
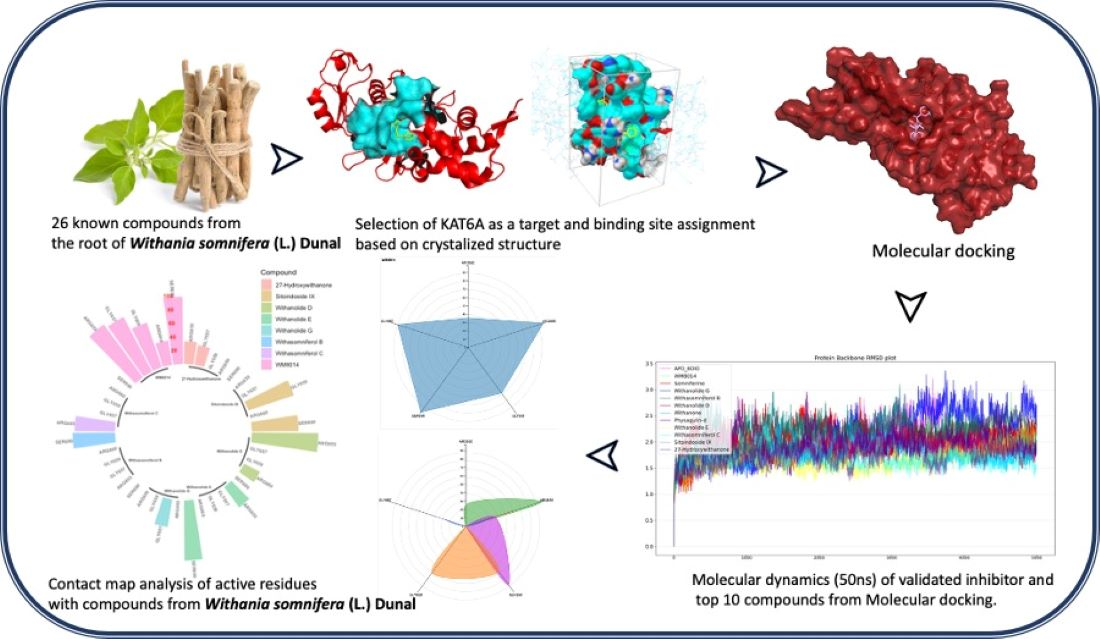
In Silico Study on the Interactions, Molecular Docking, Dynamics and Simulation of Potential Compounds from Withania somnifera (L.) Dunal Root against Cancer by Targeting KAT6A

 , , , , , ,
, , , , , ,  , ,
, ,
Abstract
:
1. Introduction
2. Results
2.1. ADMET Properties
2.2. Binding Site Assignment
2.3. Molecular Docking Results
Molecular Dynamics (MD)
3. Discussion
4. Materials and Methods
4.1. Data Source
4.2. Prediction of ADMET Properties and Network Analysis
4.3. Molecular Docking
4.3.1. Protein Preparation
4.3.2. Identification of Binding Cavity
4.3.3. Ligand Preparation
4.3.4. Docking of the Compounds Using the POAP Program
4.4. Molecular Dynamics (MD)
4.5. In Vitro Studies
4.5.1. Materials Used
4.5.2. Extraction Method
4.5.3. Analysis of Anticancer Properties by 3-(4, 5-Dimethylthiazol2-yl)−2,5-Diphenyltetrazolium Bromide (MTT) Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug Resistance in Cancer: An Overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- vel Szic, K.S.; Palagani, A.; Hassannia, B.; Sabbe, L.; Heyninck, K.; Haegeman, G.; Vanden, W. Phytochemicals and Cancer Chemoprevention: Epigenetic Friends or Foe? In Phytochemicals-Bioactivities and Impact on Health; InTech: London, UK, 2011. [Google Scholar]
- Ghanbari, M.; Safaralizadeh, R.; Mohammadi, K. A Review on Important Histone Acetyltransferase (HAT) Enzymes as Targets for Cancer Therapy. Curr. Cancer Ther. Rev. 2019, 15, 120–130. [Google Scholar] [CrossRef]
- Baell, J.B.; Leaver, D.J.; Hermans, S.J.; Kelly, G.L.; Brennan, M.S.; Downer, N.L.; Nguyen, N.; Wichmann, J.; McRae, H.M.; Yang, Y.; et al. Inhibitors of Histone Acetyltransferases KAT6A/B Induce Senescence and Arrest Tumour Growth. Nature 2018, 560, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Mitsopoulos, C.; Di Micco, P.; Fernandez, E.V.; Dolciami, D.; Holt, E.; Mica, I.L.; Coker, E.A.; Tym, J.E.; Campbell, J.; Che, K.H.; et al. CanSAR: Update to the Cancer Translational Research and Drug Discovery Knowledgebase. Nucleic Acids Res. 2021, 49, D1074–D1082. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Ankad, G.M.; Pai, S.R.; Hiremath, J.; Hegde, H.V. Traditional Horticulture Practices Increase the Production of Selected Withanolides in Withania somnifera (L.) Dunal—A RP-UFLC Analysis. J. Chromatogr. Sci. 2020, 58, 899–906. [Google Scholar] [CrossRef]
- Wang, J.; Li, J.; Li, J.; Li, J.; Liu, S.; Huang, L.; Gao, W. Production of Active Compounds in Medicinal Plants: From Plant Tissue Culture to Biosynthesis. Chin. Herb. Med. 2017, 9, 115–125. [Google Scholar] [CrossRef]
- Bhuiyan, F.R.; Howlader, S.; Raihan, T.; Hasan, M. Plants Metabolites: Possibility of Natural Therapeutics Against the COVID-19 Pandemic. Front. Med. 2020, 7, 444. [Google Scholar] [CrossRef]
