

RNA-Seq-Based Transcriptomics and GC–MS Quantitative Analysis Reveal Antifungal Mechanisms of Essential Oil of Clausena lansium (Lour.) Skeels Seeds against Candida albicans

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
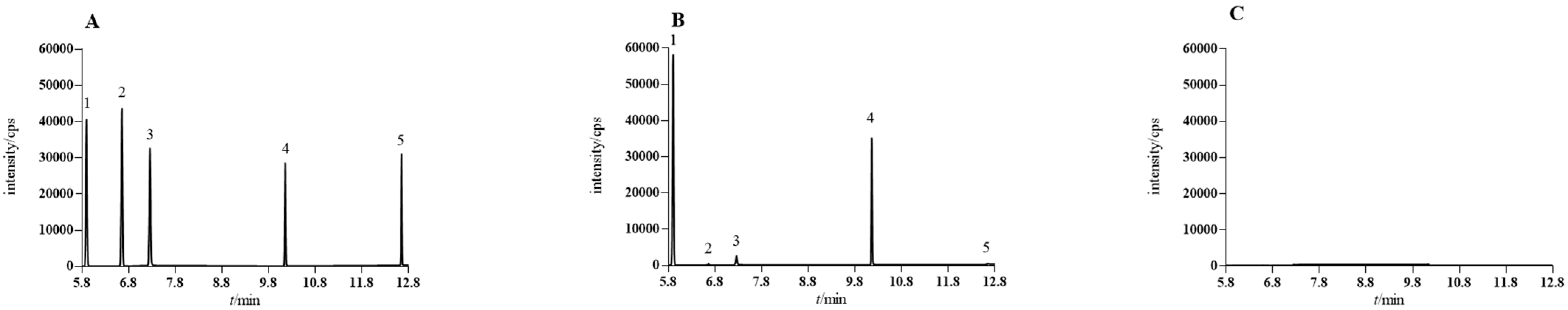
2.1. CSEO Extraction and Analysis
2.2. Antifungal Effects of Volatile Components
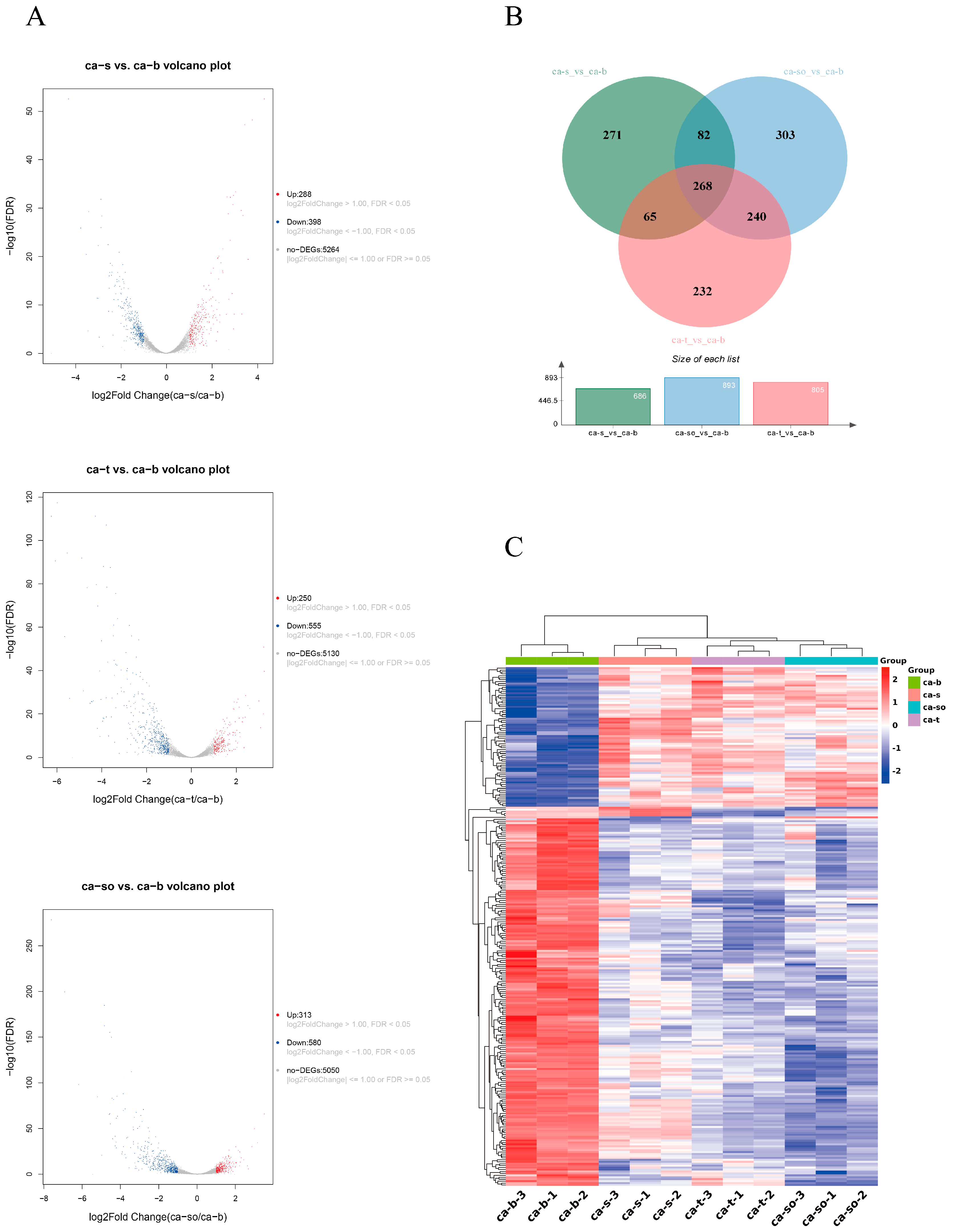
2.3. Analysis of DEGs
2.4. GO and KEGG Enrichment Analysis
3. Materials and Methods
3.1. Chemicals and Plant Materials
3.2. Strains and Culture Conditions
3.3. Extraction of CSEO
3.4. GC–MS Analysis of the Five Components of CSEO
3.5. Determination of Minimal Inhibitory Concentrations (MICs) and Minimum Fungicidal Concentrations (MFCs)
3.6. Effects of CSEO, Sabinene, and 4-Terpineol on the Kinetic Growth of C. albicans Strains
3.7. Transcriptome Determination
3.8. Bioinformatics Transcriptome Analysis
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EO | Essential oil |
| CSEO | Essential oil of Clausena lansium (Lour.) Skeels seeds |
| GC-MS | Gas chromatography–mass spectrometry |
| DMSO | Dimethyl sulfoxide |
| SDA | Sabouraud dextrose agar |
| EI | Electron impact ionization |
| MIC | Minimal inhibitory concentration |
| MFC | Minimum fungicidal concentration |
| FPKM | Fragments per kilobase of exon model per million mapped reads |
| DEGs | Differential expression genes |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LOQ | Limit of quantification |
| LOD | Limit of detection |
| S/N | Signal-to-noise ratio |
| r | Correlation coefficient |
| RSD | Relative standard deviation value |
| BP | Biological process |
| CC | Cellular component |
| MF | Molecular function |
References
- Macias-Paz, I.; Pérez-Hernández, S.; Tavera-Tapia, A.; Luna-Arias, J.; Guerra-Cárdenas, J.; Reyna-Beltrán, E. Candida albicans the main opportunistic pathogenic fungus in humans. Rev. Argent. Microbiol. 2022, 55, 189–198. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhu, R.; Luan, Z.; Ma, X. Risk of invasive candidiasis with prolonged duration of ICU stay: A systematic review and meta-analysis. BMJ Open 2020, 10, e036452. [Google Scholar] [CrossRef]
- Li, T.; Wang, Y.; Yang, S.; Law, Y.; Shiu, B.; Chen, T.; Wu, S.; Lu, M. Risk factors and prognoses of invasive Candida infection in surgical critical ill patients with perforated peptic ulcer. J. Microbiol. Immun. Infect. 2022, 55, 740–748. [Google Scholar] [CrossRef]
- Talapko, J.; Juzbašić, M.; Matijević, T.; Pustijanac, E.; Bekić, S.; Kotris, I.; Škrlec, I. Candida albicans- the virulence factors and clinical manifestations of infection. J. Fungi 2021, 7, 79. [Google Scholar] [CrossRef]
- Pristov, K.; Ghannoum, M. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef]
