
Antibacterial Activity of Peptide Derivatives of Phosphinothricin against Multidrug-Resistant Klebsiella pneumoniae

 , , and
, , and
Abstract
:1. Introduction
2. Results
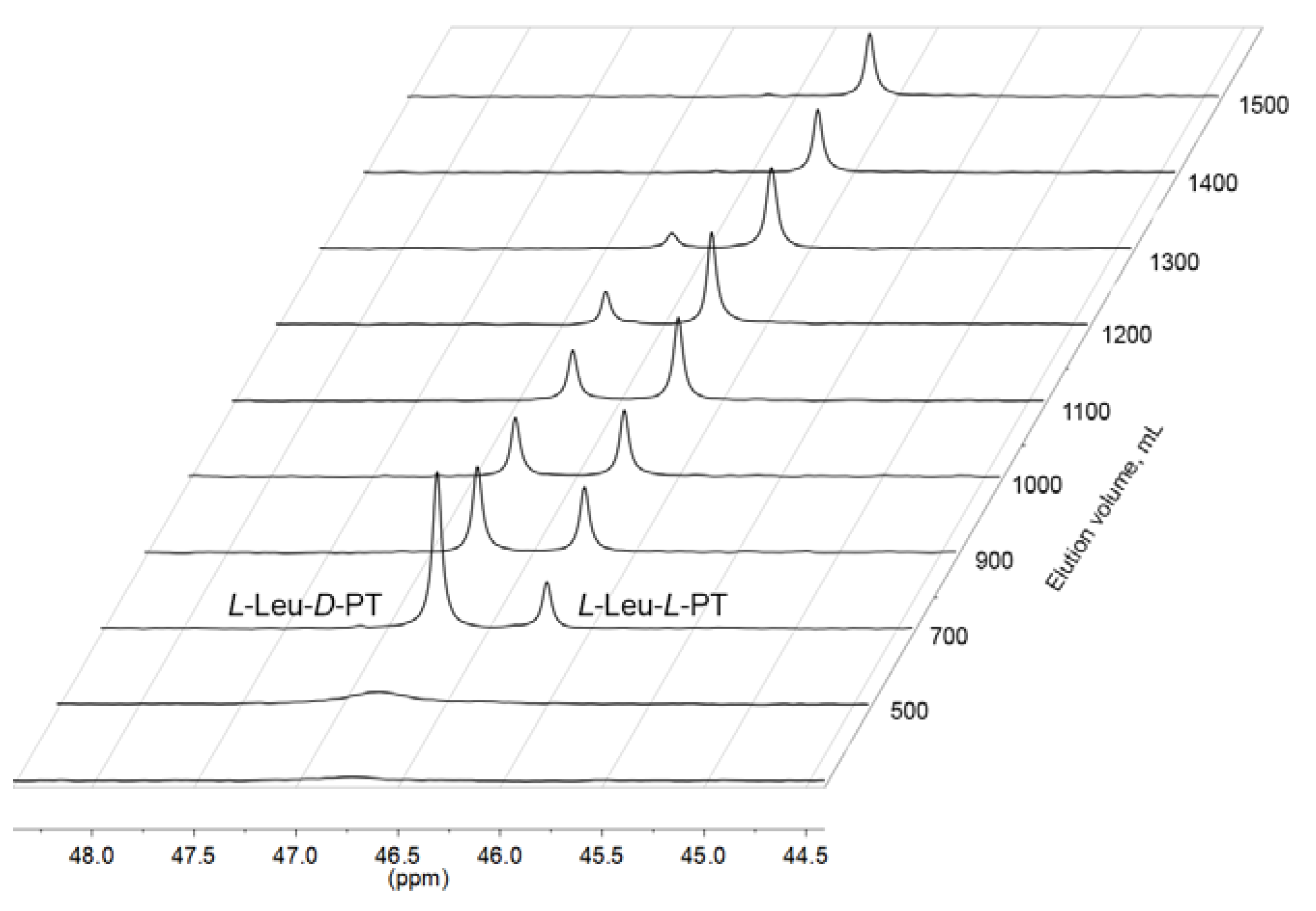
2.1. Synthesis of L-Leu-L-PT
2.2. Bialaphos and L-Leu-L-PT Inhibit the Growth of E. coli K12 in Minimal Medium
