

Microorganisms for Ginsenosides Biosynthesis: Recent Progress, Challenges, and Perspectives

Abstract
:1. Introduction
2. Ginsenosides: Classification and Cell Biological Mechanism in Anticancer Activities
3. Endophytes as Novel Biological Source of Ginsenosides
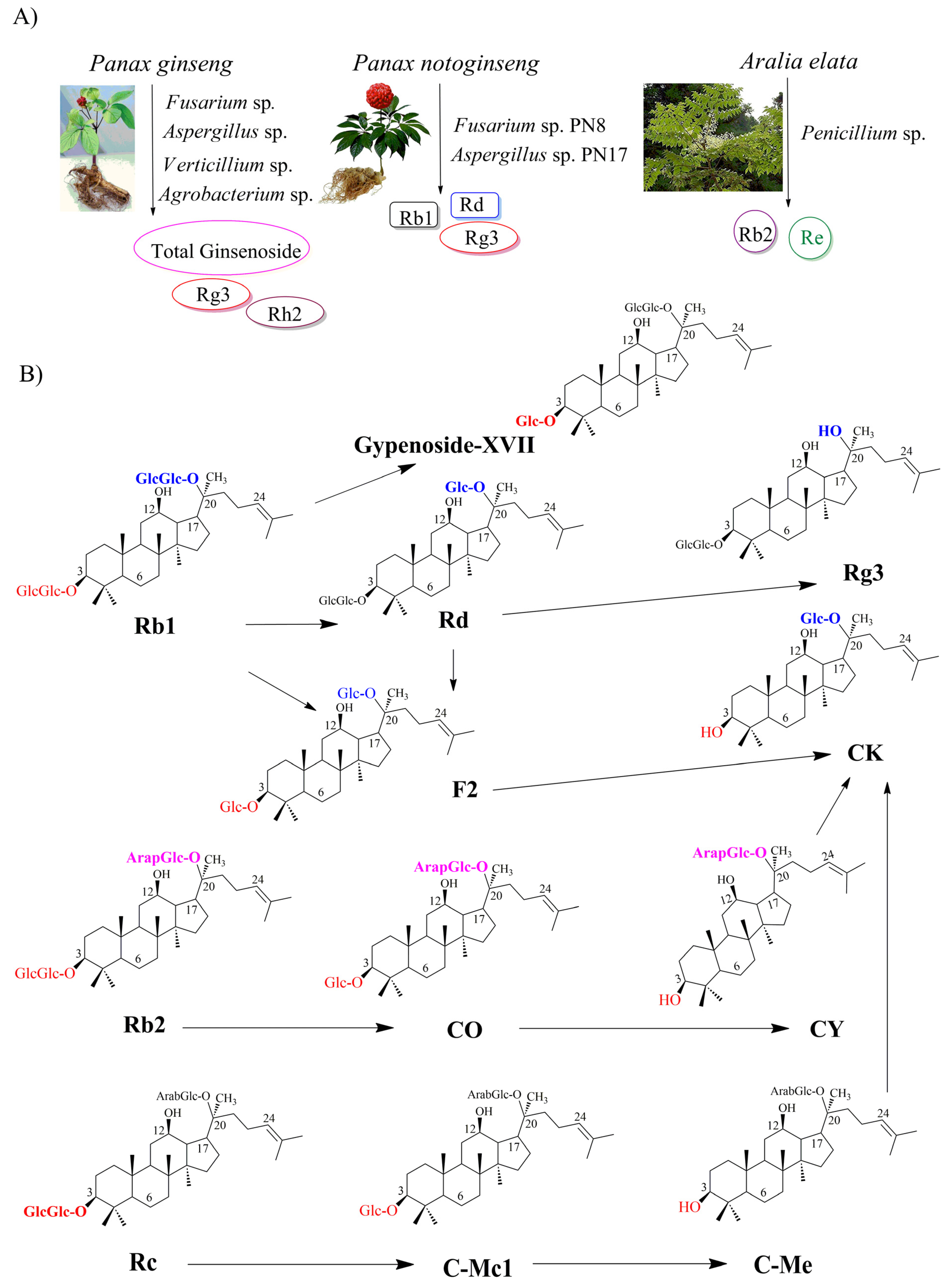
3.1. Bioproduction of Ginsenosides by Native Endophytes
3.2. Biotransformation of Major to Minor Ginsenosides by Endophytes
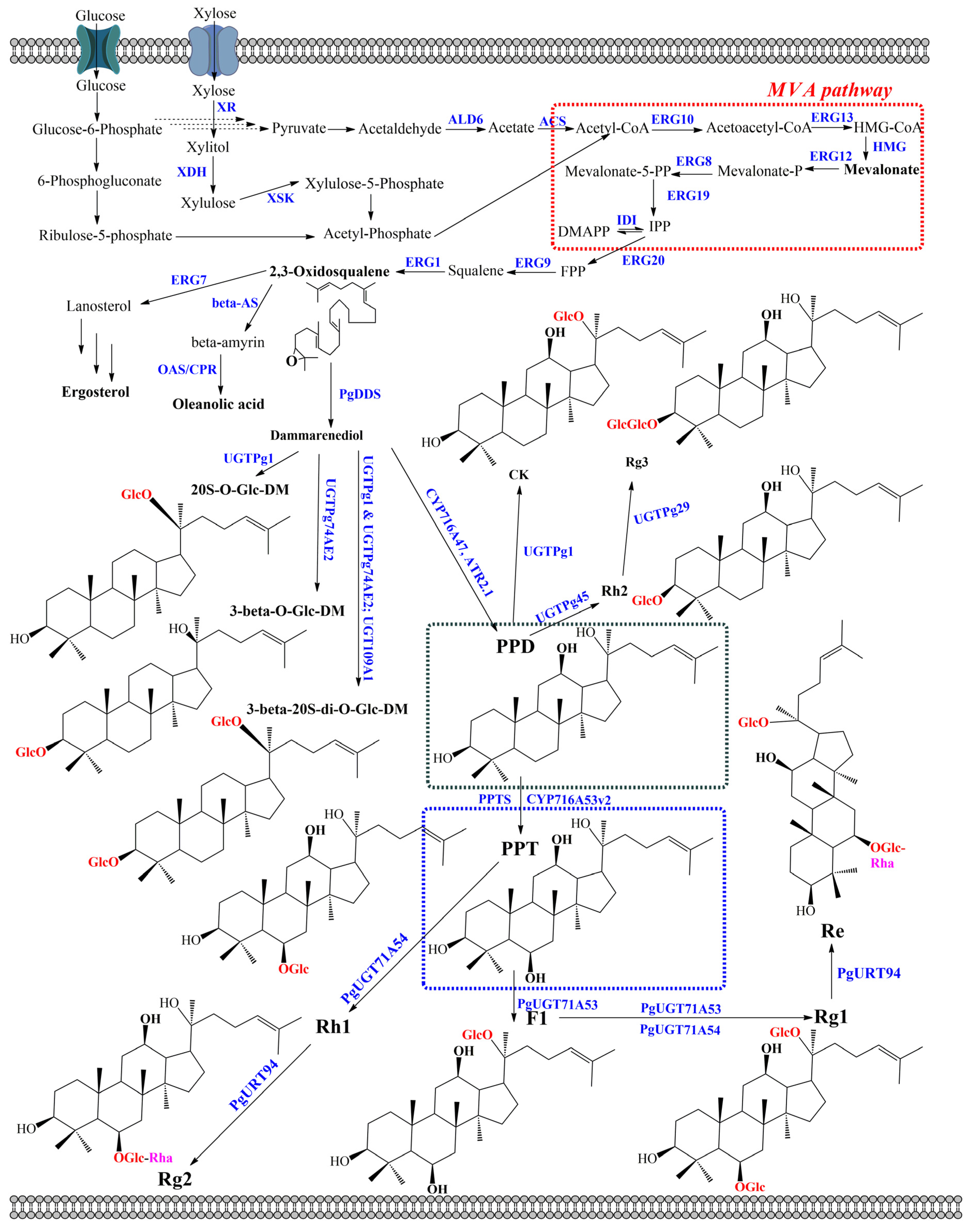
4. Ginsenoside Biosynthesis in Engineered Microorganisms
4.1. Ginsenoside Biosynthesis in Engineered Bacteria
4.2. Ginsenoside Production in Engineered Yeasts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Genes or Related Gene Cassettes | Products | Titer (mg/L) | Cultivation Condition | Major Media | Carbon Source | References |
|---|---|---|---|---|---|---|---|
| Saccharomyces cerevisiae | |||||||
| ZD-PPD-018 | tHMG1, AtCPR1, SynPgPPDS, ERG20, ERG1, ERG9 | PPD | 1189 | Fed-batch | SD | Glucose | [79] |
| DM | 1548 | ||||||
| D20RH18 | PgDDS, synPgPPDS, ATR2.1, tHMG1, ERG20, PgERG1, ERG9, UGTPg45 | Rh2 | 1.45 μmol/g DCW | Shake-flask | YPD | Glucose | [74] |
| D20RG1 | PgDDS, synPgPPDS, ATR2.1, tHMG1, ERG20, PgERG1, ERG9, UGTPg45, UGTPg29 | Rh3 | 3.49 μmol/g DCW | ||||
| ZW-Rh1-20 | ERG20, PgERG1, ERG9, tHMG1, CYP716A53v2, PgCPR1, UGTPg100 | Rh1 | 98.2 | Shake-flask | SC | Glucose | [71] |
| PPT | 3.5 | ||||||
| PPD | 43.4 | ||||||
| DM | 8.8 | ||||||
| ZW-F1-17 | ERG20, PgERG1, ERG9, tHMG1, CYP716A53v2, PgCPR1, UGTPg1 | F1 | 42.1 | ||||
| PPT | 13.9 | ||||||
| CK | 7.5 | ||||||
| PPD | 49.2 | ||||||
| DM | 3.5 | ||||||
| WLT-MVA5 | DS, PPDS-ATR1, ERG1, tHMG1, ERG9, ERG20, ERG10, ERG13, ERG12, ERG8, ERG19, IDI1, NCP1, ACSseL641P | PPD | 8090 | Fed-batch | YNBD | Glucose/ Ethanol | [80] |
| Y1CSH | HAC1, IDI1, ERG20, ERG9, ERG1, ERG7, synDS-GFP, tHMG1, synPgUGT74AE2 | 3β-O-Glc-DM | 2400 | Fed-batch | YPD | Glucose | [72] |
| Y2CSH | HAC1, IDI1, ERG20, ERG9, ERG1, ERG7, synDS-GFP, tHMG1, synUGTPg1 | 20S-O-Glc-DM | 5600 | ||||
| PPD-A3-sgRNA4 | PgDS and PgPPDS, PgCPR, tHMGR1, ERG1m, ∆ ERG7 | PPD | 294.5 | Shake-flask | YPD | Glucose | [71] |
| Rg1-02 | CYP716A53v2, PgUGT71A54, PgURT94, RHM | Rg2 | 1300 | Fed-batch | Synthetic | Glucose | [75] |
| Re-01 | CYP716A53v2, PgUGT71A53, PgUGT71A54, PgURT94, RHM | Re | 3600 | ||||
