
Impact of Starch Binding Domain Fusion on Activities and Starch Product Structure of 4-α-Glucanotransferase
 ,
,  , and
, and
Abstract
:1. Introduction
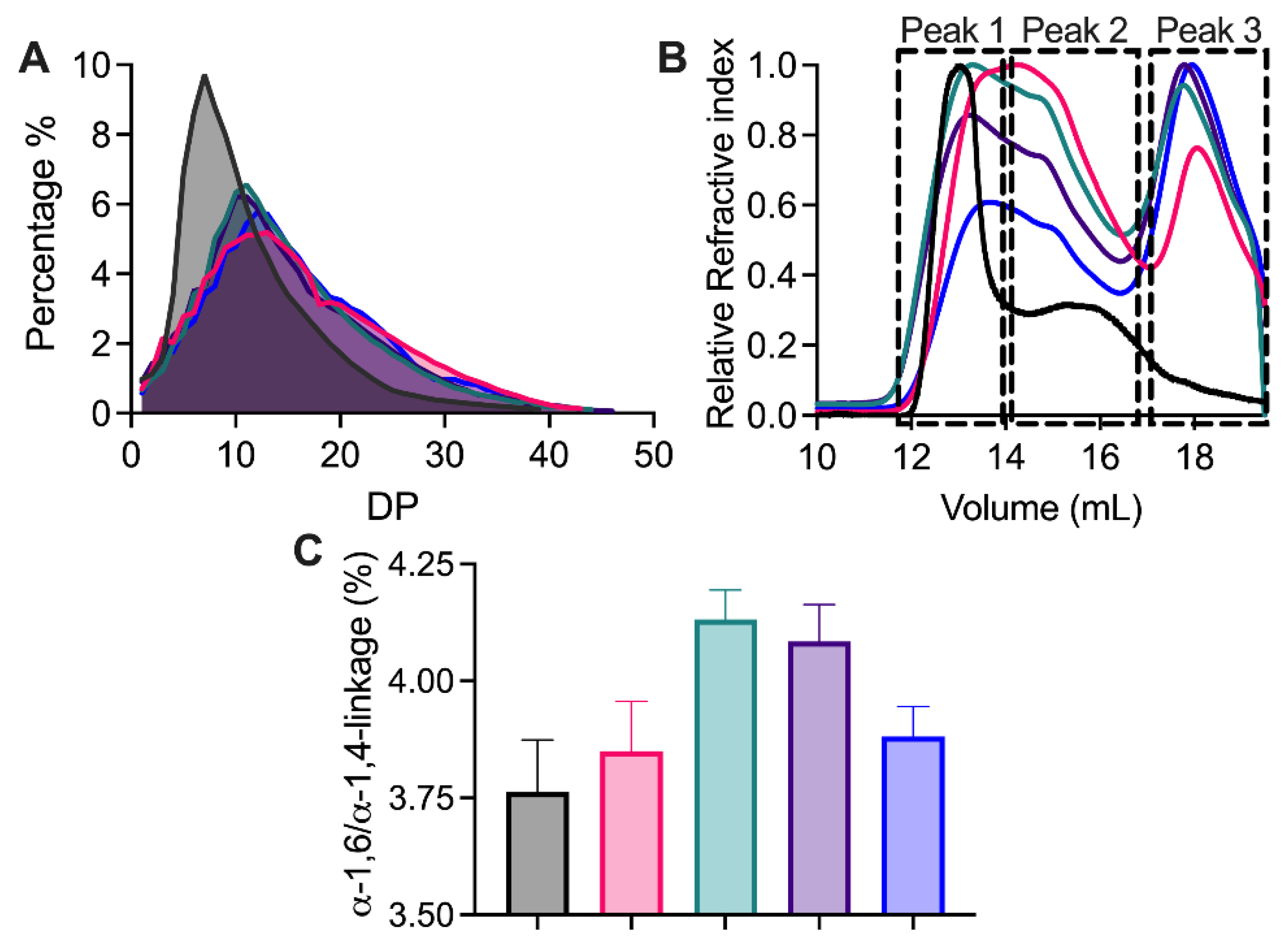
2. Results and Discussion
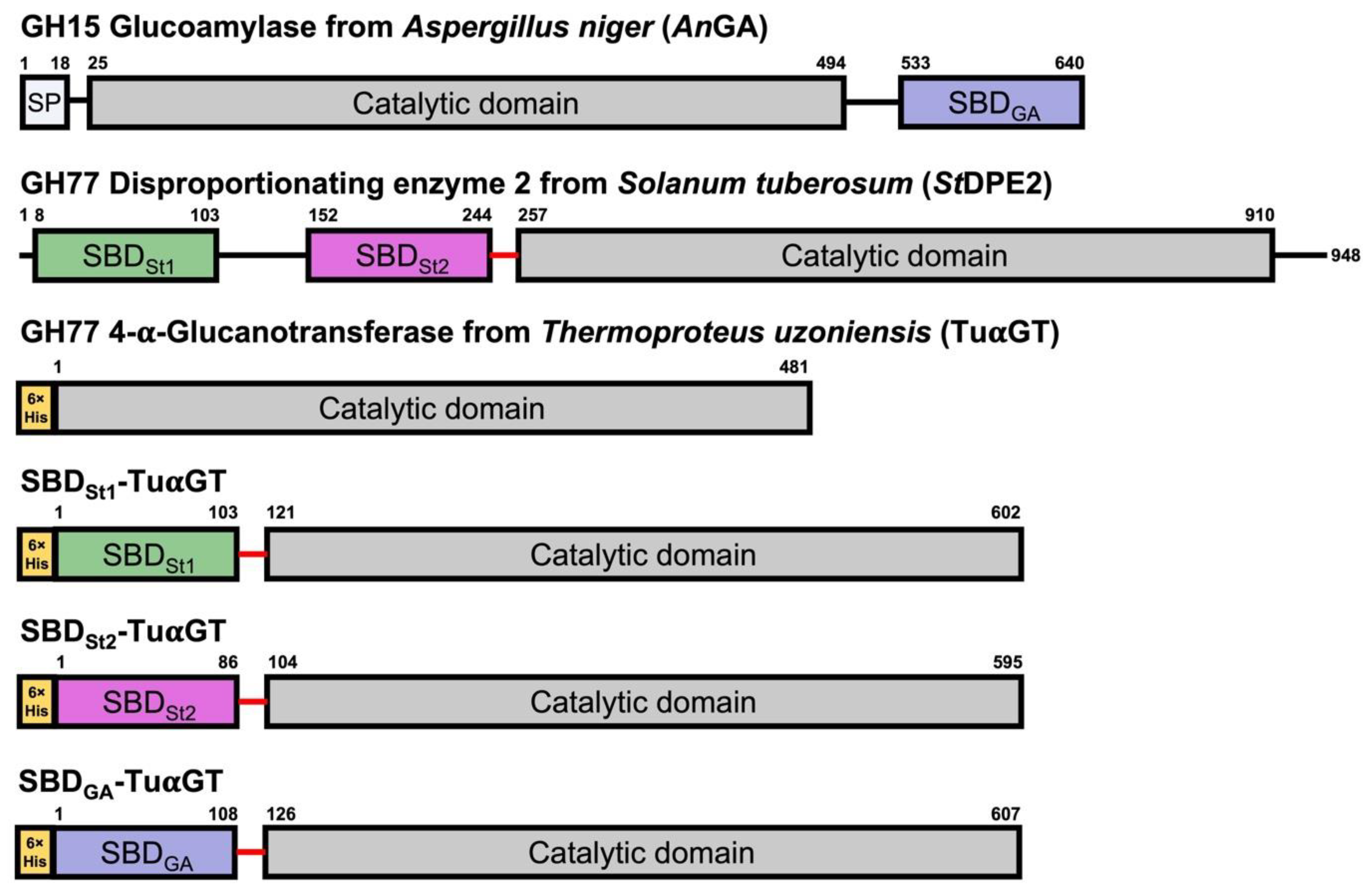
2.1. 4-α-Glucanotransferase SBD Fusions
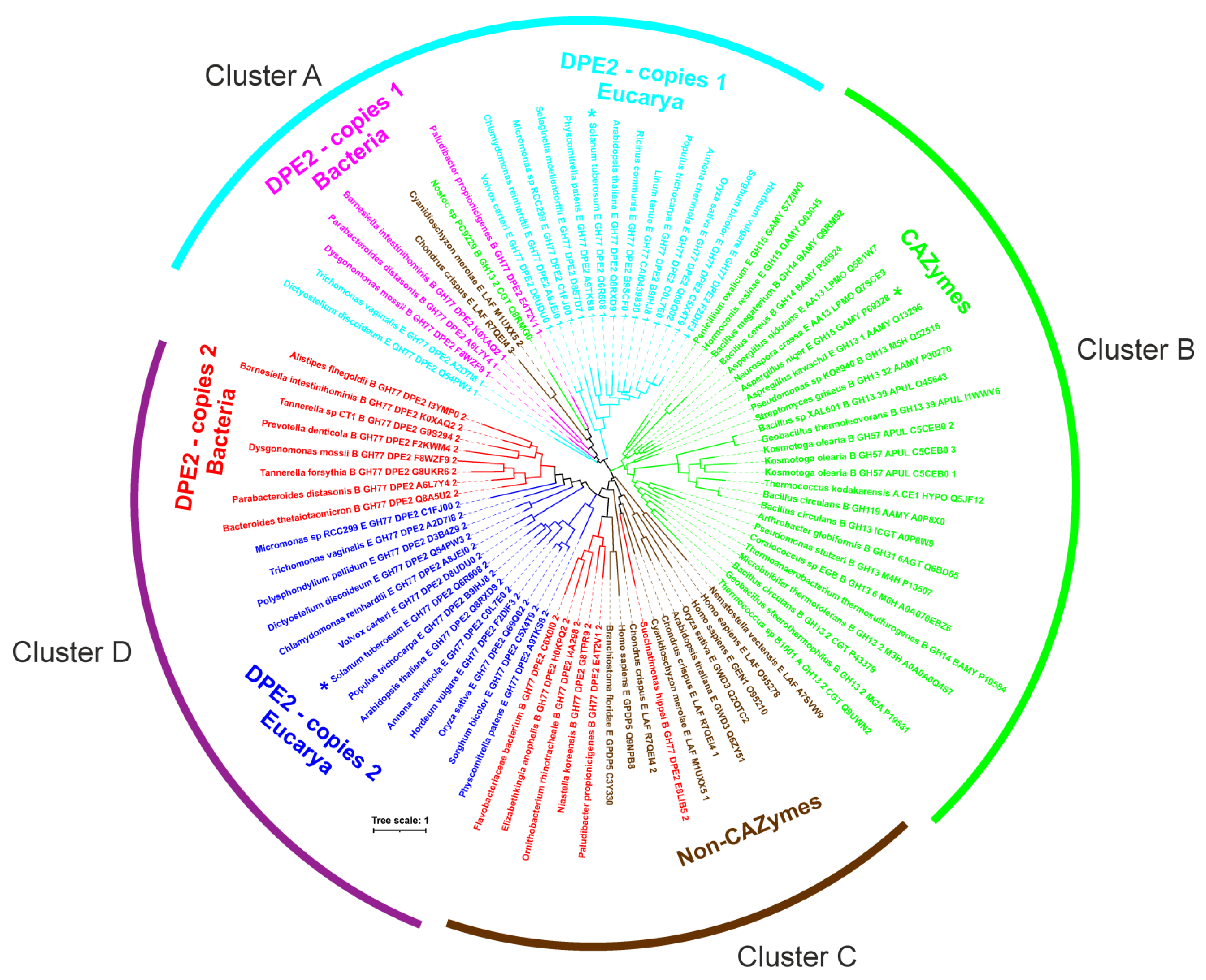
2.2. Bioinformatics Analysis
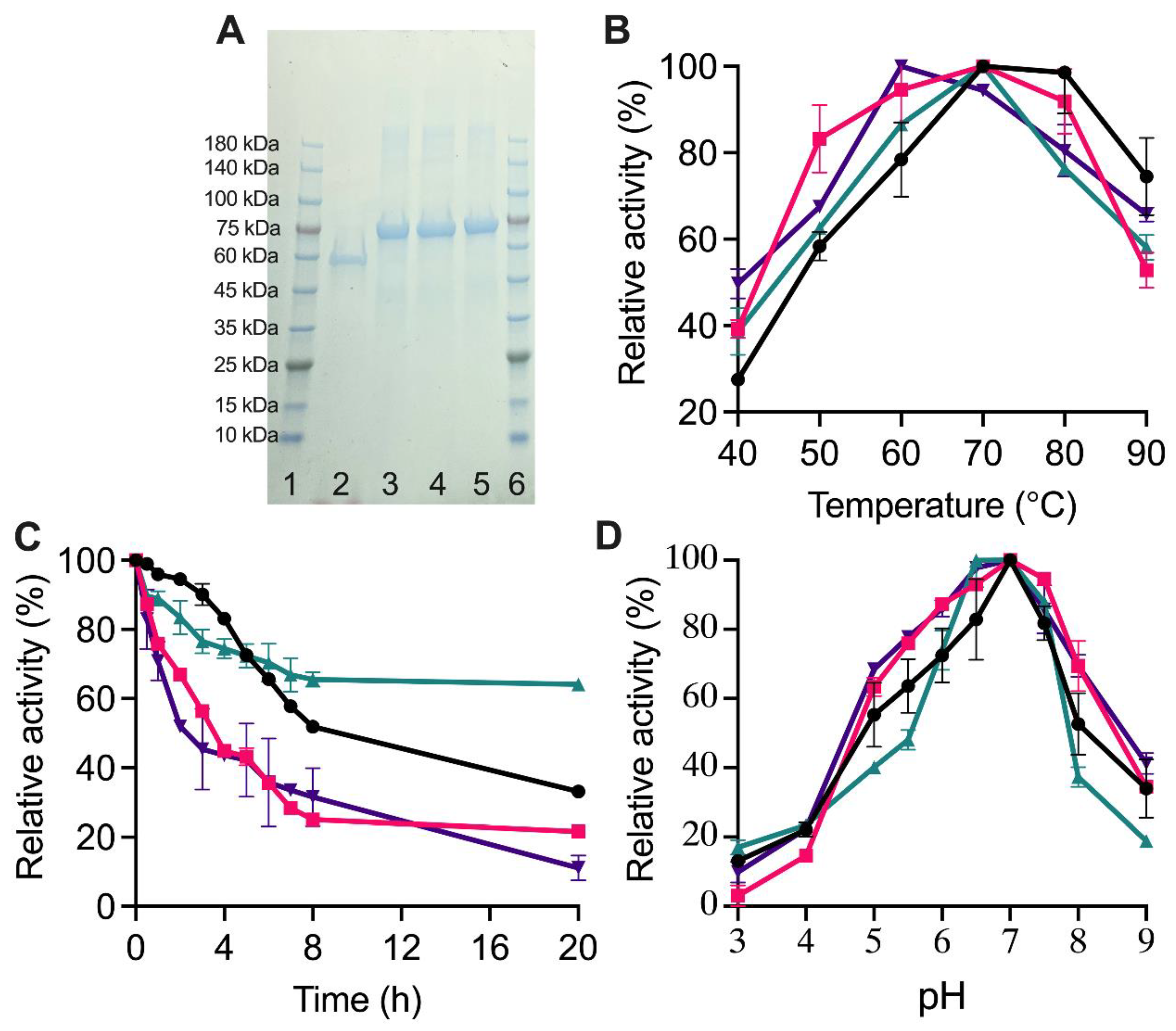
2.3. Biochemical Properties of TuαGT and SBD-TuαGT Fusions
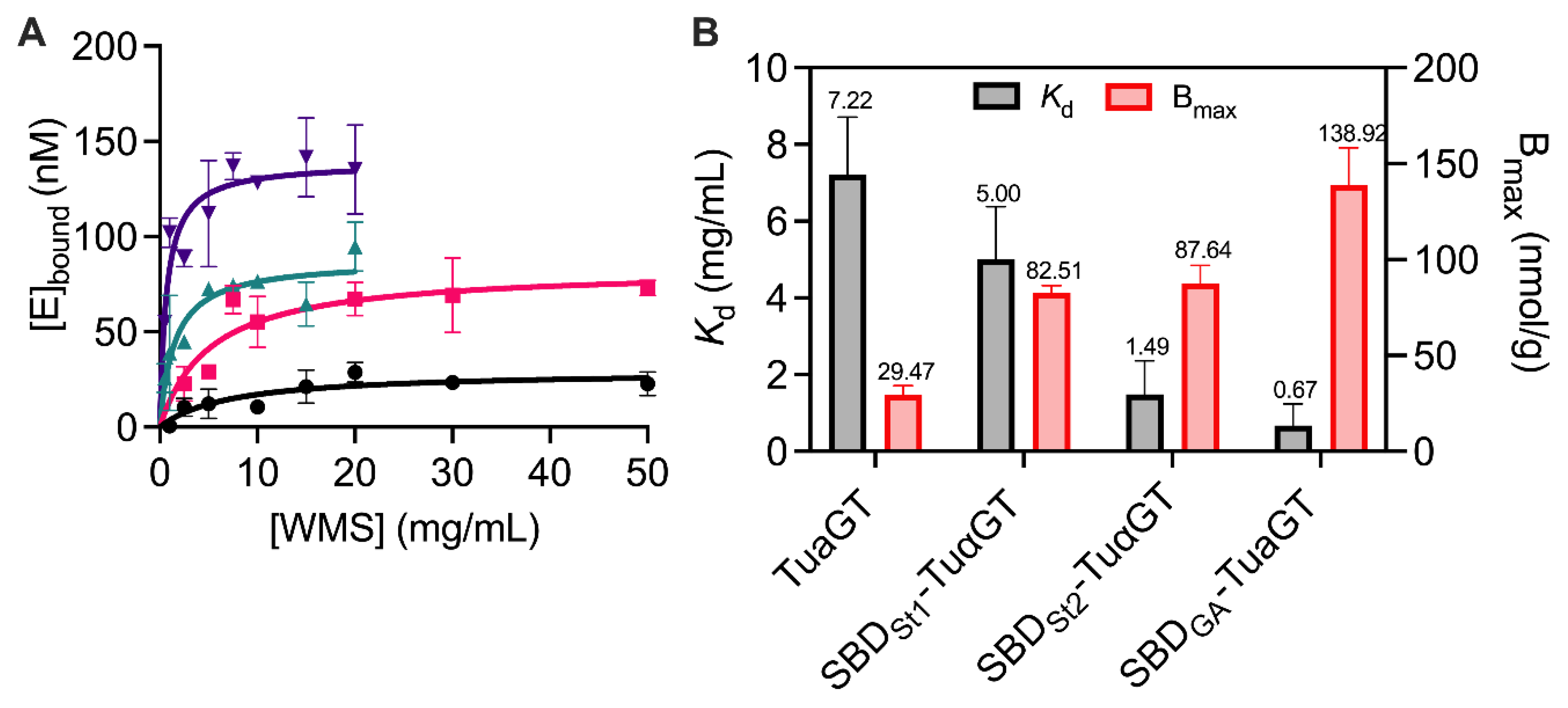
2.4. Adsorption and Enzyme Kinetic Parameters
2.5. Hydrolysis and Cyclization Activities on Different Substrates
2.6. Structure Analysis of Modified NMS
3. Material and Methods
3.1. Materials
3.2. Bioinformatics Analysis of CBM20
3.3. Construction of TuαGT and SBD-TuαGT Fusions
3.4. Production of TuαGT and SBD-TuαGT Fusions
3.5. Purification of TuαGT and SBD-TuαGT Fusions
3.6. Enzyme Activity Assays
3.6.1. Total Activity
3.6.2. Disproportionation
3.6.3. Hydrolysis
3.6.4. Cyclization
3.7. Effect of pH and Temperature on Activity
3.8. Kinetic Parameters
3.9. Adsorption to Starch Granules
3.10. Preparation of Modified Maize Starch (MMS)
3.11. Molecular Weight Distribution
3.12. Chain Length Distribution
3.13. 1H-NMR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kim, H.J.; Kim, Y.H.; Cha, H.; Kim, Y.W.; Kim, T.J.; Kim, Y.R.; Park, K.H. The action mode of Thermus aquaticus YT-1 4-α-glucanotransferase and its chimeric enzymes introduced with starch-binding domain on amylose and amylopectin. Carbohydr. Polym. 2007, 67, 164–173. [Google Scholar] [CrossRef]
- Suksiri, P.; Ismail, A.; Sirirattanachatchawan, C.; Wangpaiboon, K.; Muangsin, N.; Tananuwong, K.; Krusong, K. Enhancement of large ring cyclodextrin production using pretreated starch by glycogen debranching enzyme from Corynebacterium glutamicum. Int. J. Biol. Macromol. 2021, 193, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Y.; Wu, J.; Jin, Z.; Dijkhuizen, L.; Abou Hachem, M.; Bai, Y. Thermoproteus uzoniensis 4- α -glucanotransferase catalyzed production of a thermo-reversible potato starch gel with superior rheological properties and freeze-thaw stability. Food Hydrocoll. 2023, 134, 108026. [Google Scholar] [CrossRef]
- Janecek, S.; Marecek, F.; MacGregor, E.A.; Svensson, B. Starch-binding domains as CBM families—History, occurrence, structure, function and evolution. Biotechnol. Adv. 2019, 37, 107451. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Belshaw, N.J.; Williamson, M.P. O-glycosylation in Aspergillus glucoamylase. Conformation and role in binding. Biochem. J. 1992, 282, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Paldi, T.; Levy, I.; Shoseyov, O. Glucoamylase starch-binding domain of Aspergillus niger B1: Molecular cloning and functional characterization. Biochem. J. 2003, 372, 905–910. [Google Scholar] [CrossRef]