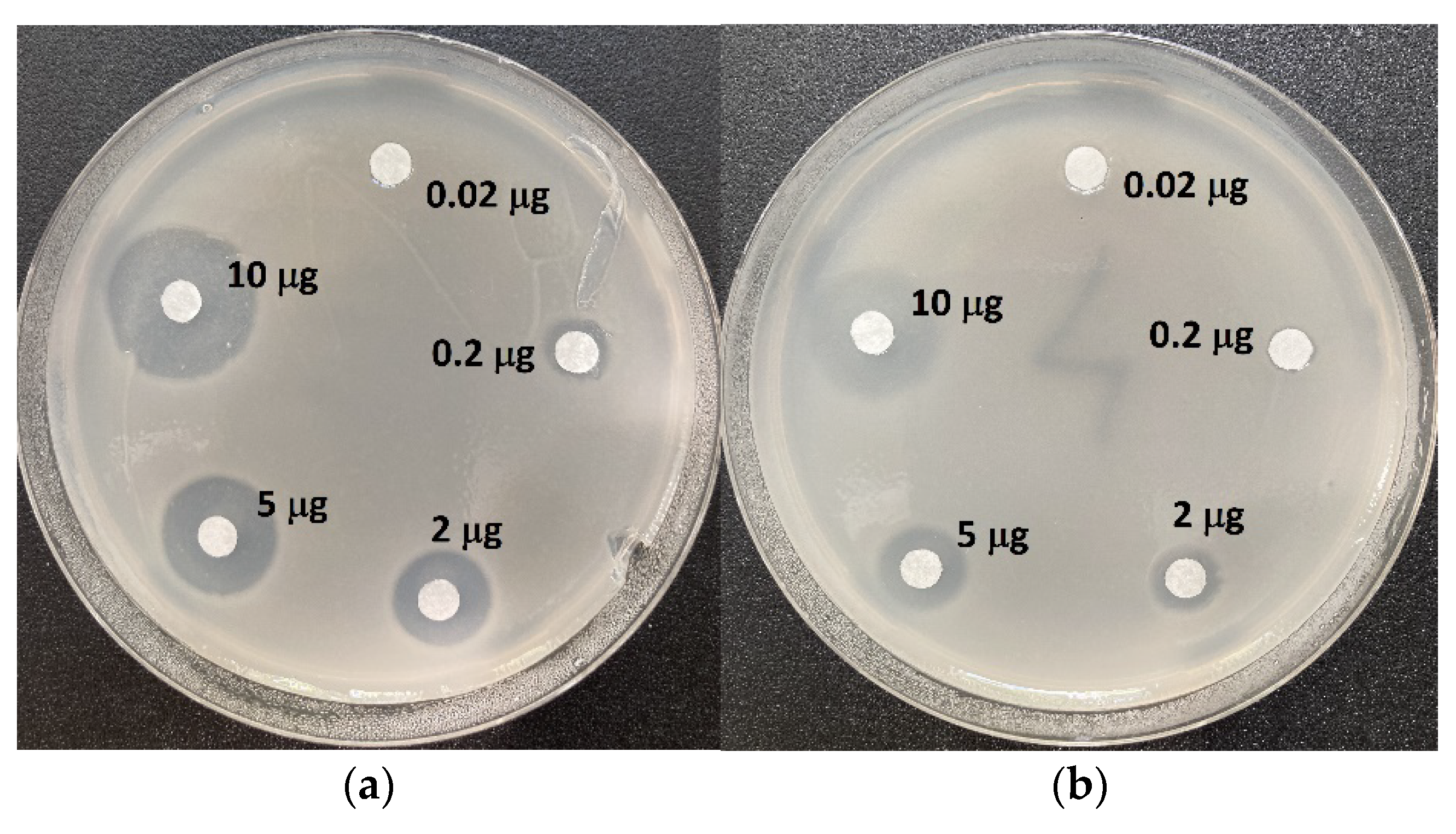
2.3. Inhibition of the Growth of Reference Strain of Klebsiella pneumoniae ATCC 13883 with Bialaphos, L-Leu-L-PT and Antibiotics of Different Classes
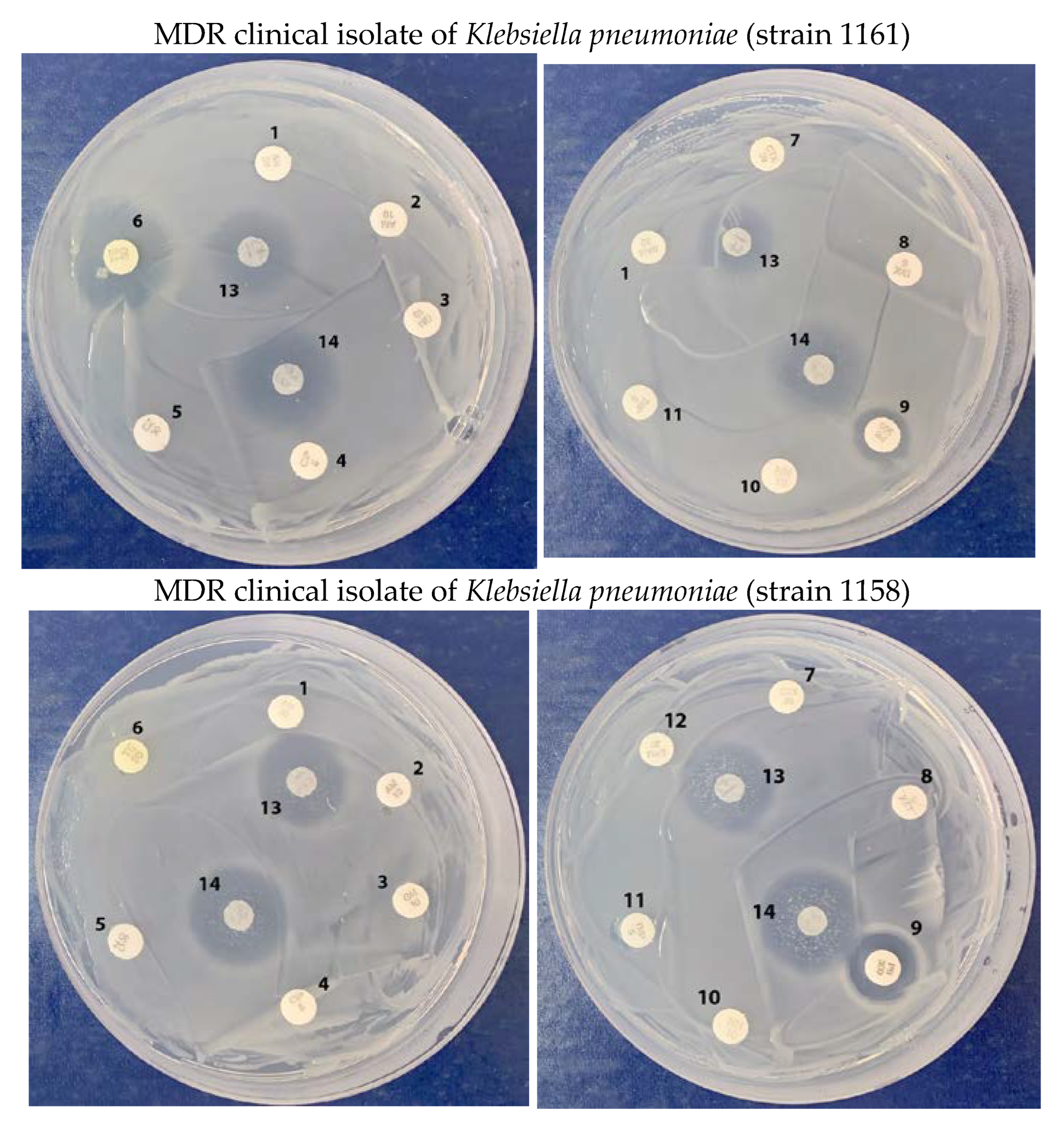
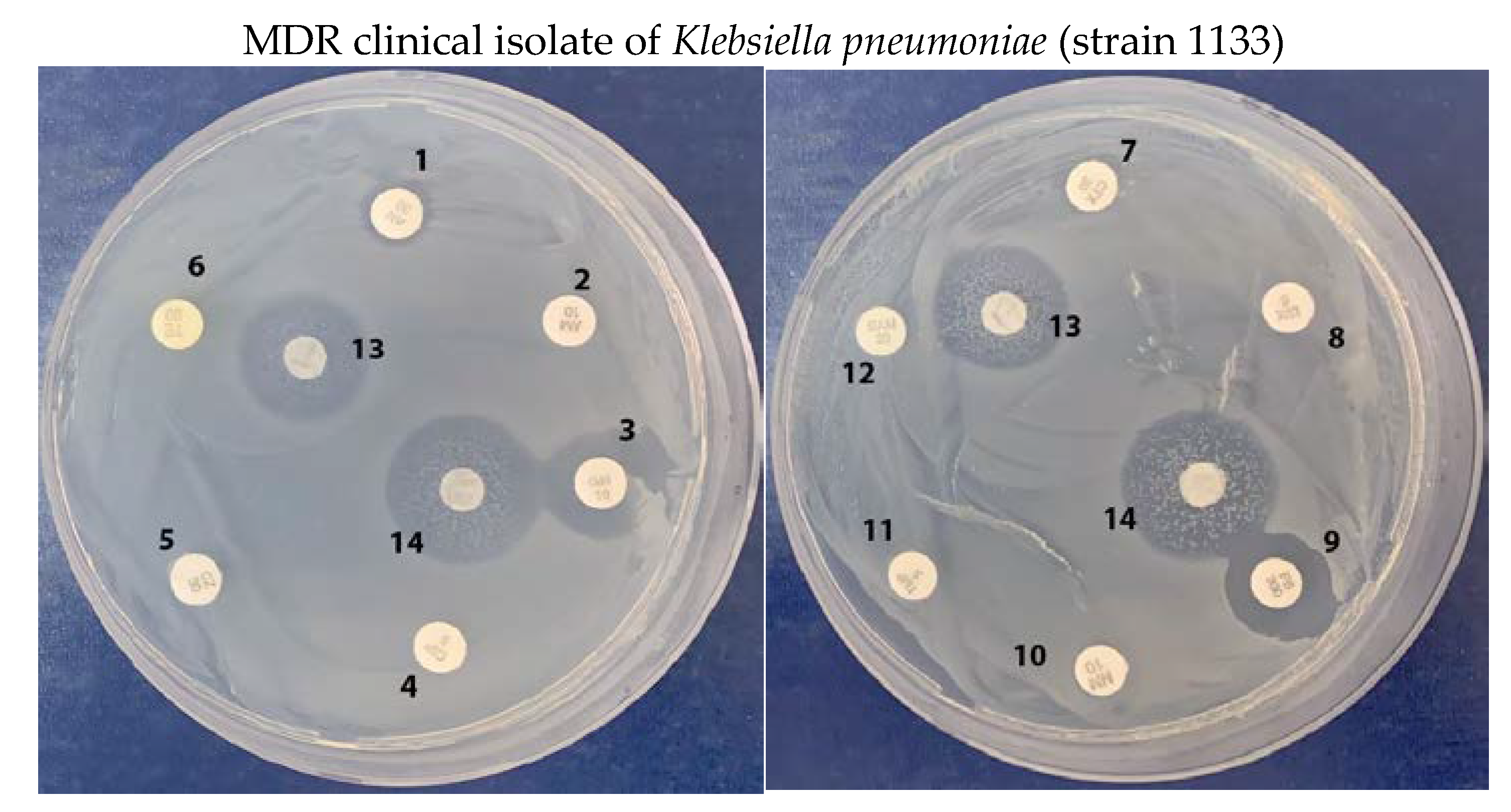
2.4. Growth Inhibition of Multidrug-Resistant Klebsiella pneumoniae Clinical Isolates
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Synthesis of L-Leucyl-L-Phosphinothricin
4.3. The Microdilution Method to Determine the Antimicrobial Activity of Tested Compounds against Escherichia coli
4.4. The Agar Diffusion Method to Analyze Antimicrobial Activity of Tested Compounds against Klebsiella pneumoniae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- 60-Minute Diagnostic Tests Tackling Antibiotic Resistance to Be Eligible for £8m Longitude Prize. Available online: https://longitudeprize.org/press-release/60-minute-diagnostic-tests-tackling-antibiotic-resistance-to-be-eligible-for-8m-longitude-prize/ (accessed on 23 November 2022).
