
(−)-Epigallocatechin-3-Gallate Reduces Perfluorodecanoic Acid-Exacerbated Adiposity and Hepatic Lipid Accumulation in High-Fat Diet-Fed Male C57BL/6J Mice


Abstract
:1. Introduction
2. Results
2.1. EGCG Reduced Body Weight Gain in PFDA-Exposed Mice Fed an HFD
2.2. EGCG Decreased Tissue and Organ Weights in PFDA-Exposed Mice Fed an HFD
2.3. EGCG Reduced Blood Glucose and Blood Lipid Parameters in PFDA-Exposed Mice Fed an HFD
2.4. EGCG Improved Liver Function in PFDA-Exposed Mice Fed an HFD
2.5. EGCG Inhibited Serum Inflammatory Cytokines
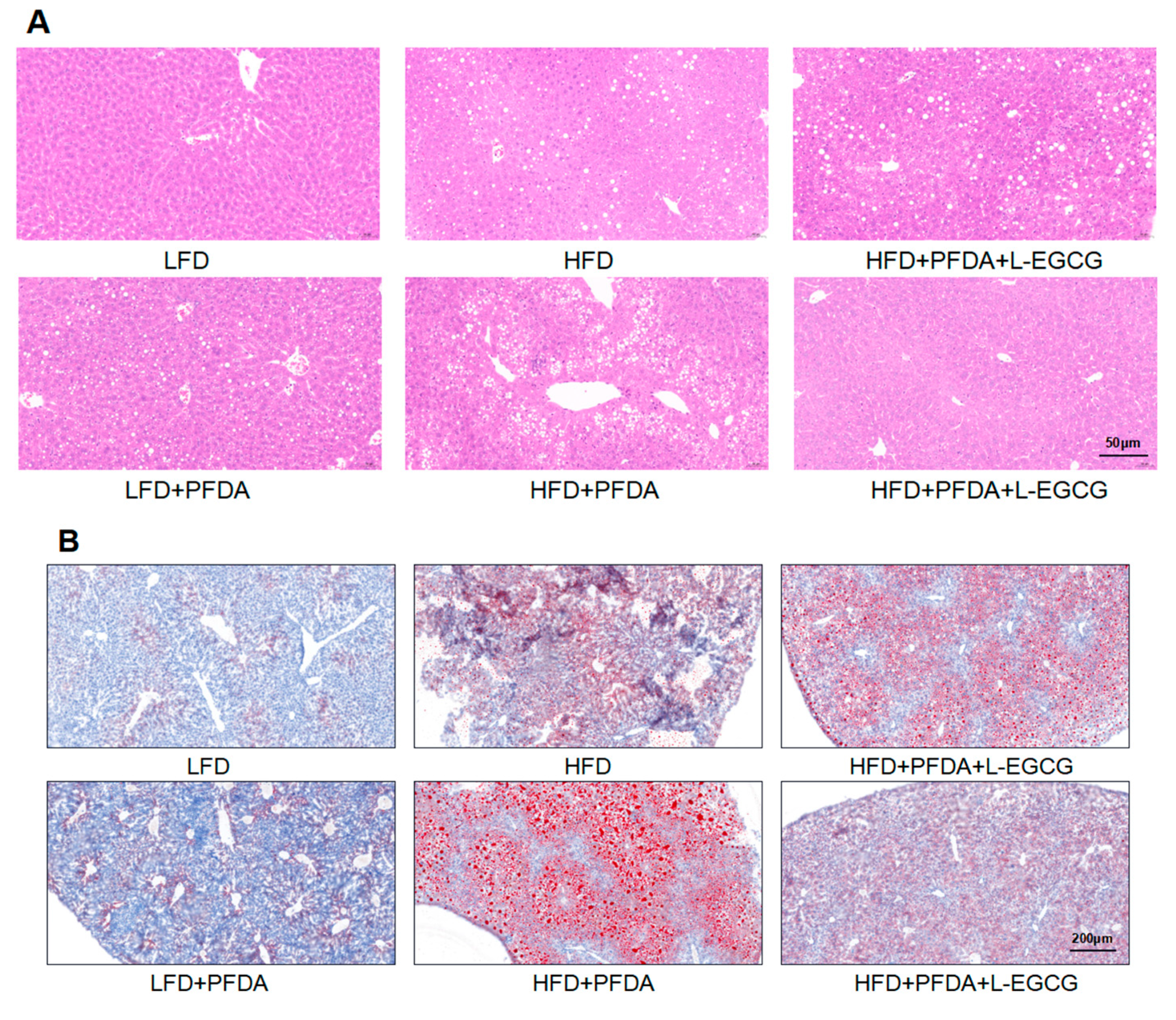
2.6. EGCG Reduced Lipid Accumulation in Mice Co-Exposed to an HFD and PFDA
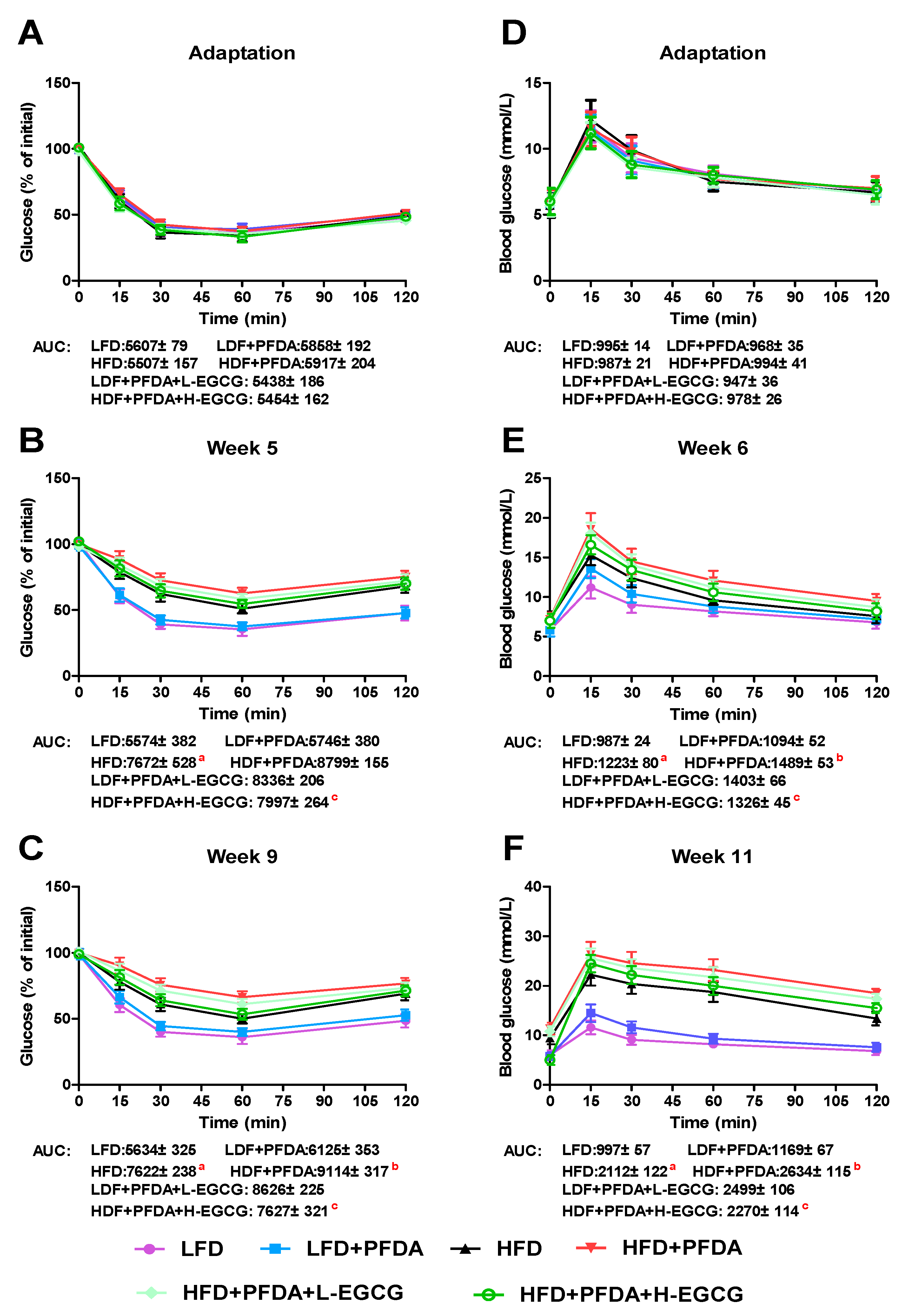
2.7. Effect of EGCG on Glucose Homeostasis
2.8. EGCG Regulated Hepatic Lipid Metabolism in PFDA-Exposed Mice Fed an HFD
2.9. EGCG Inhibited Hepatic NLRP3 Inflammasome Expression in PFDA-Exposed Mice Fed an HFD
3. Discussion
4. Materials and Methods
4.1. Animals, Diet, and Treatment
4.2. Quantification of Serum Parameters
4.3. Liver Function Analysis
4.4. Serum Inflammatory Cytokine Quantification
4.5. Metabolic Testing
4.6. The Quantification of Lipid Accumulation within the Hepatic Tissue
4.7. mRNA Expression Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eschauzier, C.; Hoppe, M.; Schlummer, M.; de Voogt, P. Presence and sources of anthropogenic perfluoroalkyl acids in high-consumption tap-water based beverages. Chemosphere 2013, 90, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zhao, Q.; Fan, Z.; Jia, S.; Liu, Q.; Liu, F.; Liu, S. The toxicity of perfluorodecanoic acid is mainly manifested as a deflected immune function. Mol. Biol. Rep. 2022, 49, 4365–4376. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.; Anitole, K.; Hodes, C.; Lai, D.; Pfahles-Hutchens, A.; Seed, J. Perfluoroalkyl acids: A review of monitoring and toxicological findings. Toxicol. Sci. 2007, 99, 366–394. [Google Scholar] [CrossRef] [PubMed]
- Calafat, A.M.; Wong, L.Y.; Kuklenyik, Z.; Reidy, J.A.; Needham, L.L. Polyfluoroalkyl chemicals in the U.S. population: Data from the National Health and Nutrition Examination Survey (NHANES) 2003–2004 and comparisons with NHANES 1999–2000. Environ. Health Perspect. 2007, 115, 1596–1602. [Google Scholar] [CrossRef] [PubMed]
- Yeung, L.W.; So, M.K.; Jiang, G.; Taniyasu, S.; Yamashita, N.; Song, M.; Wu, Y.; Li, J.; Giesy, J.P.; Guruge, K.S.; et al. Perfluorooctanesulfonate and related fluorochemicals in human blood samples from China. Environ. Sci. Technol. 2006, 40, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.Z.; Xu, C.L.; Xu, Z.F.; Zhou, L.Y.; Xie, S.J.; Wei, K.N.; Jin, Y.C.; Zeng, Z.C.; Yang, X.J.; Tan, S.H.; et al. Perfluorodecanoic acid induces meiotic defects and deterioration of mice oocytes in vitro. Toxicology 2021, 460, 152884. