 0.2 | - | 1411 | - | 1, 2 |
| α-Amorphene | - | - | - | - | - | 1.3 | 1419 | 1750 | 1, 2 |
| Macrocarpal | - | - | - | - | 0.5 | - | 1420 | - | 1, 2 |
| 10-epi-Cubebol | - | - | - | 3.4 | - | 4.6 | 1421 | - | 1, 2 |
| trans-β-Guaiene | - | 0.5 | - | - | - | - | 1425 | 1651 | 1, 2 |
| δ-Selinene | - | - | - | - | - | 0.6 | 1426 | - | 1, 2 |
| Isoaromadendrene epoxide | - | - | - | - | 0.3 | - | 1431 | - | 1, 2 |
| γ-Patchoulene | - | - | - | - | - | 1.3 | 1436 | - | 1, 2 |
| Germacrene B | - | - | - | 10.2 | - | - | 1437 | 1795 | 1, 2 |
| Spathulenol | 0.1 | 2.3 | - | - | 28.3 | - | 1438 | 2127 | 1, 2 |
| Globulol | - | 3.9 | - | 1.2 | - | 0.1 | 1442 | 2104 | 1, 2 |
| Epiglobulol | 0.1 | - | - | - | 0.6 | - | 1444 | - | 1, 2 |
| α-Cadinene | - | - | - | - | - | 1.3 | 1450 | - | 1, 2 |
| Cubeban-11-ol | - | 0.3 | - | 0.6 | - | - | 1453 | - | 1, 2 |
| Guaiol | 0.1 | - | - | - | - | - | 1454 | 2094 | 1, 2 |
| Rosifoliol | - | 0.5 | - | 1.0 | - | 0.5 | 1461 | - | 1, 2 |
| cis-Cadin-4-en-7-ol | - | - | - | - | - | 0.3 | 1473 | - | 1, 2 |
| Khusimone | - | 0.6 | - | - | - | - | 1474 | - | 1, 2 |
| 9,11-epoxy-Guaia-3,10(14)-diene | - | - | - | 0.6 | - | - | 1475 | - | 1, 2 |
| γ-Eudesmol | - | 2.9 | - | 0.1 | - | - | 1477 | 2178 | 1, 2 |
| 1,7-diepi-α-Cedrenal | - | - | - | 0.1 | - | - | 1493 | - | 1, 2 |
| Cubenol | - | - | - | - | 0.3 | 0.4 | 1484 | 2080 | 1, 2 |
| β-Eudesmol | - | 3.2 | - | 0.8 | 3.9 | 1.6 | 1485 | 2248 | 1, 2 |
| Cedr-8(15)-en-9-α-ol | - | - | - | - | 2.0 | - | 1487 | - | 1, 2 |
| α-Eudesmol | - | 2.8 | - | 0.6 | - | 0.4 | 1489 | 2247 | 1, 2 |
| α-Cadinol | - | - | - | - | 0.7 | - | 1490 | 2224 | 1, 2 |
| Selin-11-en-4-α-ol | - | - | - | - | 0.3 | - | 1492 | 2273 | 1, 2 |
| Murolan-3,9(11)-diene-10-peroxy | - | - | - | 0.2 | - | - | 1497 | - | 1, 2 |
| 7-epi-α-Eudesmol | - | - | - | - | 1.8 | - | 1506 | - | 1, 2 |
| Germacra-4(15),5,10(14)-trien-1-α-ol | - | - | - | - | 0.8 | - | 1521 | - | 1, 2 |
| Corymbolone | - | - | - | 0.1 | - | - | 1567 | - | 1, 2 |
| Eudesma-4(15),7-dien-1β-ol | - | - | - | - | 0.2 | - | 1571 | - | 1, 2 |
| Cyperotundone | - | - | - | - | 3.0 | - | 1584 | - | 1, 2 |
| Eudesm-7(11)-en-4-ol | - | - | - | - | 1.9 | - | 1586 | - | 1, 2 |
| 1,1,4,6-Tetramethyldecahydro-1H-cyclopropa[e]azulene-4,5,6-triol | - | - | - | 0.2 | - | - | 1587 | - | 1, 2 |
| epi-Cyclolorenone | - | - | - | - | 0.8 | - | 1602 | - | 1, 2 |
| Stigmasterol acetate | 0.1 | - | - | - | - | - | 2735 | - | 1, 2 |
| Stigmastan-3,5,22-trien | 0.1 | - | - | - | - | - | 2748 | - | 1, 2 |
| Stigmast-5-en-3-ol, oleate | 0.6 | - | - | 0.2 | 0.1 | - | 2769 | - | 1, 2 |
| Total | 97.2 | 97.4 | 95.3 | 97.5 | 94.1 | 97 | |||
| Monoterpene hydrocarbons | 19.6 | 14.1 | 22.5 | 0.8 | 7.4 | 9 | |||