- Sorimachi, K.; Le Gal-Coëffet, M.-F.; Williamson, G.; Archer, D.B.; Williamson, M.P. Solution structure of the granular starch binding domain of Aspergillus niger glucoamylase bound to β-cyclodextrin. Structure 1997, 5, 647–661. [Google Scholar] [CrossRef] [Green Version]
- Giardina, T.; Gunning, A.P.; Juge, N.; Faulds, C.B.; Furniss, C.S.M.; Svensson, B.; Morris, V.J.; Williamson, G. Both binding sites of the starch-binding domain of Aspergillus niger glucoamylase are essential for inducing a conformational change in amylose. J. Mol. Biol. 2001, 313, 1149–1159. [Google Scholar] [CrossRef]
- Southall, S.M.; Simpson, P.J.; Gilbert, H.J.; Williamson, G.; Williamson, M.P. The starch-binding domain from glucoamylase disrupts the structure of starch. FEBS Lett. 1999, 447, 58–60. [Google Scholar] [CrossRef]
- Armenta, S.; Moreno-Mendieta, S.; Sánchez-Cuapio, Z.; Sánchez, S.; Rodríguez-Sanoja, R. Advances in molecular engineering of carbohydrate-binding modules. Proteins 2017, 85, 1602–1617. [Google Scholar] [CrossRef] [PubMed]
- Penninga, D.; van der Veen, B.A.; Knegtel, R.M.A.; van Hijum, S.A.F.T.; Rozeboom, H.J.; Kalk, K.H.; Dijkstra, B.W.; Dijkhuizen, L. The raw starch binding domain of cyclodextrin glycosyltransferase from Bacillus circulans strain 251. J. Biol. Chem. 1996, 271, 32777–32784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, X.; Ji, H.; Zheng, D.; Jin, Z.; Bai, Y.; Svensson, B. Thermophilic 4-α-glucanotransferase from Thermoproteus uzoniensis retards the long-term retrogradation but maintains the short-term gelation strength of tapioca starch. J. Agric. Food Chem. 2020, 68, 5658–5667. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.R.; Blennow, A.; Burhenne, K.; Kossmann, J. Repression of a novel isoform of disproportionating enzyme (StDPE2) in potato leads to inhibition of starch degradation in leaves but not tubers stored at low temperature. Plant Physiol. 2004, 134, 1347–1354. [Google Scholar] [CrossRef] [Green Version]
- Svensson, B.; Pedersen, T.; Svendsen, I.; Sakai, T.; Ottesen, M. Characterization of two forms of glucoamylase from Aspergillus niger. Carlsb. Res. Commun. 1982, 47, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Kuchtova, A.; Janecek, S. In silico analysis of family GH77 with focus on amylomaltases from borreliae and disproportionating enzymes DPE2 from plants and bacteria. Biochim. Biophys. Acta 2015, 1854, 1260–1268. [Google Scholar] [CrossRef]