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Tan, Z.; Dai, M.; Song, D.; Lin, J.; Xie, M.; Yang, J.; Sun, L.; Wei, D.; Zhao, J.; et al. Dual action of peroxisome proliferator-activated receptor alpha in perfluorodecanoic acid-induced hepatotoxicity. Arch. Toxicol. 2017, 91, 897–907. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, T.; Lv, C.; Niu, Q.; Zong, W.; Tang, J.; Liu, R. Perfluorodecanoic acid-induced oxidative stress and DNA damage investigated at the cellular and molecular levels. Ecotoxicol. Environ. Saf. 2019, 185, 109699. [Google Scholar] [CrossRef]
- Li, Y.; Yan, H.; Liu, Q.; Li, X.; Ge, J.; Yu, X. Accumulation and transport patterns of six phthalic acid esters (PAEs) in two leafy vegetables under hydroponic conditions. Chemosphere 2020, 249, 126457. [Google Scholar] [CrossRef]
- Valvi, D.; Højlund, K.; Coull, B.A.; Nielsen, F.; Weihe, P.; Grandjean, P. Life-course exposure to perfluoroalkyl substances in relation to markers of glucose homeostasis in early adulthood. J. Clin. Endocrinol. Metab. 2021, 106, 2495–2504. [Google Scholar] [CrossRef]
- Yu, G.; Jin, M.; Huang, Y.; Aimuzi, R.; Zheng, T.; Nian, M.; Tian, Y.; Wang, W.; Luo, Z.; Shen, L.; et al. Environmental exposure to perfluoroalkyl substances in early pregnancy, maternal glucose homeostasis and the risk of gestational diabetes: A prospective cohort study. Environ. Int. 2021, 156, 106621. [Google Scholar] [CrossRef] [PubMed]
- Bloom, M.S.; Commodore, S.; Ferguson, P.L.; Neelon, B.; Pearce, J.L.; Baumer, A.; Newman, R.B.; Grobman, W.; Tita, A.; Roberts, J.; et al. Association between gestational PFAS exposure and Children’s adiposity in a diverse population. Environ. Res. 2022, 203, 111820. [Google Scholar] [CrossRef]
- Tan, X.; Xie, G.; Sun, X.; Li, Q.; Zhong, W.; Qiao, P.; Sun, X.; Jia, W.; Zhou, Z. High fat diet feeding exaggerates perfluorooctanoic acid-induced liverinjury in mice via modulating multiple metabolic pathways. PLoS ONE 2013, 8, e61409. [Google Scholar] [CrossRef] [PubMed]
- Lian, C.-Y.; Zhai, Z.-Z.; Li, Z.-F.; Wang, L. High fat diet-triggered non-alcoholic fatty liver disease: A review of proposed mechanisms. Chem. Interact. 2020, 330, 109199. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Maternal high-fat diet and offspring hypertension. Int. J. Mol. Sci. 2022, 23, 8179. [Google Scholar] [CrossRef] [PubMed]
- Prasad, M.; Rajagopal, P.; Devarajan, N.; Veeraraghavan, V.P.; Palanisamy, C.P.; Cui, B.; Patil, S.; Jayaraman, S. A comprehensive review on high -fat diet-induced diabetes mellitus: An epigenetic view. J. Nutr. Biochem. 2022, 107, 109037. [Google Scholar] [CrossRef] [PubMed]