| CPX113436PPXP-ADH2 | ERG10, ERG13, tHMG1, ERG12, ERG8, ERG19, IDI1, ERG20, ERG9, ERG1, ERG7, PgDS, PgPPDS, PgCPR, ADH2, (Pex11p, Pex34p, and Atg36p) | PPD | 4.1 | Shake-flask | YPDO | Glucose and Ethanol | [81] |
| BY-V | ERG10, ERG13, tHMG1, ERG12, ERG8, IDI1, MVD1, ERG20, ERG9, ERG1, PgDDS, AtCPR1, PgPPDS, INO2, ∆LPP1, ∆ERG7 | PPD | 1550 | Shake-flask | YPD | Sugarcane molasses | [78] |
| 158,800 | Fed-batch | ||||||
| YFR | tHMG1, IDI1, ERG20, ERG9, ERG1, DS-GFP, PGM1, PGM2, INO2, ERG7, ERG1, PgUGT74AE2, UGTPg1, PP-DS, ATR2 | F2 | 21.0 | Shake-flask | YPD | Glucose | [73] |
| YSR | tHMG1, IDI1, ERG20, ERG9, ERG1, DS-GFP, UGTPg1, PGM1, PGM2, INO2, ERG7, ERG1, M7 (ΔUGT74AC1) | 3β-20S-Di-O-Glc-DM | 346.1 | Glucose | |||
| 2600 | Fed-batch | ||||||
| WEA | tHMG1, ERG10, ERG13,IDI, ERG20, ERG9, ERG1, UGD1, AeBAS1, AtATR2, AeCYP716A354, AeCSLM1, AeUGT74AG6 | Chikusetsusaponin IVa | NR | Shake-flask | SD | Glucose, Galactose | [82] |
| tHMG1, ERG10, ERG13,IDI, ERG20, ERG9, ERG1, UGD1, AeBAS1, AtATR2, AeCYP716A354, AeCSLM1, AeUGT73CB3 | Zingibroside R1 | NR | |||||
| ZY-M7(4)E1 PUA | ERG20, ERG1, ERG9, tHMG1, M7-1, ∆EGH1, PGM1, UGP1, PgPPDS-AtCPR2 | Rh2 | 300 | Fed-batch | SC | Glucose | [83] |
| Yarrowia lipolytica | |||||||
| Y14 | ΔLUL, XYL1, XYL2, ylXKS, DS, PPDS-linker-ATR1, tHMG1, ERG9, ERG20, TKL, TAL, TX | PPD | 300.63 | Fed-batch | YPD or YPX | Xylose | [84] |
| 167.17 | Glucose | ||||||
| YL-MVA-CK | tHMG1, ERG9, ERG20, OpDS, PPDS-linker2-ATR1, UGT1 | CK | 161.8 | Fed-batch | YPD | Glucose | [85] |
| Pichia pastoris | |||||||
| KDPEP | PgDDS-L3-PDZlig and ERG1-ER/kPDZ with p-[PgDDS-PDZlig]/[ERG1-PDZ] | DM | 0.10 mg/g DCW | Shake-flask | YPD | Glucose, methanol | [86] |
5. Challenges and Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiang, H.; Xu, P.; Qiu, H.; Wen, W.; Zhang, A.; Tong, S. Two-dimensional chromatography in screening of bioactive components from natural products. Phytochem. Anal. 2022, 33, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Erkisa, M.; Sariman, M.; Geyik, O.G.; Geyik, C.; Stanojkovic, T.; Ulukaya, E. Natural products as a promising therapeutic strategy to target cancer stem cells. Curr. Med. Chem. 2022, 29, 741–783. [Google Scholar] [CrossRef]
- Ekiert, H.M.; Szopa, A. Biological activities of natural products II. Molecules 2022, 27, 1519. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Shan, M.; Lan, X.; Fang, X.; Song, D.; Luo, H.; Wu, D. Anti-cancer effect and potential microRNAs targets of ginsenosides against breast cancer. Front. Pharmacol. 2022, 13, 1033017. [Google Scholar] [CrossRef]