- Janecek, S.; Svensson, B.; MacGregor, E.A. Structural and evolutionary aspects of two families of non-catalytic domains present in starch and glycogen binding proteins from microbes, plants and animals. Enzyme Microb. Technol. 2011, 49, 429–440. [Google Scholar] [CrossRef]
- Kuchtova, A.; Gentry, M.S.; Janecek, S. The unique evolution of the carbohydrate-binding module CBM20 in laforin. FEBS Lett. 2018, 592, 586–598. [Google Scholar] [CrossRef] [Green Version]
- Knegtel, R.M.A.; Strokopytov, B.; Penninga, D.; Faber, O.G.; Rozeboom, H.J.; Kalk, K.H.; Dijkhuizen, L.; Dijkstra, B.W. Crystallographic studies of the interaction of cyclodextrin glycosyltransferase from Bacillus circulans strain 251 with natural substrates and products. J. Biol. Chem. 1995, 270, 29256–29264. [Google Scholar] [CrossRef] [Green Version]
- Janecek, S.; Svensson, B.; MacGregor, E.A. Relation between domain evolution, specificity, and taxonomy of the α-amylase family members containing a C-terminal starch-binding domain. Eur. J. Biochem. 2003, 270, 635–645. [Google Scholar] [CrossRef]
- Tang, C.D.; Li, J.F.; Wei, X.H.; Min, R.; Gao, S.J.; Wang, J.Q.; Yin, X.; Wu, M.C. Fusing a carbohydrate-binding module into the Aspergillus usamii β-mannanase to improve its thermostability and cellulose-binding capacity by in silico design. PLoS ONE 2013, 8, e64766. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cui, G.Z.; Song, X.F.; Feng, Y.; Cui, Q. Efficiency and stability enhancement of cis-epoxysuccinic acid hydrolase by fusion with a carbohydrate binding module and immobilization onto cellulose. Appl. Biochem. Biotechnol. 2012, 168, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Juge, N.; Nøhr, J.; Le Gal-Coëffet, M.F.; Kramhøft, B.; Furniss, C.S.M.; Planchot, V.; Archer, D.B.; Williamson, G.; Svensson, B. The activity of barley α-amylase on starch granules is enhanced by fusion of a starch binding domain from Aspergillus niger glucoamylase. Biochim. Biophys. Acta 2006, 1764, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Sauer, J.; Sigurskjold, B.W.; Christensen, U.; Frandsen, T.P.; Mirgorodskaya, E.; Harrison, M.; Roepstorff, P.; Svensson, B. Glucoamylase: Structure/function relationships, and protein engineering. Biochim. Biophys. Acta 2000, 1543, 275–293. [Google Scholar] [CrossRef]