- Kahn, B.B.; Flier, J.S. Obesity and insulin resistance. J. Clin. Investig. 2000, 106, 473–481. [Google Scholar] [CrossRef]
- Grundy, S.M. Obesity, metabolic syndrome, and cardiovascular disease. J. Clin. Endocrinol. Metab. 2004, 89, 2595–2600. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, W.; Zhou, R. NLRP3 inflammasome activation and cell death. Cell. Mol. Immunol. 2021, 18, 2114–2127. [Google Scholar] [CrossRef]
- Sharma, B.R.; Kanneganti, T.-D. NLRP3 inflammasome in cancer and metabolic diseases. Nat. Immunol. 2021, 22, 550–559. [Google Scholar] [CrossRef]
- Shao, W.; Yeretssian, G.; Doiron, K.; Hussain, S.N.; Saleh, M. The caspase-1 digestome identifies the glycolysis pathway as a target during infection and septic shock. J. Biol. Chem. 2007, 282, 36321–36329. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, C.; Yang, J.; Chen, Y.; Zhou, B.; Abbott, D.W.; Xiao, T.S. Caspase-1 engages full-length gasdermin D through two distinct interfaces that mediate caspase recruitment and substrate cleavage. Immunity 2020, 53, 106–114.e5. [Google Scholar] [CrossRef] [PubMed]
- Mitsuyoshi, H.; Yasui, K.; Hara, T.; Taketani, H.; Ishiba, H.; Okajima, A.; Seko, Y.; Umemura, A.; Nishikawa, T.; Yamaguchi, K.; et al. Hepatic nucleotide binding oligomerization domain-like receptors pyrin domain-containing 3 inflammasomes are associated with the histologic severity of non-alcoholic fatty liver disease. Hepatol. Res. 2017, 47, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Varghese, J.F.; Patel, R.; Yadav, U.C. Sterol regulatory element binding protein (SREBP) -1 mediates oxidized low-density lipoprotein (oxLDL) induced macrophage foam cell formation through NLRP3 inflammasome activation. Cell. Signal. 2018, 53, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.G.; Fang, W.L.; Wei, J.; Wang, T.; Wang, N.; Ma, J.L.; Shi, M. The involvement of NLRX1 and NLRP3 in the development of nonalcoholic steatohepatitis in mice. J. Chin. Med. Assoc. 2013, 76, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, H.; Guo, Y.; Guo, Y.; Guan, H.; Wang, D. Perfluorodecanoic acid promotes high-fat diet-triggered adiposity and hepatic lipid accumulation by modulating the NLRP3/caspase-1 pathway in male C57BL/6J mice. Food Chem. Toxicol. 2023, 78, 113943. [Google Scholar] [CrossRef]
- Wang, T.; Xu, H.; Guo, Y.; Li, Z.; Ye, H.; Wu, L.; Guo, Y.; Wang, D. Perfluorodecanoic acid promotes adipogenesis via NLRP3 inflammasome-mediated pathway in HepG2 and 3T3-L1 cells. Food Chem. Toxicol. 2023, 171, 113520. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, Q.; Liu, L.; Hu, Y.Y.; Feng, Q. Potential biological effects of (-)-epigallocatechin-3-gallate on the treatment of nonalcoholic fatty liver disease. Mol. Nutr. Food Res. 2018, 62, 1700483. [Google Scholar] [CrossRef]