- Yao, W.; Guan, Y. Ginsenosides in cancer: A focus on the regulation of cell metabolism. Biomed. Pharmacother. 2022, 156, 113756. [Google Scholar] [CrossRef]
- Gupta, S.P.; Sharma, A.; Patil, V.M. Molecular processes exploited as drug targets for cancer chemotherapy. Anticancer Agents Med. Chem. 2021, 21, 1638–1649. [Google Scholar]
- Seczyk, L.; Sugier, D.; Dervisoglu, G.; Ozdemir, F.A.; Kołodziej, B. Phytochemical profile, in vitro bioaccessibility, and anticancer potential of golden root (Rhodiola rosea L.) extracts. Food Chem. 2023, 404, 134779. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wu, S.D.; Chen, Y.; Li, X.Y.; Zhu, Q.; Nakayama, K.; Zhang, W.Q.; Weng, C.Z.; Zhang, J.; Wang, H.K.; et al. Alterations in gut microbiome and metabolomics in chronic hepatitis B infection-associated liver disease and their impact on peripheral immune response. Gut Microbes 2023, 15, 2155018. [Google Scholar] [CrossRef]
- Pandey, R.P.; Chu, L.L.; Kim, T.S.; Sohng, J.K. Bioconversion of tetracycline antibiotics to novel glucoside derivatives by single-vessel multienzymatic glycosylation. J. Microbiol. Biotechnol. 2018, 28, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.L.; Pandey, R.P.; Shin, J.Y.; Jung, H.J.; Sohng, J.K. Synthetic analog of anticancer drug daunorubicin from daunorubicinone using one-pot enzymatic UDP-recycling glycosylation. J. Mol. Catal. B Enzym. 2015, 124, 1–10. [Google Scholar] [CrossRef]
- Wilson, D. Endophyte: The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Hu, B.; Zhao, X.; Wang, E.; Zhou, J.; Li, J.; Chen, J.; Du, G. Efficient heterologous expression of cytochrome P450 enzymes in microorganisms for the biosynthesis of natural products. Crit. Rev. Biotechnol. 2022, 6, 1–15. [Google Scholar] [CrossRef]
- Wang, G.; Huang, M.; Nielsen, J. Exploring the potential of Saccharomyces cerevisiae for biopharmaceutical protein production. Curr. Opin. Biotechnol. 2017, 48, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Prabowo, C.P.S.; Eun, H.; Park, S.Y.; Cho, I.J.; Jiao, S.; Lee, S.Y. Escherichia coli as a platform microbial host for systems metabolic engineering. Essays Biochem. 2021, 65, 225–246. [Google Scholar]