- Rho, S.J.; Mun, S.; Hong, J.S.; Kim, Y.L.; Do, H.V.; Kim, Y.W.; Han, S.I.; Kim, Y.R. Physicochemical interactions of cycloamylose with phenolic compounds. Carbohydr. Polym. 2017, 174, 980–989. [Google Scholar] [CrossRef]
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Pruitt, K.D.; Schoch, C.L.; Sherry, S.T.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res. 2021, 49, D92–D96. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Burley, S.K.; Bhikadiya, C.; Bi, C.; Bittrich, S.; Chen, L.; Crichlow, G.V.; Christie, C.H.; Dalenberg, K.; Di Costanzo, L.; Duarte, J.M.; et al. RCSB Protein Data Bank: Powerful new tools for exploring 3D structures of biological macromolecules for basic and applied research and education in fundamental biology, biomedicine, biotechnology, bioengineering and energy sciences. Nucleic Acids Res. 2021, 49, D437–D451. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Shatsky, M.; Nussinov, R.; Wolfson, H.J. A method for simultaneous alignment of multiple protein structures. Proteins 2004, 56, 143–156. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Bai, Y.; van der Kaaij, R.M.; Leemhuis, H.; Pijning, T.; van Leeuwen, S.S.; Jin, Z.; Dijkhuizen, L. Biochemical characterization of the Lactobacillus reuteri glycoside hydrolase family 70 GTFB type of 4,6-α-glucanotransferase enzymes that synthesize soluble dietary starch fibers. Appl. Environ. Microbiol. 2015, 81, 7223–7232. [Google Scholar] [CrossRef] [Green Version]
- Huggett, A.S.G.; Nixon, D.A. Enzymic determination of blood glucose. Biochem. J. 1957, 66, 12. [Google Scholar]
- Ballschmiter, M.; Futterer, O.; Liebl, W. Identification and characterization of a novel intracellular alkaline α-amylase from the hyperthermophilic bacterium Thermotoga maritima MSB8. Appl. Environ. Microbiol. 2006, 72, 2206–2211. [Google Scholar] [CrossRef] [Green Version]
- Schiano-di-Cola, C.; Røjel, N.; Jensen, K.; Kari, J.; Sørensen, T.H.; Borch, K.; Westh, P. Systematic deletions in the cellobiohydrolase (CBH) Cel7a from the fungus Trichoderma reesei reveal flexible loops critical for CBH activity. J. Biol. Chem. 2019, 294, 1807–1815. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.; Minagawa, H.; Terada, Y.; Takaha, T.; Kuriki, T.; Shimada, J.; Kaneko, H. Use of random and saturation mutageneses to improve the properties of Thermus aquaticus amylomaltase for efficient production of cycloamyloses. Appl. Environ. Microbiol. 2005, 71, 5823–5827. [Google Scholar] [CrossRef] [Green Version]