- Dahiya, S.; Sharma, N.; Punia, A.; Choudhary, P.; Gulia, P.; Parmar, V.; Chhillar, A. Antimycotic drugs and their mechanisms of resistance to Candida species. Curr. Drug Targets 2022, 23, 116–125. [Google Scholar] [CrossRef]
- Iyer, K.; Robbins, N.; Cowen, L. The role of Candida albicans stress response pathways in antifungal tolerance and resistance. iScience 2022, 25, 103953. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, C.; Zhao, X.; Wang, D.; Liu, Y.; Sun, S. Antifungal activity and potential mechanism of Asiatic acid alone and in combination with fluconazole against Candida albicans. Biomed. Pharmacother. 2021, 139, 111568. [Google Scholar] [CrossRef]
- Candiracci, M.; Citterio, B.; Piatti, E. Antifungal activity of the honey flavonoid extract against Candida albicans. Food Chem. 2012, 131, 493–499. [Google Scholar] [CrossRef]
- Semerdjieva, I.; Zheljazkov, V.; Dincheva, I.; Astatkie, T.; Kačániová, M. Chemotypes of Juniperus oxycedrus in Bulgaria and the antimicrobial activity of galbuli essential oils. Ind. Crop. Prod. 2020, 158, 113005. [Google Scholar] [CrossRef]
- Chraibi, M.; Fadil, M.; Farah, A.; Lebrazi, S.; Fikri-Benbrahim, K. Antimicrobial combined action of Mentha pulegium, Ormenis mixta and Mentha piperita essential oils against S. aureus, E. coli and C. tropicalis: Application of mixture design methodology. LWT-Food Sci. Technol. 2021, 145, 111352. [Google Scholar] [CrossRef]
- Hristova, Y.; Wanner, J.; Jirovetz, L.; Stappen, I.; Iliev, I.; Gochev, V. Chemical composition and antifungal activity of essential oil of Hyssopus officinalis L. from Bulgaria against clinical isolates of Candida species. Biotechnol. Biotechnol. Equip. 2015, 29, 592–601. [Google Scholar] [CrossRef]
- Jafri, H.; Ahmad, I. Thymus vulgaris essential oil and thymol inhibit biofilms and interact synergistically with antifungal drugs against drug resistant strains of Candida albicans and Candida tropicalis. J. Mycol. Med. 2020, 30, 100911. [Google Scholar] [CrossRef]
- Lim, T. Clausena lansium. In Edible Medicinal and Non-Medicinal Plants; Springer: Dordrecht, The Netherlands, 2012; Volume 4, pp. 871–883. [Google Scholar]
- Peng, W.; Fu, X.; Xiong, Z.; Xiang, M.; Yang, Y.; Wu, H.; Shi, X.; Huo, G.; Cui, R.; Li, B. Chemical components from the stems and leaves of Clausena lansium Lour. Skeels and their potential herbicidal effects. Pest Manag. Sci. 2021, 77, 1355–1360. [Google Scholar] [CrossRef]
- Phachonpai, W.; Tongun, T. Cognition enhancing effects of Clausena lansium (Lour.) peel extract attenuate chronic restraint stress-induced memory deficit in rats. Heliyon 2021, 7, e07003. [Google Scholar] [CrossRef]