| Oxygenated monoterpenes | 75.7 | 58.8 | 72.8 | 39.2 | 34.7 | 67 | |||
| Sesquiterpene hydrocarbons | 0.2 | 8.0 | - | 15.9 | 5.5 | 12.2 | |||
| Oxygenated sesquiterpenes | 0.7 | 16.5 | - | 8.9 | 46.2 | 7.9 | |||
| Others | 1.0 | - | - | 32.7 | 0.3 | 0.9 |
| Doses (mg/mL) | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus | |
|---|---|---|---|---|---|---|
| E. brevifolia | 10 | 0.00 ± 0.00 | 70.91 ± 3.47 d | 73.53 ± 5.07 d | 39.71 ± 2.78 d | 0.00± 0.00 |
| 20 | 0.00 ± 0.00 | 77.30 ± 1.67 d | 87.38 ± 1.23 d | 44.25 ± 2.67 d | 35.69 ± 3.08 d | |
| E. extensa | 5 | 0.00 ± 0.00 | 30.89 ± 2.67 d | 55.10 ± 1.25 d | 57.11 ± 4.45 d | 9.94 ± 0.78 d |
| 10 | 0.00 ± 0.00 | 89.76 ± 2.06 d | 83.27± 1.36 d | 63.61 ± 1.67 d | 73.73 ± 3.37 d | |
| E. lehmannii | 10 | 0.00 ± 0.00 | 21.52 ± 1.44 d | 14.98 ± 0.76 d | 75.38 ± 3.98 d | 16.06 ± 1.02 d |
| 20 | 34.28 ± 2.24 d | 39.08 ± 2.44 d | 32.27 ± 2.01 d | 78.13 ± 3.35 d | 61.15 ± 3.65 d | |
| E. leptophylla | 10 | 84.96 ± 1.02 d | 54.92 ± 3.52 d | 15.40 ± 2.01 d | 83.12 ± 2.09 d | 58.05 ± 1.76 d |
| 20 | 88.91 ± 1.32 d | 60.86 ± 5.05 d | 60.47 ± 4.54 d | 93.17 ± 1.02 d | 79.12 ± 2.67 d | |
| E. patellaris | 10 | 32.37 ± 2.45 d | 56.04 ± 4.12 d | 77.34 ± 2.76 d | 74.41 ± 2.87 d | 72.48 ± 0.01 d |
| 20 | 37.63 ± 1.98 d | 59.83 ± 2.86 d | 88.56 ± 1.08 d | 84.70 ± 2.05 d | 89.39 ± 3.02 d | |
| E. woollsiana | 10 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 12.91 ± 1.67 d | 45.89 ± 4.01 d |
| 20 | 37.59 ± 2.47 d | 6.05 ± 0.33 d | 0.00 ± 0.00 | 15.60 ± 1.54 d | 67.27 ± 5.97 d |
| Doses (mg/mL) | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus | |
|---|---|---|---|---|---|---|
| E. brevifolia | 10 | 18.99 ± 2.76 d | 27.46 ± 1.66 d | 25.53 ± 2.07 d | 20.27 ± 1.89 d | 12.21 ± 0.98 d |
| 20 | 28.83 ± 1.87 d | 78.8 ± 4.07 d | 32.46 ± 3.33 d | 30.1 ± 2.76 d | 76.26 ± 2.65 d | |
| E. extensa | 5 | 49.95 ± 4.44 d | 17.72 ± 1.01 d | 83.86 ± 4.91 d | 0.00 ± 0.00 | 0.00 ± 0.00 |
| 10 | 65.40 ± 4.57 d | 56.63 ± 0.97 d | 89.26 ± 3.54 d | 45.37 ± 1.33 d | 11.30 ± 1.07 d | |
| E. lehmannii | 10 | 51.39 ± 4.55 d | 33.34 ± 2.91 d | 63.36 ± 3.91 d | 84.05 ± 1.67 d | 59.57 ± 3.67 d |
| 20 | 54.35 ± 3.85 d | 21.74 ± 1.76 d | 64.73 ± 3.91 d | 67.77 ± 3.92 d | 68.66 ± 2.64 d | |
| E. leptophylla | 10 | 54.92 ± 3.14 d | 15.41 ± 1.05 d | 83.12 ± 1.18 d | 58.05 ± 1.66 d | 51.23 ± 3.01 d |
| 20 | 60.86 ± 3.94 d | 60.47 ± 3.08 d | 93.17 ± 1.13 d | 79.12 ± 2.97 d | 81.04 ± 2.76 d | |
| E. patellaris | 10 | 26.47 ± 2.01 d | 19.48 ± 0.78 d | 0.00 ± 0.00 | 16.44 ± 1.02 d | 29.89 ± 1.79 d |
| 20 | 58.75 ± 3.02 d | 36.12 ± 2.41 d | 22.11 ± 2.55 d | 37.25 ± 3.07 d | 32.21 ± 1.98 d | |
| E. woolsiana | 10 | 75.08 ± 4.22 d | 32.55 ± 2.26 d | 0.00 ± 0.00 | 0.00 ± 0.00 | 48.04 ± 5.01 d |
| 20 | 93.27 ± 1.01 d | 39.58 ± 4.12 d | 7.02 ± 1.22 d | 10.76 ± 1.01 d | 55.59 ± 3.72 d |