- Brooke, D.; Movahed, N.; Bothner, B. Universal buffers for use in biochemistry and biophysical experiments. AIMS Biophys. 2015, 2, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Møller, M.S.; El Bouaballati, S.; Henrissat, B.; Svensson, B. Functional diversity of three tandem C-terminal carbohydrate-binding modules of a β-mannanase. J. Biol. Chem. 2021, 296, 100638. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Bai, Y.; Liu, Y.; Wang, Y.; Zhan, X.; Long, J.; Chen, L.; Qiu, C.; Jin, Z. Deciphering external chain length and cyclodextrin production with starch catalyzed by cyclodextrin glycosyltransferase. Carbohydr. Polym. 2022, 284, 119156. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.J.; Madsen, M.S.; Zinck, S.S.; Hedberg, C.; Sørensen, O.B.; Svensson, B.; Meyer, A.S. Enzymatic potato starch modification and structure-function analysis of six diverse GH77 4-α-glucanotransferases. Int. J. Biol. Macromol. 2023, 224, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Bai, Y.; Ji, H.; Wang, Y.; Jin, Z.; Svensson, B. Controlling the fine structure of glycogen-like glucan by rational enzymatic synthesis. J. Agric. Food Chem. 2021, 69, 14951–14960. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | B/A/E b | Organism | Family c | Enzyme d | GenBank e | UniProt e | Length f | CBM20_1 g | CBM20_2 g | CBM20_3 g | Insert h |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | E | Annona cherimola | GH77 | DPE2 | ACN50178.1 | C0L7E0 | 953 | 10–119 | 154–268 | 606–750 | |
| 2 | E | Arabidopsis thaliana | GH77 | DPE2 | AAL91204.1 | Q8RXD9 | 955 | 13–122 | 157–270 | 608–752 | |
| 3 | E | Chlamydomonas reinhardtii | GH77 | DPE2 | EDO97689.1 | A8JEI0 | 941 | 1–119 | 155–271 | 631–775 | |
| 4 | E | Dictyostelium discoideum | GH77 | DPE2 | EAL65318.1 | Q54PW3 | 907 | 1–102 | 134–241 | 594–729 | |
| 5 | E | Hordeum vulgare | GH77 | DPE2 | BAJ94874.1 | F2DIF3 | 931 | 1–108 | 143–257 | 595–739 | |
| 6 | E | Linum tenue | GH77 | DPE2 | CAI0439830.1 | --- | 1137 | 10–119 | 499–643 | ||
| 7 | E | Micromonas sp. RCC299 | GH77 | DPE2 | ACO70268.1 | C1FJ00 | 975 | 1–114 | 169–286 | 636–793 | |
| 8 | E | Oryza sativa | GH77 | DPE2 | BAD31425.1 | Q69Q02 | 946 | 7–115 | 150–264 | 602–746 | |
| 9 | E | Physcomitrella patens | GH77 | DPE2 | EDQ55980.1 | A9TKS8 | 1006 | 14–123 | 165–279 | 618–763 | |
| 10 | E | Polysphondylium pallidum | GH77 | DPE2 | EFA84397.1 | D3B4Z9 | 1070 | 167–279 | 627–761 | ||
| 11 | E | Populus trichocarpa | GH77 | DPE2 | EEF04969.1 | B9IHJ8 | 975 | 10–119 | 155–268 | 606–750 | |
| 12 | E | Ricinus communis | GH77 | DPE2 | EEF38704.1 | B9SCF0 | 901 | 10–119 | 533–676 | ||