- Kala, C.P.; Dhyani, P.P.; Sajwan, B.S. Developing the Medicinal Plants Sector in Northern India: Challenges and Opportunities. J. Ethnobiol. Ethnomed. 2006, 2, 32. [Google Scholar] [CrossRef]
- Kulkarni, S.K.; Dhir, A. Withania somnifera: An Indian Ginseng. Prog. Neuro-Psychopharmacol. Biol. Psychiatr. 2008, 32, 1093–1105. [Google Scholar] [CrossRef]
- Bagchi, C.; Das, S.; Mitra, A.; Pati, A.; Tripathi, S.; Datta, A. Antidiabetic and Antihyperlipidemic Activity of Hydroalcoholic Extract of Withania coagulans Dunal Dried Fruit in Experimental Rat Models. J. Ayurveda Integr. Med. 2013, 4, 99. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, D.; Bhattacharyya, S.; Bose, S. Efficacy and Safety of Ashwagandha (Withania somnifera (L.) Dunal) Root Extract in Improving Memory and Cognitive Functions. J. Diet. Suppl. 2017, 14, 599–612. [Google Scholar] [CrossRef]
- Pingali, U.; Pilli, R.; Fatima, N. Effect of Standardized Aqueous Extract of Withania somnifera on Tests of Cognitive and Psychomotor Performance in Healthy Human Participants. Pharmacognosy Res. 2014, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Devi, P.U.; Sharada, A.C.; Solomon, F.E. In Vivo Growth Inhibitory and Radiosensitizing Effects of Withaferin A on Mouse Ehrlich Ascites Carcinoma. Cancer Lett. 1995, 95, 189–193. [Google Scholar] [CrossRef]
- Mehta, V.; Chander, H.; Munshi, A. Mechanisms of Anti-Tumor Activity of Withania somnifera (Ashwagandha). Nutr. Cancer 2021, 73, 914–926. [Google Scholar] [CrossRef]
- Haider, B.; Thakur, S.S. Withania somnifera Has Potential to Treat Cancer. In Science of Ashwagandha: Preventive and Therapeutic Potentials; Springer: Cham, Switzerland, 2017; pp. 213–551. [Google Scholar] [CrossRef]
- Lee, B.S.; Yoo, M.J.; Kang, H.; Lee, S.R.; Kim, S.; Yu, J.S.; Kim, J.-C.; Jang, T.S.; Pang, C.; Kim, K.H. Withasomniferol D, a New Anti-Adipogenic Withanolide from the Roots of Ashwagandha (Withania somnifera). Pharmaceuticals 2021, 14, 1017. [Google Scholar] [CrossRef]
- Balkrishna, A.; Nain, P.; Chauhan, A.; Sharma, N.; Gupta, A.; Ranjan, R.; Varshney, A. Super Critical Fluid Extracted Fatty Acids from Withania somnifera Seeds Repair Psoriasis-Like Skin Lesions and Attenuate Pro-Inflammatory Cytokines (TNF-α and IL-6) Release. Biomolecules 2020, 10, 185. [Google Scholar] [CrossRef] [Green Version]
- Maher, S.; Choudhary, M.I.; Saleem, F.; Rasheed, S.; Waheed, I.; Halim, S.A.; Azeem, M.; Abdullah, I.B.; Froeyen, M.; Mirza, M.U.; et al. Isolation of Antidiabetic Withanolides from Withania coagulans Dunal and Their In Vitro and In Silico Validation. Biology 2020, 9, 197. [Google Scholar] [CrossRef]
- White, P.T.; Subramanian, C.; Motiwala, H.F.; Cohen, M.S. Natural Withanolides in the Treatment of Chronic Diseases. In Anti-inflammatory Nutraceuticals and Chronic Diseases; Gupta, S., Prasad, S., Aggarwal, B., Eds.; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2016; Volume 928, pp. 329–373. [Google Scholar]