- Naito, Y.; Ushiroda, C.; Mizushima, K.; Inoue, R.; Yasukawa, Z.; Abe, A.; Takagi, T. Epigallocatechin-3-gallate (EGCG) attenuates non-alcoholic fatty liver disease via modulating the interaction between gut microbiota and bile acids. J. Clin. Biochem. Nutr. 2020, 67, 2–9. [Google Scholar] [CrossRef]
- Dey, P.; Olmstead, B.D.; Sasaki, G.Y.; Vodovotz, Y.; Yu, Z.; Bruno, R.S. Epigallocatechin gallate but not catechin prevents nonalcoholic steatohepatitis in mice similar to green tea extract while differentially affecting the gut microbiota. J. Nutr. Biochem. 2020, 84, 108455. [Google Scholar] [CrossRef]
- Huang, J.; Li, W.; Liao, W.; Hao, Q.; Tang, D.; Wang, D.; Wang, Y.; Ge, G. Green tea polyphenol epigallocatechin-3-gallate alleviates nonalcoholic fatty liver disease and ameliorates intestinal immunity in mice fed a high-fat diet. Food Funct. 2020, 11, 9924–9935. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Paglicawan, L.; Soomro, S.; Abunofal, O.; Baig, S.; Vanarsa, K.; Hicks, J.; Mohan, C. Epigallocatechin-3-gallate dampens non-alcoholic fatty liver by modulating liver function, lipid profile and macrophage polarization. Nutrients 2021, 13, 599. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.; Shen, C.L. Novel insights of dietary polyphenols and obesity. J. Nutr. Biochem. 2014, 25, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bose, M.; Lambert, J.D.; Ju, J.; Reuhl, K.R.; Shapses, S.A.; Yang, C.S. The major green tea polyphenol, (-)-epigallocatechin-3-gallate, inhibits obesity, metabolic syndrome, and fatty liver disease in high-fat-fed mice. J. Nutr. 2008, 138, 1677–1683. [Google Scholar] [CrossRef]
- Jang, H.J.; Ridgeway, S.D.; Kim, J.A. Effects of the green tea polyphenol epigallocatechin-3-gallate on high-fat diet-induced insulin resistance and endothelial dysfunction. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1444–E1451. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Cao, Y.; Fan, C.; Fan, Y.; Bai, S.; Teng, W.; Shan, Z. Epigallocatechin gallate improves insulin signaling by decreasing toll-like receptor 4 (TLR4) activity in adipose tissues of high-fat diet rats. Mol. Nutr. Food Res. 2014, 58, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Li, Y.; Ling, F.; Guan, Y.; Zhang, D.; Zhu, Q.; Liu, J.; Wu, Y.; Niu, Y. The phytochemical epigallocatechin gallate prolongs the lifespan by improving lipid metabolism, reducing inflammation and oxidative stress in high-fat diet-fed obese rats. Aging Cell 2020, 19, e13199. [Google Scholar] [CrossRef]