| 13 | E | Selaginella moellendorffii | GH77 | DPE2 | EFJ19739.1 | D8S7D7 | 930 | 15–128 | 600–740 | ||
| 14 | E | Solanum tuberosum | GH77 | DPE2 | AAR99599.1 | Q6R608 | 948 | 1–112 | 147–259 | 597–741 | |
| 15 | E | Sorghum bicolor | GH77 | DPE2 | EER97686.1 | C5X4T9 | 946 | 6–114 | 149–263 | 601–745 | |
| 16 | E | Trichomonas vaginalis | GH77 | DPE2 | EAY23705.1 | A2D7I8 | 930 | 1–112 | 142–249 | 594–704 | |
| 17 | E | Volvox carteri | GH77 | DPE2 | EFJ42152.1 | D8UDU0 | 995 | 51–178 | 214–329 | 671–786 | |
| 18 | B | Alistipes finegoldii | GH77 | DPE2 | AFL78258.1 | I3YMP0 | 867 | 115–225 | 556–691 | ||
| 19 | B | Bacteroides thetaiotaomicron | GH77 | DPE2 | AAO77253.1 | Q8A5U2 | 893 | 119–235 | 573–714 | ||
| 20 | B | Barnesiella intestinihominis | GH77 | DPE2 | EJZ64889.1 | K0XAQ2 | 893 | 1–97 | 123–239 | 577–718 | |
| 21 | B | Dysgonomonas mossii | GH77 | DPE2 | EGK04046.1 | F8WZF9 | 888 | 1–95 | 119–231 | 571–712 | |
| 22 | B | Elizabethkingia anophelis | GH77 | DPE2 | EHM98897.1 | H0KPQ2 | 885 | 119–225 | 572–711 | ||
| 23 | B | Flavobacteriaceae bacterium | GH77 | DPE2 | ACU06866.1 | C6X0I0 | 884 | 117–226 | 570–709 | ||
| 24 | B | Niastella koreensis | GH77 | DPE2 | AEV98902.1 | G8TPR9 | 895 | 127–241 | 579–720 | ||
| 25 | B | Ornithobacterium rhinotracheale | GH77 | DPE2 | AFL98082.1 | I4A298 | 874 | 109–217 | 563–698 | ||
| 26 | B | Paludibacter propionicigenes | GH77 | DPE2 | ADQ79045.1 | E4T2V1 | 897 | 1–101 | 128–243 | 582–722 | |
| 27 | B | Parabacteroides distasonis | GH77 | DPE2 | ABR41798.1 | A6L7Y4 | 895 | 1–98 | 124–240 | 578–719 | |
| 28 | B | Prevotella denticola | GH77 | DPE2 | AEA21596.1 | F2KWM4 | 897 | 126–233 | 581–722 | ||
| 29 | B | Succinatimonas hippei | GH77 | DPE2 | EFY07743.1 | E8LIB5 | 879 | 112–223 | 562–703 | ||
| 30 | B | Tannerella forsythia | GH77 | DPE2 | AEW22695.1 | G8UKR6 | 881 | 108–223 | 561–701 | ||
| 31 | B | Tannerella sp. CT1 | GH77 | DPE2 | EHL87887.1 | G9S294 | 894 | 124–232 | 577–718 | ||
| 32 | E | Aspregillus kawachii | GH13_1 | AAMY | BAA22993.1 | O13296 | 640 | 533–640 | |||
| 33 | B | Bacillus circulans | GH13_2 | CGT | CAA55023.1 | P43379 | 713 | 608–713 | |||
| 34 | B | Geobacillus stearothermophilus | GH13_2 | MGA | AAA22233.1 | P19531 | 719 | 609–719 | |||
| 35 | B | Nostoc sp. PC9229 | GH13_2 | CGT | AAM16154.1 | Q8RMG0 | 642 | 534–642 | |||
| 36 | B | Microbulbifer thermotolerans | GH13_2 | M3H | AID53183.1 | A0A0A0Q4S7 | 761 | 657–761 | |||
| 37 | A | Thermococcus sp. B1001 | GH13_2 | CGT | BAA88217.1 | Q9UWN2 | 739 | 629–739 | |||