- O’Neill, J.; The Review on Antimicrobial Resistance. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Available online: http://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf (accessed on 23 November 2022).
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Publishes List of Bacteria for which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed/ (accessed on 23 November 2022).
- WHO Regional Office for Europe; European Centre for Disease Prevention and Control. Antimicrobial Resistance Surveillance in Europe 2022—2020 Data; WHO Regional Office for Europe: Copenhagen, Denmark, 2022. [Google Scholar]
- Jeon, K.; Jeong, S.; Lee, N.; Park, M.-J.; Song, W.; Kim, H.-S.; Kim, H.S.; Kim, J.-S. Impact of COVID-19 on antimicrobial consumption and spread of multidrug-resistance in bacterial infections. Antibiotics 2022, 11, 535. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Garrity, G.M. Bergey’s Manual of Systematic Bacteriology, V. 2: The Proteobacteria, Part B: The Gammaproteobacteria; Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; ISBN 978-0-387-24144-9. [Google Scholar]
- Podschun, R.; Pietsch, S.; Höller, C.; Ullmann, U. Incidence of Klebsiella species in surface waters and their expression of virulence factors. Appl. Environ. Microbiol. 2001, 67, 3325–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asri, N.A.M.; Ahmad, S.; Mohamud, R.; Hanafi, N.M.; Zaidi, N.F.M.; Irekeola, A.A.; Shueb, R.H.; Yee, L.C.; Noor, N.M.; Mustafa, F.H.; et al. Global Prevalence of Nosocomial Multidrug-Resistant Klebsiella pneumoniae: A Systematic Review and Meta-Analysis. Antibiotics 2021, 10, 1508. [Google Scholar] [CrossRef]
- Galani, I.; Karaiskos, I.; Giamarellou, H. Multidrug-resistant Klebsiella pneumoniae: Mechanisms of resistance including updated data for novel β-lactam-β-lactamase inhibitor combinations. Expert Rev. Anti-Infect. Ther. 2021, 19, 1457–1468. [Google Scholar] [CrossRef]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. BioMed Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [Green Version]
- Ashurst, J.V.; Dawson, A. Klebsiella pneumonia; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Gill, H.S.; Eisenberg, D. The crystal structure of phosphinothricin in the active site of glutamine synthetase illuminates the mechanism of enzymatic inhibition. Biochemistry 2001, 40, 1903–1912. [Google Scholar] [CrossRef]
- Abell, L.M.; Villafranca, J.J. Investigation of the mechanism of phosphinothricin inactivation of Escherichia coli glutamine synthetase using rapid quench kinetic technique. Biochemistry 1991, 30, 6135–6141. [Google Scholar] [CrossRef]
- Mowbray, S.L.; Kathiravan, M.K.; Pandey, A.A.; Odell, L.R. Inhibition of glutamine synthetase: A potential drug target in Mycobacterium tuberculosis. Molecules 2014, 19, 13161–13176. [Google Scholar] [CrossRef]