- Prasad, K.S.; Prasad, S.K.; Veerapur, R.; Lamraoui, G.; Prasad, A.; Prasad, M.N.N.; Singh, S.K.; Marraiki, N.; Syed, A.; Shivamallu, C. Antitumor Potential of Green Synthesized ZnONPs Using Root Extract of Withania somnifera against Human Breast Cancer Cell Line. Separations 2021, 8, 8. [Google Scholar] [CrossRef]
- Gurung, A.B.; Ali, M.A.; Lee, J.; Farah, M.A.; Al-Anazi, K.M. Molecular Docking and Dynamics Simulation Study of Bioactive Compounds from Ficus Carica L. with Important Anticancer Drug Targets. PLoS ONE 2021, 16, e0254035. [Google Scholar] [CrossRef] [PubMed]
- Tewari, D.; Chander, V.; Dhyani, A.; Sahu, S.; Gupta, P.; Patni, P.; Kalick, L.S.; Bishayee, A. Withania somnifera (L.) Dunal: Phytochemistry, Structure-Activity Relationship, and Anticancer Potential. Phytomedicine 2022, 98, 153949. [Google Scholar] [CrossRef] [PubMed]
- Holbert, M.A.; Sikorski, T.; Carten, J.; Snowflack, D.; Hodawadekar, S.; Marmorstein, R. The Human Monocytic Leukemia Zinc Finger Histone Acetyltransferase Domain Contains DNA-Binding Activity Implicated in Chromatin Targeting. J. Biol. Chem. 2007, 282, 36603–36613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-C.; Tsai, Y.-L.; Wu, Y.-C.; Chang, F.-R.; Liu, M.-H.; Chen, W.-Y.; Wu, C.-C. Withanolides-Induced Breast Cancer Cell Death Is Correlated with Their Ability to Inhibit Heat Protein 90. PLoS ONE 2012, 7, e37764. [Google Scholar] [CrossRef] [PubMed]
- Saglam, O.; Tang, Z.; Tang, G.; Medeiros, L.J.; Toruner, G.A. KAT6A Amplifications Are Associated with Shorter Progression-Free Survival and Overall Survival in Patients with Endometrial Serous Carcinoma. PLoS ONE 2020, 15, e0238477. [Google Scholar] [CrossRef]
- Sheikh, B.N.; Lee, S.C.W.; El-Saafin, F.; Vanyai, H.K.; Hu, Y.; Pang, S.H.M.; Grabow, S.; Strasser, A.; Nutt, S.L.; Alexander, W.S.; et al. MOZ Regulates B-Cell Progenitors and, Consequently, Moz Haploinsufficiency Dramatically Retards MYC-Induced Lymphoma Development. Blood 2015, 125, 1910–1921. [Google Scholar] [CrossRef]
- Singh, N.; Bhalla, M.; De Jager, P.; Gilca, M. An Overview on Ashwagandha: A Rasayana (Rejuvenator) of Ayurveda. African J. Tradit. Complement. Altern. Med. 2011, 8, 208–213. [Google Scholar] [CrossRef]
- Agarwal, R.; Diwanay, S.; Patki, P.; Patwardhan, B. Studies on Immunomodulatory Activity of Withania somnifera (Ashwagandha) Extracts in Experimental Immune Inflammation. J. Ethnopharmacol. 1999, 67, 27–35. [Google Scholar] [CrossRef]
- Gurav, S.; Wanjari, M.; Bhole, R.; Raut, N.; Prasad, S.; Saoji, S.; Chikhale, R.; Khanal, P.; Pant, A.; Ayyanar, M.; et al. Ethnological Validation of Ashwagandha (Withania somnifera L. Dunal) Ghrita as ‘Vajikarana Rasayana’: In-Silico, in-Vitro and in-Vivo Approach. J. Ethnopharmacol. 2023, 304, 116064. [Google Scholar] [CrossRef]