- Le, T.H.; Lee, G.J.; Vu, H.K.; Kwon, S.W.; Nguyen, N.K.; Park, J.H.; Nguyen, M.D. Ginseng saponins in different parts of Panax vietnamensis. Chem. Pharm. Bull. 2015, 63, 950–954. [Google Scholar] [CrossRef]
- Lee, S.M.; Bae, B.S.; Park, H.W.; Ahn, N.G.; Cho, B.G.; Cho, Y.L.; Kwak, Y.S. Characterization of Korean Red Ginseng (Panax ginseng Meyer): History, preparation method, and chemical composition. J. Ginseng Res. 2015, 39, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, A.; Kim, J.H.; Kim, J.H.; Yi, Y.S.; Cho, J.Y. Functional role of ginseng-derived compounds in cancer. J. Ginseng Res. 2018, 42, 248–254. [Google Scholar] [CrossRef]
- Chu, L.L.; Montecillo, J.A.V.; Bae, H. Recent advances in the metabolic engineering of yeasts for ginsenoside biosynthesis. Front. Bioeng. Biotechnol. 2020, 8, 139. [Google Scholar] [CrossRef]
- Nag, S.A.; Qin, J.J.; Wang, W.; Wang, M.H.; Wang, H.; Zhang, R. Ginsenosides as anticancer agents: In vitro and in vivo activities, structure-activity relationships, and molecular mechanisms of action. Front. Pharmacol. 2012, 3, 25. [Google Scholar] [CrossRef]
- Xia, T.; Wang, Y.N.; Zhou, C.X.; Wu, L.M.; Liu, Y.; Zeng, Q.H.; Zhang, X.L.; Yao, J.H.; Wang, M.; Fang, J.P. Ginsenoside Rh2 and Rg3 inhibit cell proliferation and induce apoptosis by increasing mitochondrial reactive oxygen species in human leukemia Jurkat cells. Mol. Med. Rep. 2017, 15, 3591–3598. [Google Scholar] [CrossRef] [PubMed]
- Mohanan, P.; Subramaniyam, S.; Mathiyalagan, R.; Yang, D.C. Molecular signaling of ginsenosides Rb1, Rg1, and Rg3 and their mode of actions. J. Ginseng Res. 2018, 42, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cao, Y.; Luo, Y.; Liu, T.; Yan, G.; Chen, L.; Ji, L.; Wang, L.; Chen, B.; Yaseen, A.; et al. Two new triterpenoid saponins derived from the leaves of Panax ginseng and their anti-inflammatory activity. J. Ginseng Res. 2019, 43, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, Z.K.; Li, C.Y.; Liang, Y.Q.; Yang, F. Anticancer properties and pharmaceutical applications of ginsenoside compound K: A review. Chem. Biol. Drug Des. 2022, 99, 286–300. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Ding, M.; Xin, Y.; Xuan, Y.; Zhao, Y. Genotoxicity and subchronic toxicological study of a novel ginsenoside derivative 25-OCH3-PPD in beagle dogs. J. Ginseng Res. 2019, 43, 562–571. [Google Scholar] [CrossRef]