- Eberlé, D.; Hegarty, B.; Bossard, P.; Ferré, P.; Foufelle, F. SREBP transcription factors: Master regulators of lipid homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar] [CrossRef]
- Sharma, A.; Anand, S.K.; Singh, N.; Dwivedi, U.N.; Kakkar, P. Berbamine induced AMPK activation regulates mTOR/SREBP-1c axis and Nrf2/ARE pathway to allay lipid accumulation and oxidative stress in steatotic HepG2 cells. Eur. J. Pharmacol. 2020, 882, 173244. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Y.; Wen, Y.; Zhu, S.; Huang, S.; He, L.; Hou, S.; Lai, X.; Chen, S.; Dai, Z.; et al. Phloridzin ameliorates lipid deposition in high-fat-diet-fed mice with nonalcoholic fatty liver disease via inhibiting the mTORC1/SREBP-1c pathway. J. Agric. Food Chem. 2021, 69, 8671–8683. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, C.; Song, Y.; Chen, L.; Chen, X.; Zheng, G.; Yang, Y.; Cao, P.; Qiu, Z. Gallic acid impairs fructose-driven de novo lipogenesis and ameliorates hepatic steatosis via AMPK-dependent suppression of SREBP-1/ACC/FASN cascade. Eur. J. Pharmacol. 2023, 940, 175457. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Kim, C.T.; Kim, Y. Green tea (-)-epigallocatechin-3-gallate reduces body weight with regulation of multiple genes expression in adipose tissue of diet-induced obese mice. Ann. Nutr. Metab. 2009, 54, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Bezzina, R.; Hinch, E.; Lewandowski, P.A.; Cameron-Smith, D.; Mathai, M.L.; Jois, M.; Sinclair, A.J.; Begg, D.P.; Wark, J.D.; et al. Green tea, black tea, and epigallocatechin modify body composition, improve glucose tolerance, and differentially alter metabolic gene expression in rats fed a high-fat diet. Nutr. Res. 2009, 29, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, Z.Z.; Wu, Y.; Wang, R.Q.; Chen, J.W.; Chen, J.; Zhang, Y.; Chen, Y.J.; Geng, M.; Xu, Z.D.; et al. (-)-Epigallocatechin-3-gallate ameliorates atherosclerosis and modulates hepatic lipid metabolic gene expression in apolipoprotein E knockout mice: Involvement of TTC39B. Front. Pharmacol. 2018, 9, 195. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, S.; Chen, M.; Wang, J.; Xie, B.; Sun, Z. (-)-Epigallocatechin-3-gallate (EGCG) inhibits starch digestion and improves glucose homeostasis through direct or indirect activation of PXR/CAR-mediated phase II metabolism in diabetic mice. Food Funct. 2018, 9, 4651–4663. [Google Scholar] [CrossRef] [PubMed]
- Borges, T.; Robertson, L.W.; Peterson, R.E.; Glauert, H.P. Dose-related effects of perfluorodecanoic acid on growth, feed intake and hepatic peroxisomal beta-oxidation. Arch. Toxicol. 1992, 66, 18–22. [Google Scholar] [CrossRef]