- Sajida; Prabhu, A. Anti-Angiogenic, Apoptotic and Matrix Metalloproteinase Inhibitory Activity of Withania somnifera (Ashwagandha) on Lung Adenocarcinoma Cells. Phytomedicine 2021, 90, 153639. [Google Scholar] [CrossRef]
- Nile, S.H.; Nile, A.; Gansukh, E.; Baskar, V.; Kai, G. Subcritical Water Extraction of Withanosides and Withanolides from Ashwagandha (Withania somnifera L) and Their Biological Activities. Food Chem. Toxicol. 2019, 132, 110659. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New Data Content and Improved Web Interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef]
- Degtyarenko, K.; de Matos, P.; Ennis, M.; Hastings, J.; Zbinden, M.; McNaught, A.; Alcantara, R.; Darsow, M.; Guedj, M.; Ashburner, M. ChEBI: A Database and Ontology for Chemical Entities of Biological Interest. Nucleic Acids Res. 2007, 36, D344–D350. [Google Scholar] [CrossRef]
- Mohanraj, K.; Karthikeyan, B.S.; Vivek-Ananth, R.P.; Chand, R.P.B.; Aparna, S.R.; Mangalapandi, P.; Samal, A. IMPPAT: A Curated Database of Indian Medicinal Plants, Phytochemistry and Therapeutics. Sci. Rep. 2018, 8, 4329. [Google Scholar] [CrossRef] [Green Version]
- Pires, D.E.V.; Blundell, T.L.; Ascher, D.B. PkCSM: Predicting Small-Molecule Pharmacokinetic and Toxicity Properties Using Graph-Based Signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef]
- Liu, T.; Lin, Y.; Wen, X.; Jorissen, R.N.; Gilson, M.K. BindingDB: A Web-Accessible Database of Experimentally Determined Protein-Ligand Binding Affinities. Nucleic Acids Res. 2007, 35, D198–D201. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Leaver, D.J.; Cleary, B.; Nguyen, N.; Priebbenow, D.L.; Lagiakos, H.R.; Sanchez, J.; Xue, L.; Huang, F.; Sun, Y.; Mujumdar, P.; et al. Discovery of Benzoylsulfonohydrazides as Potent Inhibitors of the Histone Acetyltransferase KAT6A. J. Med. Chem. 2019, 62, 7146–7159. [Google Scholar] [CrossRef]
- Berman, H.M. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Ravindranath, P.A.; Sanner, M.F. AutoSite: An Automated Approach for Pseudo-Ligands Prediction—From Ligand-Binding Sites Identification to Predicting Key Ligand Atoms. Bioinformatics 2016, 32, 3142–3149. [Google Scholar] [CrossRef] [Green Version]
- UniProt Consortium. UniProt: A Hub for Protein Information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulo, N. The PROSITE Database. Nucleic Acids Res. 2006, 34, D227–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binkowski, T.A. CASTp: Computed Atlas of Surface Topography of Proteins. Nucleic Acids Res. 2003, 31, 3352–3355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samdani, A.; Vetrivel, U. POAP: A GNU Parallel Based Multithreaded Pipeline of Open Babel and AutoDock Suite for Boosted High Throughput Virtual Screening. Comput. Biol. Chem. 2018, 74, 39–48. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An Open Chemical Toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Goodsell, D.S.; Sanner, M.F.; Olson, A.J.; Forli, S. The AutoDock Suite at 30. Protein Sci. 2021, 30, 31–43. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the Scope of the Protein–Ligand Interaction Profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef]