- Xia, T.; Wang, J.; Wang, Y.; Wang, Y.; Cai, J.; Wang, M.; Chen, Q.; Song, J.; Yu, Z.; Huang, W.; et al. Inhibition of autophagy potentiates anticancer property of 20(S)-ginsenoside Rh2 by promoting mitochondria-dependent apoptosis in human acute lymphoblastic leukaemia cells. Oncotarget 2016, 7, 27336–27349. [Google Scholar] [CrossRef]
- Shen, W.; Wei, Y.; Tang, D.; Jia, X.; Chen, B. Metabolite profiles of ginsenosides Rk1 and Rg5 in zebrafish using ultraperformance liquid chromatography/quadrupole-time-of-flight MS. J. Ginseng Res. 2017, 41, 78–84. [Google Scholar] [CrossRef]
- Zhang, H.; Park, S.; Huang, H.; Kim, E.; Yi, J.; Choi, S.K.; Ryoo, Z.; Kim, M. Anticancer effects and potential mechanisms of ginsenoside Rh2 in various cancer types (Review). Oncol. Rep. 2021, 45, 33. [Google Scholar] [CrossRef] [PubMed]
- Lyu, X.; Xu, X.; Song, A.; Guo, J.; Zhang, Y.; Zhang, Y. Ginsenoside Rh1 inhibits colorectal cancer cell migration and invasion in vitro and tumor growth in vivo. Oncol. Lett. 2019, 18, 4160–4166. [Google Scholar] [CrossRef]
- Kwon, H.J.; Lee, H.; Choi, G.E.; Kwon, S.J.; Song, A.Y.; Kim, S.J.; Choi, W.S.; Hwang, S.H.; Kim, S.C.; Kim, H.S. Ginsenoside F1 promotes cytotoxic activity of NK cells via insulin-like growth factor-1-dependent mechanism. Front. Immunol. 2018, 9, 2785. [Google Scholar] [CrossRef]
- de Oliveira Zanuso, B.; de Oliveira Dos Santos, A.R.; Miola, V.F.B.; Guissoni Campos, L.M.; Spilla, C.S.G.; Barbalho, S.M. Panax ginseng and aging related disorders: A systematic review. Exp. Gerontol. 2022, 161, 111731. [Google Scholar] [CrossRef]
- Cui, Y.; Su, Y.; Deng, L.; Wang, W. Ginsenoside-Rg5 inhibits retinoblastoma proliferation and induces apoptosis through suppressing BCL2 expression. Chemotherapy 2018, 63, 293–300. [Google Scholar] [CrossRef]
- Omomowo, O.I.; Babalola, O.O. Bacterial and fungal endophytes: Tiny giants with immense beneficial potential for plant growth and sustainable agricultural productivity. Microorganisms 2019, 7, 481. [Google Scholar] [CrossRef] [Green Version]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Gao, H.; Li, G.; Lou, H.X. Structural diversity and biological activities of novel secondary metabolites from endophytes. Molecules 2018, 23, 646. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Li, S.; Zhang, X.; Zhao, C. Biological activities of some new secondary metabolites isolated from endophytic fungi: A review study. Int. J. Mol. Sci. 2021, 22, 959. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, S.; Barac, T.; Greenberg, B.; Borremans, B.; Vangronsveld, J.; van der Lelie, D. Horizontal gene transfer to endogenous endophytic bacteria from poplar improves phytoremediation of toluene. Appl. Environ. Microbiol. 2005, 71, 8500–8505. [Google Scholar] [CrossRef] [PubMed]