| 38 | B | Coralococcus sp. EGB | GH13_6 | M6H | AII00648.1 | A0A076EBZ6 | 522 | 421–522 | |||
| 39 | B | Streptomyces griseus | GH13_32 | AAMY | CAA40798.1 | P30270 | 566 | 465–566 | |||
| 40 | B | Geobacillus thermoleovorans | GH13_39 | APUL | AFI70750.1 | I1WWV6 | 1655 | 1252–1349 | |||
| 41 | B | Bacillus sp. XAL601 | GH13_39 | APUL | BAA05832.1 | Q45643 | 2032 | 1330–1427 | |||
| 42 | B | Pseudomonas stutzeri | GH13 | M4H | AAA25707.1 | P13507 | 548 | 446–548 | |||
| 43 | B | Pseudomonas sp. KO-8940 | GH13 | M5H | BAA01600.1 | Q52516 | 614 | 509–614 | |||
| 44 | B | Bacillus circulans | GH13 | ICGT | BAF37283.1 | A0P8W9 | 995 | 888–995 | |||
| 45 | B | Bacillus cereus | GH14 | BAMY | BAA75890.1 | P36924 | 551 | 444–551 | |||
| 46 | B | Bacillus megaterium | GH14 | BAMY | CAB61483.1 | Q9RM92 | 545 | 444–545 | |||
| 47 | B | Thermoanaerobacterium thermosulfurogenes | GH14 | BAMY | AAA23204.1 | P19584 | 551 | 448–551 | |||
| 48 | E | Aspergillus niger | GH15 | GAMY | CAA25303.1 | P69328 | 640 | 533–640 | |||
| 49 | E | Hormoconis resinae | GH15 | GAMY | CAA48243.1 | Q03045 | 616 | 501–608 | |||
| 50 | E | Penicillium oxalicum | GH15 | GAMY | EPS30575.1 | S7ZIW0 | 616 | 508–616 | |||
| 51 | B | Arthrobacter globiformis | GH31 | 6AGT | BAD34980.1 | Q6BD65 | 965 | 859–965 | |||
| 52 | B | Kosmotoga_olearia | GH57 | APUL | ACR80150.1 | C5CEB0 | 1354 | 32–136 | 155–258 | 267–372 | |
| 53 | B | Bacillus circulans | GH119 | AAMY | BAF37284.1 | A0P8X0 | 1290 | 1183–1290 | |||
| 54 | E | Aspergillus nidulans | AA13 | LPMO | CBF81866.1 | Q5B1W7 | 385 | 278–385 | |||
| 55 | E | Neurospora crassa | AA13 | LPMO | EAA34371.2 | Q7SCE9 | 385 | 278–385 | |||
| 56 | A | Thermococcus kodakarensis | CE1 | HYPO | BAD84711.1 | Q5JF12 | 449 | 83–188 | |||
| 57 | E | Arabidopsis thaliana | GWD3 | AAC26245.1 | Q6ZY51 | 1196 | 66–166 | ||||
| 58 | E | Oryza sativa | GWD3 | ABA97816.2 | Q2QTC2 | 1206 | 67–168 | ||||
| 59 | E | Branchiostoma floridae | GPDP5 | EEN65442.1 | C3Y330 | 680 | 1–110 | ||||
| 60 | E | Homo sapiens | GPDP5 | BAA92672.1 | Q9NPB8 | 672 | 1–115 | ||||
| 61 | E | Homo sapiens | GEN1 | AAC78827.1 | O95210 | 358 | 258–358 | ||||
| 62 | E | Chondrus crispus | LAF | CDF36183.1 | R7QEI4 | 549 | 1–100 | 167–282 | 285–387 | ||
| 63 | E | Cyanidioschyzon merolae | LAF | BAM83396.1 | M1UXX5 | 532 | 156–267 | 268–374 | |||
| 64 | E | Homo sapiens | LAF | AAG18377.1 | O95278 | 331 | 1–124 | ||||
| 65 | E | Nematostella vectensis | LAF | EDO32135.1 | A7SVW9 | 324 | 1–125 |
| Substrate | Parameter | TuaGT | SBDSt1-TuaGT | SBDSt2-TuaGT | SBDGA-TuaGT |