- Bowers, L.; James, K.; Quirk, A.; Simpson, A.; Stewart, D.; Hodsoll, J. Reducing Conflict and Containment Rates on Acute Psychiatric Wards: The Safewards Cluster Randomised Controlled Trial. Int. J. Nurs. Stud. 2015, 52, 1412–1422. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Maxwell, D.S.; Tirado-Rives, J. Development and Testing of the OPLS All-Atom Force Field on Conformational Energetics and Properties of Organic Liquids. J. Am. Chem. Soc. 1996, 118, 11225–11236. [Google Scholar] [CrossRef]
- Shivakumar, D.; Williams, J.; Wu, Y.; Damm, W.; Shelley, J.; Sherman, W. Prediction of Absolute Solvation Free Energies Using Molecular Dynamics Free Energy Perturbation and the OPLS Force Field. J. Chem. Theory Comput. 2010, 6, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant Pressure Molecular Dynamics Algorithms. J. Chem. Phys. 1994, 101, 4177–4189. [Google Scholar] [CrossRef]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA Methods to Estimate Ligand-Binding Affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Bagewadi, Z.K.; Muddapur, U.M.; Madiwal, S.S.; Mulla, S.I.; Khan, A. Biochemical and Enzyme Inhibitory Attributes of Methanolic Leaf Extract of Datura Inoxia Mill. Environ. Sustain. 2019, 2, 75–87. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Binding Energy (Kcal/mol) | Interactions | ||
|---|---|---|---|---|
| Hydrophobic Interaction | Hydrogen Bond | Salt Bridge | ||
| WM-8014 | −10.8 | LEU601, ILE649, ARG660 | ARG660, GLY657, GLY659, ARG690 | |
| Somniferine | −9.6 | GLN760, LYS763 | ARG655, ARG656, GLY657, TYR658, GLY659 | |
| Sitoindoside IX | −9.6 | ARG660, LYS763 | ILE649, GLN654, SER684, LYS763 | |
| Withasomniferol B | −9.2 | ARG655, ARG660, LEU689, TRP697 | ARG655, ARG660 | |
| Withanolide G | −9.1 | ARG660, LEU689, TRP697, LYS763 | ARG655, GLY657 | |
| Withanone | −9 | ARG660, LEU689, TRP697, LYS763 | ARG655, GLY657, LYS763 | |
| Withanolide E | −8.8 | ARG660, LEU689, LYS763, PRO765 | ARG655, GLY657, GLN760 | LYS763 |
| Physagulin-d | −8.6 | ARG660, ILE663, LEU686, LYS763 | ILE647, ILE649, GLN654, SER684 | |
| Withasomniferol C | −8.6 | ARG660, LEU689, LYS763 | ARG655, GLY657, ARG660 | ARG655 |
| Withanolide D | −8.5 | ARG660, GLN760, LYS763 | ARG655, ARG656, GLY657, TYR658, GLY659 | |
| 27-Hydroxywithanone | −8.5 | ARG660, LEU689, ARG692, TRP697, GLN760, LYS763 | ARG655, GLY657, GLN760 | |
| Compound | Total Atoms | Protein RMSD (Å) (Mean) | Ligand RMSD | ΔG (Total Binding Energy) Mean |
|---|---|---|---|---|