- Rochin-Hernandez, L.S.; Rochin-Hernandez, L.J.; Flores-Cotera, L.B. Endophytes, a potential source of bioactive compounds to curtail the formation-accumulation of advanced glycation end products: A review. Molecules 2022, 27, 4469. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.L.; Dang, L.Z.; Li, J.F.; Zou, C.G.; Zhang, K.Q.; Li, G.H. Biotransformation of saponins by endophytes isolated from Panax notoginseng. Chem. Biodivers. 2013, 10, 2021–2031. [Google Scholar] [CrossRef]
- Jin, Z.; Gao, L.; Zhang, L. Antimicrobial activity of saponins produced by two novel endophytic fungi from Panax notoginseng. Nat. Prod. Res. 2013, 31, 2700–2703. [Google Scholar] [CrossRef]
- Wu, H.; Yang, H.Y.; You, X.L.; Li, Y.H. Diversity of endophytic fungi from roots of Panax ginseng and their saponin yield capacities. Springerplus 2013, 2, 107. [Google Scholar] [CrossRef]
- Wu, H.; Yang, H.; You, X.; Li, Y. Isolation and characterization of saponin-producing fungal endophytes from Aralia elata in Northeast China. Int. J. Mol. Sci. 2012, 13, 16255–16266. [Google Scholar] [CrossRef]
- Yan, H.; Jin, H.; Fu, Y.; Yin, Z.; Yin, C. Production of rare ginsenosides Rg3 and Rh2 by endophytic bacteria from Panax ginseng. J. Agric. Food Chem. 2019, 67, 8493–8499. [Google Scholar] [CrossRef]
- Qi, L.W.; Wang, C.Z.; Yuan, C.S. Ginsenosides from American ginseng: Chemical and pharmacological diversity. Phytochemistry 2011, 72, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Lu, Z.; Teng, Y.; Guo, Y.; Liu, S. Structural characterization of ginsenosides from flower buds of Panax ginseng by RRLC-Q-TOF MS. J. Chromatogr. Sci. 2016, 54, 136–143. [Google Scholar] [PubMed]
- Xiu, Y.; Li, X.; Sun, X.; Xiao, D.; Miao, R.; Zhao, H.; Liu, S. Simultaneous determination and difference evaluation of 14 ginsenosides in Panax ginseng roots cultivated in different areas and ages by high-performance liquid chromatography coupled with triple quadrupole mass spectrometer in the multiple reaction-monitoring mode combined with multivariate statistical analysis. J. Ginseng Res. 2019, 43, 508–516. [Google Scholar] [PubMed]