- Yan, H.; Xiong, Z.; Xie, N.; Liu, S.; Zhang, L.; Xu, F.; Guo, W.; Feng, J. Bioassay-guided isolation of antifungal amides against Sclerotinia sclerotiorum from the seeds of Clausena lansium. Ind. Crop. Prod. 2018, 121, 352–359. [Google Scholar] [CrossRef]
- Fan, F.; Liu, Y.; Liu, Y.; Lv, R.; Sun, W.; Ding, W.; Cai, Y.; Li, W.; Liu, X.; Qu, W. Candida albicans biofilms: Antifungal resistance, immune evasion, and emerging therapeutic strategies. Int. J. Antimicrob. Agents 2022, 60, 106673. [Google Scholar] [CrossRef]
- Shen, S.; Liao, Q.; Huang, L.; Li, D.; Zhang, Q.; Wang, Y.; Lee, S.; Lin, L. Water soluble fraction from ethanolic extract of Clausena lansium seeds alleviates obesity and insulin resistance and changes the composition of gut microbiota in high-fat diet-fed mice. J. Funct. Foods 2018, 47, 192–199. [Google Scholar] [CrossRef]
- Zhao, J.; Nan, P.; Zhong, Y. Chemical composition of the essential oils of Clausena lansium from Hainan island, China. Z. Naturforsch. C 2004, 59, 153–156. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, Y.; Zhou, X.; Yang, H.; Zhang, H.; Chen, W.; Zhang, H.; Zhang, Y.; He, X. The influence of the chemical composition of essential oils of Clausena lansium seeds on the growth of Candida strains. Sci. Rep. 2021, 11, 19666. [Google Scholar] [CrossRef]
- He, X.; Ma, Y.; Yi, G.; Wen, Y.; Zhang, Y.; Zhang, H.; Zhang, L. Rapid and sensitive analysis of volatile components of different parts of Clausena lansium by ionic liquid based headspace gas chromatography-mass spectrometry. Molecules 2019, 24, 91. [Google Scholar] [CrossRef] [PubMed]
- Araújo, D.; Henriques, M.; Silva, S. Portrait of Candida species biofilm regulatory network genes. Trends Microbiol. 2017, 25, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Wang, J.; Chu, M.; Jia, C. Activity of coumarin against Candida albicans biofilms. J. Mycol. Med. 2018, 29, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Chen, H.; Mei, W.; Kong, F.; Li, F.; Chen, P.; Cai, C.; Huang, M.; Dai, H. Nematicidal amide alkaloids from the seeds of Clausena lansium. Fitoterapia 2017, 128, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, L. The ALS gene family of Candida albicans. Trends Microbiol. 2001, 9, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Finkel, J.S.; Mitchell, A.P. Genetic control of Candida albicans biofilm development. Nat. Rev. Microbiol. 2011, 9, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Fernanda, F.; Barros, P.; Pereira, C.; Junqueira, J.; Jorge, A. Photodynamic inactivation in the expression of the Candida albicans genes ALS3, HWP1, BCR1, TEC1, CPH1, and EFG1 in biofilms. Laser Med. Sci. 2018, 33, 1447–1454. [Google Scholar]