| (Å) (Mean) | ||||
| WM-8014 | 37,402 | 1.96 | 0.43 | −88.38 |
| Somniferine | 34,529 | 2.06 | 0.79 | −59.47 |
| Withanolide G | 34,409 | 2.06 | 1.89 | −41.29 |
| Withasomniferol B | 37,273 | 2.16 | 0.56 | −47.62 |
| Withanolide D | 34,544 | 1.90 | 0.88 | −71.16 |
| Withanone | 34,409 | 1.76 | 1.87 | −35.76 |
| Physagulin-d | 34,457 | 1.94 | 1.42 | −41.30 |
| Withanolide E | 34,416 | 1.65 | 0.49 | −48.91 |
| Withasomniferol C | 34,418 | 1.69 | 0.39 | −41.77 |
| Sitoindoside IX | 34,448 | 2.14 | 2.00 | −60.39 |
| 27-Hydroxywithanone | 34,542 | 1.77 | 0.93 | −52.60 |
| Compounds | Total Energy (kcal/mol) | Std. Deviation |
|---|---|---|
| Withanolide D | −89,776.661 | 122.76 |
| Sitoindoside IX | −89,993.379 | 121.87 |
| Withanolide E | −89,973.926 | 123.642 |
| Physagulin-d | −90,020.423 | 122.803 |
| Somniferine | −89,748.161 | 120.236 |
| 27-Hydroxywithanone | −89,940.324 | 122.504 |
| Withanolide G | −89,971.235 | 123.043 |
| Withanone | −90,033.506 | 121.551 |
| Withasomniferol B | −89,663.812 | 125.308 |
| Withasomniferol C | −97,730.765 | 127.169 |
| WM8014 | −97,754.831 | 125.992 |
| APO_60IO | −90,298.600 | 122.209 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deshpande, S.H.; Muhsinah, A.B.; Bagewadi, Z.K.; Ankad, G.M.; Mahnashi, M.H.; Yaraguppi, D.A.; Shaikh, I.A.; Khan, A.A.; Hegde, H.V.; Roy, S. In Silico Study on the Interactions, Molecular Docking, Dynamics and Simulation of Potential Compounds from Withania somnifera (L.) Dunal Root against Cancer by Targeting KAT6A. Molecules 2023, 28, 1117. https://doi.org/10.3390/molecules28031117
Deshpande SH, Muhsinah AB, Bagewadi ZK, Ankad GM, Mahnashi MH, Yaraguppi DA, Shaikh IA, Khan AA, Hegde HV, Roy S. In Silico Study on the Interactions, Molecular Docking, Dynamics and Simulation of Potential Compounds from Withania somnifera (L.) Dunal Root against Cancer by Targeting KAT6A. Molecules. 2023; 28(3):1117. https://doi.org/10.3390/molecules28031117
Chicago/Turabian StyleDeshpande, Sanjay H., Abdullatif Bin Muhsinah, Zabin K. Bagewadi, Gireesh M. Ankad, Mater H. Mahnashi, Deepak A. Yaraguppi, Ibrahim Ahmed Shaikh, Aejaz Abdullatif Khan, Harsha V. Hegde, and Subarna Roy. 2023. "In Silico Study on the Interactions, Molecular Docking, Dynamics and Simulation of Potential Compounds from Withania somnifera (L.) Dunal Root against Cancer by Targeting KAT6A" Molecules 28, no. 3: 1117. https://doi.org/10.3390/molecules28031117
APA StyleDeshpande, S. H., Muhsinah, A. B., Bagewadi, Z. K., Ankad, G. M., Mahnashi, M. H., Yaraguppi, D. A., Shaikh, I. A., Khan, A. A., Hegde, H. V., & Roy, S. (2023). In Silico Study on the Interactions, Molecular Docking, Dynamics and Simulation of Potential Compounds from Withania somnifera (L.) Dunal Root against Cancer by Targeting KAT6A. Molecules, 28(3), 1117. https://doi.org/10.3390/molecules28031117