- Maher, J.M.; Aleksunes, L.M.; Dieter, M.Z.; Tanaka, Y.; Peters, J.M.; Manautou, J.E.; Klaassen, C.D. Nrf2- and PPAR alpha-mediated regulation of hepatic Mrp transporters after exposure to perfluorooctanoic acid and perfluorodecanoic acid. Toxicol. Sci. 2008, 106, 319–328. [Google Scholar] [CrossRef]
- Zhou, X.; Dong, T.; Fan, Z.; Peng, Y.; Zhou, R.; Wang, X.; Song, N.; Han, M.; Fan, B.; Jia, J.; et al. Perfluorodecanoic acid stimulates NLRP3 inflammasome assembly in gastric cells. Sci. Rep. 2017, 7, 45468. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Q.; Wang, T.; Kan, Z.; Li, X.; Hu, L.; Peng, C.Y.; Qian, F.; Wang, Y.; Granato, D. Green tea polyphenols and epigallocatechin-3-gallate protect against perfluorodecanoic acid induced liver damage and inflammation in mice by inhibiting NLRP3 inflammasome activation. Food Res. Int. 2020, 127, 108628. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.; Hu, X.; Xu, Q.; Zhang, Y.; Liu, H.; Diao, Y.; Zhang, X.; Li, L.; Yu, J.; et al. Epigallocatechin-3-gallate prevents inflammation and diabetes -Induced glucose tolerance through inhibition of NLRP3 inflammasome activation. Int. Immunopharmacol. 2021, 93, 107412. [Google Scholar] [CrossRef]
- Luo, Z.L.; Sun, H.Y.; Wu, X.B.; Cheng, L.; Ren, J.D. Epigallocatechin-3-gallate attenuates acute pancreatitis induced lung injury by targeting mitochondrial reactive oxygen species triggered NLRP3 inflammasome activation. Food Funct. 2021, 12, 5658–5667. [Google Scholar] [CrossRef]
- Di, M.; Zhang, Q.; Wang, J.; Xiao, X.; Huang, J.; Ma, Y.; Yang, H.; Li, M. Epigallocatechin-3-gallate (EGCG) attenuates inflammatory responses and oxidative stress in lipopolysaccharide (LPS)-induced endometritis via silent information regulator transcript-1 (SIRT1)/nucleotide oligomerization domain (NOD)-like receptor pyrin domain-containing 3 (NLRP3) pathway. J. Biochem. Mol. Toxicol. 2022, 36, e23203. [Google Scholar]
- Barra, N.G.; Henriksbo, B.D.; Anhê, F.F.; Schertzer, J.D. The NLRP3 inflammasome regulates adipose tissue metabolism. Biochem. J. 2020, 477, 1089–1107. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.K. Lipids, inflammasomes, metabolism, and disease. Immunol. Rev. 2020, 297, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Stienstra, R.; Joosten, L.A.; Koenen, T.; van Tits, B.; van Diepen, J.A.; van den Berg, S.A.; Rensen, P.C.; Voshol, P.J.; Fantuzzi, G.; Hijmans, A.; et al. The inflammasome-mediated caspase-1 activation controls adipocyte differentiation and insulin sensitivity. Cell. Metab. 2010, 12, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, K.; Wan, X.; Wang, F.; Guo, Z.; Mo, Z. NLRP3 inflammasome activation in mesenchymal stem cells inhibits osteogenic differentiation and enhances adipogenic differentiation. Biochem. Biophys. Res. Commun. 