- Orsi, C.; Borghi, E.; Colombari, B.; Neglia, R.; Quaglino, D.; Ardizzoni, A.; Morace, G.; Blasi, E. Impact of Candida albicans hyphal wall protein 1 (HWP1) genotype on biofilm production and fungal susceptibility to microglial cells. Microb. Pathog. 2014, 69–70, 20–27. [Google Scholar] [CrossRef]
- Naglik, J.; Challacombe, S.; Hube, B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef]
- Gergondey, R.; Garcia, C.; Serre, V.; Camadro, J.M.; Auchère, F. The adaptive metabolic response involves specific protein glutathionylation during the filamentation process in the pathogen Candida albicans. BBA—Mol. Basis Dis. 2016, 1862, 1309–1323. [Google Scholar] [CrossRef]
- CLSI Document M38-A2; Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Retention Time (min) | Linear Range (μg·mL−1) | Slope | Intercept | r | LOQ (ng·mL−1) | LOD (ng·mL−1) |
|---|---|---|---|---|---|---|---|
| Sabinene | 5.89 | 0.25–31.50 | 15,960 | 198.14 | 0.9998 | 50.40 | 10.08 |
| α-phellandrene | 6.65 | 0.28–35.02 | 13,466 | −1049.30 | 0.9990 | 56.03 | 11.21 |
| β-phellandrene | 7.25 | 0.27–33.55 | 10,534 | 565.17 | 0.9997 | 53.68 | 10.74 |
| 4-terpineol | 10.15 | 0.26–32.93 | 8789.5 | −660.18 | 0.9999 | 52.68 | 10.54 |
| β-caryophyllene | 12.65 | 0.25–32.40 | 5958.1 | 1349.6 | 0.9997 | 103.68 | 20.74 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Sui, J.; Wang, Y.; Sun, W.; Yi, G.; Wu, J.; Qiu, S.; Wang, L.; Zhang, A.; He, X. RNA-Seq-Based Transcriptomics and GC–MS Quantitative Analysis Reveal Antifungal Mechanisms of Essential Oil of Clausena lansium (Lour.) Skeels Seeds against Candida albicans. Molecules 2023, 28, 8052. https://doi.org/10.3390/molecules28248052
Ma Y, Sui J, Wang Y, Sun W, Yi G, Wu J, Qiu S, Wang L, Zhang A, He X. RNA-Seq-Based Transcriptomics and GC–MS Quantitative Analysis Reveal Antifungal Mechanisms of Essential Oil of Clausena lansium (Lour.) Skeels Seeds against Candida albicans. Molecules. 2023; 28(24):8052. https://doi.org/10.3390/molecules28248052
Chicago/Turabian StyleMa, Yinzheng, Jinlei Sui, Yan Wang, Wanying Sun, Guohui Yi, Jinyan Wu, Shi Qiu, Lili Wang, Aihua Zhang, and Xiaowen He. 2023. "RNA-Seq-Based Transcriptomics and GC–MS Quantitative Analysis Reveal Antifungal Mechanisms of Essential Oil of Clausena lansium (Lour.) Skeels Seeds against Candida albicans" Molecules 28, no. 24: 8052. https://doi.org/10.3390/molecules28248052
APA StyleMa, Y., Sui, J., Wang, Y., Sun, W., Yi, G., Wu, J., Qiu, S., Wang, L., Zhang, A., & He, X. (2023). RNA-Seq-Based Transcriptomics and GC–MS Quantitative Analysis Reveal Antifungal Mechanisms of Essential Oil of Clausena lansium (Lour.) Skeels Seeds against Candida albicans. Molecules, 28(24), 8052. https://doi.org/10.3390/molecules28248052