| EOs | IC50 (mg/mL) | |||
|---|---|---|---|---|
| AChE | BChE | α-Amylase | α-Glucosidase | |
| E. brevifolia | 0.3 ± 0.04 a | 11.86 ± 3.54 c | 0.88 ± 0.14 a | 27.31 ± 2.11 a |
| E. extensa | 0.25 ± 0.01 a | 7.37 ± 0.06 b | n.a | n.a |
| E. lehmanii | 1.2 ± 0.40 b | 3.48± 0.37 a | 16.94 ± 2.14 b | n.a |
| E. leptophylla | 0.57 ± 0.03 a | 4.54 ± 0.11 ab | 0.88 ± 0.05 a | 29.0 ± 1.32 a |
| E. patellaris | 0.60 ± 0.04 a | 3.50 ± 0.10 a | 0.91 ± 0.01 a | n.a |
| E. woollsiana | 0.38 ± 0.01 a | 4.49 ± 0.01 ab | 11.9 ± 1.3 b | n.a |
| Galantamine | 0.008 ± 0.003 | 0.05 ± 0.01 | - | - |
| Acarbose | - | - | 0.004 ± 0.002 | 0.6 ± 0.3 |
| Species | Arboretum (Governorate) | Harvest Period | Bioclimatic Condition | Yields % |
|---|---|---|---|---|
| E. brevifolia | Hajeb Layoun (Kairouan) | April 2023 | Semi-arid upper with moderate winters | 2.42 |
| E. extensa | Souiniet (Ain Draham) | June 2021 | Upper humid | 1.43 |
| E. lehmannii | Korbous (Nabeul) | May 2022 | Sub-humid | 1.91 |
| E. leptophylla | Hajeb Layoun (Kairouan) | April 2023 | Semi-arid upper with moderate winters | 2.03 |
| E. patellaris | Hajeb Layoun (Kairouan) | April 2023 | Semi-arid upper with moderate winters | 0.25 |
| E. woolsiana | Djebel Mansour (Zaghouan) | Mars 2021 | Semi-arid | 1.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khedhri, S.; Polito, F.; Caputo, L.; De Feo, V.; Khamassi, M.; Kochti, O.; Hamrouni, L.; Mabrouk, Y.; Nazzaro, F.; Fratianni, F.; et al. Chemical Composition, Antibacterial Properties, and Anti-Enzymatic Effects of Eucalyptus Essential Oils Sourced from Tunisia. Molecules 2023, 28, 7211. https://doi.org/10.3390/molecules28207211
Khedhri S, Polito F, Caputo L, De Feo V, Khamassi M, Kochti O, Hamrouni L, Mabrouk Y, Nazzaro F, Fratianni F, et al. Chemical Composition, Antibacterial Properties, and Anti-Enzymatic Effects of Eucalyptus Essential Oils Sourced from Tunisia. Molecules. 2023; 28(20):7211. https://doi.org/10.3390/molecules28207211
Chicago/Turabian StyleKhedhri, Sana, Flavio Polito, Lucia Caputo, Vincenzo De Feo, Marwa Khamassi, Oumayama Kochti, Lamia Hamrouni, Yassine Mabrouk, Filomena Nazzaro, Florinda Fratianni, and et al. 2023. "Chemical Composition, Antibacterial Properties, and Anti-Enzymatic Effects of Eucalyptus Essential Oils Sourced from Tunisia" Molecules 28, no. 20: 7211. https://doi.org/10.3390/molecules28207211
APA StyleKhedhri, S., Polito, F., Caputo, L., De Feo, V., Khamassi, M., Kochti, O., Hamrouni, L., Mabrouk, Y., Nazzaro, F., Fratianni, F., & Amri, I. (2023). Chemical Composition, Antibacterial Properties, and Anti-Enzymatic Effects of Eucalyptus Essential Oils Sourced from Tunisia. Molecules, 28(20), 7211. https://doi.org/10.3390/molecules28207211