2017, 484, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Dhana, K.; Furtado, J.D.; Rood, J.; Zong, G.; Liang, L.; Qi, L.; Bray, G.A.; DeJonge, L.; Coull, B.; et al. Perfluoroalkyl substances and changes in body weight and resting metabolic rate in response to weight-loss diets: A prospective study. PLoS Med. 2018, 15, e1002502. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, B.; Hu, Y.; Rood, J.; Liang, L.; Qi, L.; Bray, G.A.; DeJonge, L.; Coull, B.; Grandjean, P.; et al. Associations of perfluoroalkyl substances with blood lipids and Apolipoproteins in lipoprotein subspecies: The POUNDS-lost study. Environ. Health 2020, 19, 5. [Google Scholar] [CrossRef]
- Vannice, G.; Rasmussen, H. Position of the academy of nutrition and dietetics: Dietary fatty acids for healthy adults. J. Acad. Nutr. Diet. 2014, 114, 136–153. [Google Scholar] [CrossRef]
- Chen, I.J.; Liu, C.Y.; Chiu, J.P.; Hsu, C.H. Therapeutic effect of high-dose green tea extract on weight reduction: A randomized, double-blind, placebo-controlled clinical trial. Clin. Nutr. 2016, 35, 592–599. [Google Scholar] [CrossRef]
- Huang, L.H.; Liu, C.Y.; Wang, L.Y.; Huang, C.J.; Hsu, C.H. Effects of green tea extract on overweight and obese women with high levels of low density-lipoprotein-cholesterol (LDL-C): A randomised, double-blind, and cross-over placebo-controlled clinical trial. BMC Complement. Altern. Med. 2018, 18, 294. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LFD | LFD + PFDA | HFD | HFD + PFDA | HFD + PFDA + L-EGCG | HFD + PFDA + H-EGCG | |
|---|---|---|---|---|---|---|
| Liver | 3.51 ± 0.10 | 3.46 ± 0.10 | 3.58 ± 0.16 | 4.54 ± 0.22 b | 4.24 ± 0.18 | 3.72 ± 0.20 d |
| Heart | 0.44 ± 0.05 | 0.47 ± 0.06 | 0.49 ± 0.08 | 0.44 ± 0.06 | 0.48 ± 0.10 | 0.46 ± 0.08 |
| Spleen | 0.26 ± 0.04 | 0.25 ± 0.05 | 0.29 ± 0.04 | 0.28 ± 0.03 | 0.27 ± 0.05 | 0.28 ± 0.04 |
| Kidney | 1.09 ± 0.06 | 1.12 ± 0.05 | 1.16 ± 0.06 | 0.90 ± 0.05 b | 0.96 ± 0.06 | 1.02 ± 0.06 |
| Pancreas | 0.47 ± 0.03 | 0.48 ± 0.03 | 0.50 ± 0.04 | 0.42 ± 0.04 | 0.46 ± 0.05 | 0.48 ± 0.04 |
| Adopose tissue | ||||||
| Epididymal | 2.10 ± 0.20 | 2.16 ± 0.30 | 3.70 ± 0.62 a | 5.64 ± 0.48 b | 5.15 ± 0.50 c | 4.62 ± 0.54 d |
| Subcutaneous | 1.26 ± 0.24 | 1.32 ± 0.28 | 2.54 ± 0.40 a | 3.78 ± 0.39 b | 3.24 ± 0.36 | 2.64 ± 0.30 d |
| Retroperitoneal | 0.50 ± 0.10 | 0.52 ± 0.09 | 0.90 ± 0.12 a | 1.82 ± 0.11 b | 1.54 ± 0.11 | 1.26 ± 0.13 d |
| Mesenteric | 1.28 ± 0.14 | 1.28 ± 0.20 | 2.12 ± 0.26 a | 3.22 ± 0.28 b | 2.86 ± 0.24 | 2.46 ± 0.24 d |
| Total | 5.14 ± 0.45 | 5.28 ± 0.56 | 9.26 ± 0.92 a | 14.46 ± 1.12 b | 12.79 ± 1.04 c | 10.98 ± 0.98 d |