- Kumari, M.; Subbarao, N. Virtual screening to identify novel potential inhibitors for glutamine synthetase of Mycobacterium tuberculosis. J. Biomol. Struct. Dyn. 2020, 38, 5062–5080. [Google Scholar] [CrossRef]
- Nordqvist, A.; Nilsson, M.T.; Röttger, S.; Odell, L.R.; Krajewski, W.W.; Evalena Andersson, C.; Larhed, M.; Mowbray, S.L.; Karlén, A. Evaluation of the amino acid binding site of Mycobacterium tuberculosis glutamine synthetase for drug discovery. Bioorg. Med. Chem. 2008, 16, 5501–5513. [Google Scholar] [CrossRef] [Green Version]
- Bayer, E.; Gugel, K.H.; Hägele, K.; Hagenmaier, H.; Jessipow, S.; König, W.A.; Zähner, H. Stoffwechselprodukte von mikroorganismen. 98 Mitteilung. Phosphinothricin und phosphinothricyl-alanyl-alanin. Helv. Chim. Acta 1972, 55, 224–239. [Google Scholar] [CrossRef] [PubMed]
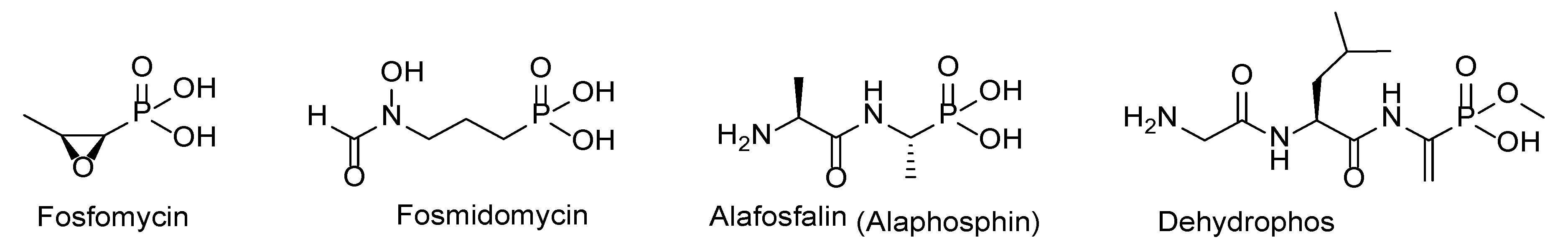
- Kafarski, P. Phosphonopeptides containing free phosphonic groups: Recent advances. RSC Adv. 2020, 10, 25898–25910. [Google Scholar] [CrossRef] [PubMed]
- Hörömpöli, D.; Ciglia, C.; Glüsenkamp, K.H.; Haustedt, L.O.; Falkenstein-Paul, H.; Bendas, G.; Berscheid, A.; Brötz-Oesterhelt, H. The antibiotic negamycin crosses the bacterial cytoplasmic membrane by multiple routes. Antimicrob. Agents Chemother. 2021, 65, e00986-20. [Google Scholar] [CrossRef]
- Allen, J.G.; Atherton, F.R.; Hall, M.J.; Hassall, C.H.; Holmes, S.W.; Lambert, R.W.; Nisbet, L.J.; Ringrose, P.S. Phosphonopeptides, a new class of synthetic antibacterial agents. Nature 1978, 272, 56–58. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Kim, Y.J.; Yoshizawa, M.; Takenaka, S.; Murakami, S.; Aoki, K. Ammonia assimilation in Klebsiella pneumoniae F-5-2 that can utilize ammonium and nitrate ions simultaneously: Purification and characterization of glutamate dehydrogenase and glutamine synthetase. J. Biosci. Bioeng. 2002, 93, 584–588. [Google Scholar] [CrossRef]
- Horsman, G.P.; Zechel, D.L. Phosphonate biochemistry. Chem. Rev. 2017, 117, 5704–5783. [Google Scholar] [CrossRef]
- Metcalf, W.W.; van der Donk, W.A. Biosynthesis of phosphonic and phosphinic acid natural products. Annu. Rev. Biochem. 2009, 78, 65–94. [Google Scholar] [CrossRef] [Green Version]
- Falagas, M.E.; Vouloumanou, K.; Samonis, G.; Vardakas, K.Z. Fosfomycin. Clin. Microbiol. Rev. 2016, 29, 321–347. [Google Scholar] [CrossRef]
- Jawaid, S.; Seidle, H.; Zhou, W.; Abdirahman, H.; Abadeer, M.; Hix, J.H.; van Hoek, M.L.; Couch, R.D. Kinetic characterization and phosphoregulation of the Francisella tularensis 1-deoxy-D-xylulose 5-phosphate reductoisomerase (MEP synthase). PLoS ONE 2009, 4, e8288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pines, G.; Oh, E.J.; Bassalo, M.C.; Choudhury, A.; Garst, A.D.; Fankhauser, R.G.; Eckert, C.A.; Gill, R.T. Genomic Deoxyxylulose Phosphate Reductoisomerase (DXR) mutations conferring resistance to the antimalarial drug Fosmidomycin in E. coli. ACS Synth. Biol. 2018, 7, 2824–2832. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Doroghazi, J.R.; Janga, S.C.; Zhang, J.K.; Circello, B.; Grifiin, B.M.; Labeda, D.P.; Metcalf, W.W. Diversity and abundance of phosphonate biosynthetic genes in nature. Proc. Natl. Acad. Sci. USA 2013, 110, 20759–20764. [Google Scholar] [CrossRef] [Green Version]