|---|---|---|---|---|---|
| Maltotriose | Activity (U/mg) | 27.5 ± 0.7 | 3.1 ± 0.5 | 10.3 ± 0.2 | 7.4 ± 0.4 |
| Km (μM) | 1.5 ± 0.1 | 3.5 ± 0.2 | 1.1 ± 0.1 | 1.4 ± 0.1 | |
| kcat (s−1) | 0.04 ± 0.01 | 0.01 ± 0.0002 | 0.01 ± 0.002 | 0.01 ± 0.0005 | |
| kcat/Km (μM−1∙s−1) | 0.03 ± 0.004 | 0.002 ± 0.0003 | 0.01 ± 0.001 | 0.01 ± 0.0004 | |
| Amylose | Activity (U/mg) | 1.3 ± 0.1 | 3.1 ± 0.2 | 2.5 ± 1.1 | 1.6 ± 0.9 |
| Km (mg/mL) | 0.6 ± 0.04 | 1.9 ± 0.1 | 0.6 ± 0.1 | 0.8 ± 0.02 | |
| kcat (s−1) | 2.5 ± 0.3 | 7.0 ± 0.3 | 5.5 ± 0.4 | 3.3 ± 0.2 | |
| kcat/Km (mL∙[mg∙s]−1) | 3.9 ± 0.2 | 3.6 ± 0.03 | 8.6 ± 0.4 | 4.0 ± 0.2 |
| Activity | Substrate | TuaGT | SBDSt1-TuaGT | SBDSt2-TuaGT | SBDGA-TuaGT |
|---|---|---|---|---|---|
| Cyclization | Amylose | 3.2 ± 0.2 | 4.8 ± 0.2 | 3.9 ± 0.3 | 3.3 ± 0.1 |
| Hydrolysis | Amylose | 0.3 ± 0.01 | 0.4 ± 0.01 | 0.4 ± 0.02 | 0.3 ± 0.01 |
| WMS | 0.3 ± 0.02 | 0.5 ± 0.1 | 0.5 ± 0.1 | 0.4 ± 0.02 | |
| NMS | 0.2 ± 0.02 | 0.3 ± 0.03 | 0.3 ± 0.1 | 0.2 ± 0.1 |
| Type of Chain a | NMS | TuaGT | SBDSt1-TuaGT | SBDSt2-TuaGT | SBDGA-TuaGT |
|---|---|---|---|---|---|
| A-chain | 67.2 ± 0.4 | 38.3 ± 0.7 | 35.8 ± 0.9 | 40.2 ± 2.0 | 41.6 ± 0.4 |
| B1-chain | 28.0 ± 0.7 | 46.3 ± 2.0 | 52.0 ± 1.5 | 43.8 ± 3.0 | 45.1 ± 2.0 |
| B2-chain | 4.4 ± 0.2 | 13.5 ± 0.8 | 10.2 ± 0.9 | 11.7 ± 0.9 | 11.8 ± 1.9 |
| B3-chain | 0.6 ± 0.03 | 2.5 ± 0.2 | 2.5 ± 0.3 | 2.6 ± 0.6 | 1.9 ± 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wu, Y.; Christensen, S.J.; Janeček, Š.; Bai, Y.; Møller, M.S.; Svensson, B. Impact of Starch Binding Domain Fusion on Activities and Starch Product Structure of 4-α-Glucanotransferase. Molecules 2023, 28, 1320. https://doi.org/10.3390/molecules28031320
Wang Y, Wu Y, Christensen SJ, Janeček Š, Bai Y, Møller MS, Svensson B. Impact of Starch Binding Domain Fusion on Activities and Starch Product Structure of 4-α-Glucanotransferase. Molecules. 2023; 28(3):1320. https://doi.org/10.3390/molecules28031320
Chicago/Turabian StyleWang, Yu, Yazhen Wu, Stefan Jarl Christensen, Štefan Janeček, Yuxiang Bai, Marie Sofie Møller, and Birte Svensson. 2023. "Impact of Starch Binding Domain Fusion on Activities and Starch Product Structure of 4-α-Glucanotransferase" Molecules 28, no. 3: 1320. https://doi.org/10.3390/molecules28031320
APA StyleWang, Y., Wu, Y., Christensen, S. J., Janeček, Š., Bai, Y., Møller, M. S., & Svensson, B. (2023). Impact of Starch Binding Domain Fusion on Activities and Starch Product Structure of 4-α-Glucanotransferase. Molecules, 28(3), 1320. https://doi.org/10.3390/molecules28031320