| Ingredient | Low-Fat Diet Amount (g/kg) (LAD 3001M) | High-Fat Diet Amount (g/kg) (TP 02420X) |
|---|---|---|
| Casein | 139 | 172 |
| Corn starch | 530.992 | 267.960 |
| Maltodextrin | 109 | 135 |
| Sucrose | 82 | 101 |
| Soybean oil | 40 | 202 |
| Cellulose | 50 | 62 |
| Mineral and vitamin mix | 45 | 55 |
| L-Cystine | 2 | 2 |
| Choline bitartrate | 2 | 3 |
| tert-Butylhydroquinone | 0.008 | 0.04 |
| Total | 1000 | 1000 |
| Genes | Direction | Sequences |
|---|---|---|
| SREBP1 | Forward | 5′-AGGTGTATTTGCTGGCTTGGT-3′ |
| Reverse | 5′-AGAGATGACTAGGGAACTGTGTGT-3′ | |
| SREBP2 | Forward | 5′-AGAAAGAGCGGTGGAGTCCTTG-3′ |
| Reverse | 5′-GAACTGCTGGAGAATGGTGAGG-3′ | |
| FASN | Forward | 5′-GGAGGTTGCTTGGAAGAG-3′ |
| Reverse | 5′- CTGGATGTGATCGAATGCT-3′ | |
| SCD1 | Forward | 5′-GCTGGGCAGGAACTAGTGAG-3′ |
| Reverse | 5′-GAAGGCATGGAAGGTTCAAA-3′ | |
| HMGCS2 | Forward | 5′-GCCGTGAACTGGGTCGAA-3′ |
| Reverse | 5′-GCATATATAGCAATGTCTCCTGCAA-3′ | |
| CPT-1a | Forward | 5′-GAGACTTCCAACGCATGACA-3′ |
| Reverse | 5′-ATGGGTTGGGGTGATGTAGA-3′ | |
| NLRP3 | Forward | 5′-AACAGCCACCTCACTTCCAG-3′ |
| Reverse | 5′-CCAACCACAATCTCCGAATG-3′ | |
| Caspase−1 | Forward | 5′-GCACAAGACCTCTGACAGCA-3′ |
| Reverse | 5′-TTGGGCAGTTCTTGGTATTC-3′ | |
| IL−1β | Forward | 5′-CCTGTCCTGCGTGTTGAAAGA-3′ |
| Reverse | 5′-GGGAACTGGGCAGACTCAAA-3′ | |
| IL−18 | Forward | 5′-CCTTCCATCCTTCACAGATAGG-3′ |
| Reverse | 5′-CCTGATATCGACCGAACAGC-3′ | |
| GAPDH | Forward | 5′- TCATTGACCTCAACTACATGGT-3′ |
| Reverse | 5′-CTAAGCAGTTGGTGGTGCAG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Zhong, X.; Wang, T.; Wu, S.; Guan, H.; Wang, D. (−)-Epigallocatechin-3-Gallate Reduces Perfluorodecanoic Acid-Exacerbated Adiposity and Hepatic Lipid Accumulation in High-Fat Diet-Fed Male C57BL/6J Mice. Molecules 2023, 28, 7832. https://doi.org/10.3390/molecules28237832
Xu H, Zhong X, Wang T, Wu S, Guan H, Wang D. (−)-Epigallocatechin-3-Gallate Reduces Perfluorodecanoic Acid-Exacerbated Adiposity and Hepatic Lipid Accumulation in High-Fat Diet-Fed Male C57BL/6J Mice. Molecules. 2023; 28(23):7832. https://doi.org/10.3390/molecules28237832
Chicago/Turabian StyleXu, Hong, Xu Zhong, Taotao Wang, Shanshan Wu, Huanan Guan, and Dongxu Wang. 2023. "(−)-Epigallocatechin-3-Gallate Reduces Perfluorodecanoic Acid-Exacerbated Adiposity and Hepatic Lipid Accumulation in High-Fat Diet-Fed Male C57BL/6J Mice" Molecules 28, no. 23: 7832. https://doi.org/10.3390/molecules28237832
APA StyleXu, H., Zhong, X., Wang, T., Wu, S., Guan, H., & Wang, D. (2023). (−)-Epigallocatechin-3-Gallate Reduces Perfluorodecanoic Acid-Exacerbated Adiposity and Hepatic Lipid Accumulation in High-Fat Diet-Fed Male C57BL/6J Mice. Molecules, 28(23), 7832. https://doi.org/10.3390/molecules28237832