- Ju, K.-S.; Gao, J.; Doroghazi, J.R.; Wang, K.-K.A.; Thibodeaux, C.J.; Li, S.; Metzger, E.; Fudala, J.; Su, J.; Zhang, J.K.; et al. Discovery of phosphonic acid natural products by mining the genomes of 10,000 actinomycetes. Proc. Natl. Acad. Sci. USA 2015, 112, 12175–12180. [Google Scholar] [CrossRef] [Green Version]
- Nowak, M.G.; Skwarecki, A.S.; Milewska, M.J. Amino acid based antimicrobial agents—Synthesis and properties. ChemMedChem 2021, 16, 3513–3544. [Google Scholar] [CrossRef] [PubMed]
- Wicke, D.; Schulz, L.M.; Lentes, S.; Scholz, P.; Poehlein, A.; Gibhardt, J.; Daniel, R.; Ischebeck, T.; Commichau, F.M. Identification of the first glyphosate transporter by genomic adaptation. Environ. Microbiol. 2019, 21, 1287–1305. [Google Scholar] [CrossRef] [PubMed]
- Circello, B.T.; Miller, C.G.; Lee, J.-H.; van der Donk, W.A.; Metcalf, W.W. The antibiotic dehydrophos is converted to a toxic pyruvate analog by peptide bond cleavage in Salmonella enterica. Antimicrob. Agents Chemother. 2011, 55, 3357–3362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunik, V.I.; Artiukhov, A.; Kazantsev, A.; Goncalves, R.; Daloso, D.; Oppermann, H.; Kulakovskaya, E.; Lukashev, N.; Fernie, A.; Brand, M.; et al. Specific inhibition by synthetic analogs of pyruvate reveals that the pyruvate dehydrogenase reaction is essential for metabolism and viability of glioblastoma cells. Oncotarget 2015, 6, 40036–40052. [Google Scholar] [CrossRef] [Green Version]
- Blodgett, J.A.; Thomas, P.M.; Li, G.; Velasquez, J.E.; van der Donk, W.A.; Kelleher, N.L.; Metcalf, W.W. Unusual transformations in the biosynthesis of the antibiotic phosphinothricin tripeptide. Nat. Chem. Biol. 2007, 3, 480–485. [Google Scholar] [CrossRef] [Green Version]
- Ujváry, I. Pest Control Agents from Natural Products. In Hayes’ Handbook of Pesticide Toxicology, 3rd ed.; Krieger, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 1, pp. 183–188. [Google Scholar]
- Omura, S.; Murata, M.; Hanaki, H.; Hinotozawa, K.; Oiwa, R.; Tanaka, H. Phosalacine, a new herbicidal antibiotic containing phosphinothricin. Fermentation, isolation, biological activity and mechanism of action. J. Antibiot. 1984, 37, 829–835. [Google Scholar] [CrossRef]
- Anderson, G.W.; Zimmerman, J.E.; Callahan, F.M. N-Hydroxysuccinimide esters in peptide synthesis. J. Am. Chem. Soc. 1963, 85, 3039. [Google Scholar] [CrossRef]
- De Biase, D.; Cappadocio, F.; Pennacchietti, E.; Giovannercole, F.; Coluccia, A.; Vepsäläinen, J.; Khomutov, A. Enzymatic kinetic resolution of desmethylphosphinothricin indicates that phosphinic group is a bioisostere of carboxyl group. Commun. Chem. 2020, 3, 121. [Google Scholar] [CrossRef]
- Antimicrobial Susceptibility Testing: EUCAST Disk Diffusion Method. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Disk_test_documents/2021_manuals/Manual_v_9.0_EUCAST_Disk_Test_2021.pdf (accessed on 23 November 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Medium | ||
|---|---|---|---|
| Minimal EG | Mueller–Hinton | ||
| Amino acid | D,L-Phosphinothricin (D,L-PT) | 266–304 | n.d. |
| Dipeptides | L-Ala-L-aminoethylphosphonic acid (Alaphosphin) | 0.001–0.01 * | 1000 (MIC50) |
| L-Leu-L-PT | 0.04 | 1600 | |
| Tripeptide | Bialaphos (L-PT-L-Ala-L-Ala) | <0.001 | 500 (MIC87) |
| PT Derivatives (μg/Disk) | Strain 1161 | Strain 1158 | Strain 1133 | ATCC 13883 |
|---|---|---|---|---|
| Diameter of Inhibition Zones, mm | ||||
| Bialaphos, 0.01 | 0/0 | 0/0 | 0/0 | 0/0 |
| Bialaphos, 0.1 | 0/0 | 0/0 | 0/0 | 9/8 ** |
| Bialaphos, 0.5 | 0/0 | 10/9 | 13/15 | n.t. |
| Bialaphos, 1.0 | 0/0 | 8/7 | 13/13 | 15/14 *** |
| Bialaphos, 5.0 | 14/13 | 18/15 | 16/17 | 20/20 |
| Bialaphos, 10.0 | 18/17 | 21/19 | 21/21 | 25/27 |
| L-Leu-L-PT, 0.01 | 0/0 | 0/0 | 0/0 | 0/0 |
| L-Leu-L-PT, 0.1 | 0/0 | 0/0 | 11/12 | 8/7 ** |
| L-Leu-L-PT, 0.5 | 10/11 | 8/9 | 12/14 | n.t. |
| L-Leu-L-PT, 1.0 | 13/13 | 13/13 | 16/16 | 12/10 *** |
| L-Leu-L-PT, 5.0 | 16/20 | 20/20 | 20/22 | 14/12 |
| L-Leu-L-PT, 10.0 | 18/21 | 23/23 | 25/25 | 19/20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demiankova, M.V.; Giovannercole, F.; Khomutov, M.A.; Salikhov, A.I.; Onillon, L.; Valuev-Elliston, V.T.; Vasilieva, B.F.; Khurs, E.N.; Gabrielyan, N.I.; Kochetkov, S.N.; et al. Antibacterial Activity of Peptide Derivatives of Phosphinothricin against Multidrug-Resistant Klebsiella pneumoniae. Molecules 2023, 28, 1234. https://doi.org/10.3390/molecules28031234
Demiankova MV, Giovannercole F, Khomutov MA, Salikhov AI, Onillon L, Valuev-Elliston VT, Vasilieva BF, Khurs EN, Gabrielyan NI, Kochetkov SN, et al. Antibacterial Activity of Peptide Derivatives of Phosphinothricin against Multidrug-Resistant Klebsiella pneumoniae. Molecules. 2023; 28(3):1234. https://doi.org/10.3390/molecules28031234
Chicago/Turabian StyleDemiankova, Marija V., Fabio Giovannercole, Maxim A. Khomutov, Arthur I. Salikhov, Laura Onillon, Vladimir T. Valuev-Elliston, Byazilya F. Vasilieva, Elena N. Khurs, Nina I. Gabrielyan, Sergey N. Kochetkov, and et al. 2023. "Antibacterial Activity of Peptide Derivatives of Phosphinothricin against Multidrug-Resistant Klebsiella pneumoniae" Molecules 28, no. 3: 1234. https://doi.org/10.3390/molecules28031234
APA StyleDemiankova, M. V., Giovannercole, F., Khomutov, M. A., Salikhov, A. I., Onillon, L., Valuev-Elliston, V. T., Vasilieva, B. F., Khurs, E. N., Gabrielyan, N. I., Kochetkov, S. N., Efremenkova, O. V., De Biase, D., & Khomutov, A. R. (2023). Antibacterial Activity of Peptide Derivatives of Phosphinothricin against Multidrug-Resistant Klebsiella pneumoniae. Molecules, 28(3), 1234. https://doi.org/10.3390/molecules28031234