- Wei, G.; Chen, Z.; Wang, B.; Wei, F.; Zhang, G.; Wang, Y.; Zhu, G.; Zhou, Y.; Zhao, Q.; He, M.; et al. Endophytes isolated from Panax notoginseng converted ginsenosides. Microb. Biotechnol. 2021, 14, 1730–1746. [Google Scholar] [CrossRef]
- Fu, Y.; Yin, Z.H.; Wu, L.P.; Yin, C.R. Biotransformation of ginsenoside Rb1 to ginsenoside C-K by endophytic fungus Arthrinium sp. GE 17-18 isolated from Panax ginseng. Lett. Appl. Microbiol. 2016, 63, 196–201. [Google Scholar] [CrossRef]
- Fu, Y.; Yin, Z.H.; Yin, C.Y. Biotransformation of ginsenoside Rb1 to ginsenoside Rg3 by endophytic bacterium Burkholderia sp. GE 17-7 isolated from Panax ginseng. J. Appl. Microbiol. 2017, 122, 1579–1585. [Google Scholar] [CrossRef]
- Fu, Y. Biotransformation of ginsenoside Rb1 to Gyp-XVII and minor ginsenoside Rg3 by endophytic bacterium Flavobacterium sp. GE 32 isolated from Panax ginseng. Lett. Appl. Microbiol. 2019, 68, 134–141. [Google Scholar] [CrossRef]
- Cui, L.; Wu, S.Q.; Zhao, C.A.; Yin, C.R. Microbial conversion of major ginsenosides in ginseng total saponins by Platycodon grandiflorum endophytes. J. Ginseng Res. 2016, 40, 366–374. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, Y.; Gu, M.; Liu, Z.; Zhang, J.; Zeng, Q.; Zhu, D. Biotransformation of ginsenoside Rc to Rd by endophytic bacterium Bacillus sp. G9y isolated from Panax quinquefolius. Antonie Van Leeuwenhoek 2021, 114, 437–444. [Google Scholar] [CrossRef]
- An, C.; Ma, S.; Shi, X.; Liu, C.; Ding, H.; Xue, W. Diversity and ginsenoside biotransformation potential of cultivable endophytic fungi associated with Panax bipinnatifidus var. bipinnatifidus in Qinling Mountains, China. Front. Pharmacol. 2022, 13, 762862. [Google Scholar] [PubMed]
- Yu, L.; Chen, Y.; Shi, J.; Wang, R.; Yang, Y.; Yang, L.; Zhao, S.; Wang, Z. Biosynthesis of rare 20(R)-protopanaxadiol/protopanaxatriol type ginsenosides through Escherichia coli engineered with uridine diphosphate glycosyltransferase genes. J. Ginseng Res. 2019, 43, 116–124. [Google Scholar] [CrossRef]
- Santoscoy, M.C.; Jarboe, L.R. Production of cholesterol-like molecules impacts Escherichia coli robustness, production capacity, and vesicle trafficking. Metab. Eng. 2022, 73, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.H.; Kim, D.J.; Jung, S.C.; Kim, S.C.; Im, W.T. Enhanced production of Gypenoside LXXV using a novel ginsenoside-transforming β-Glucosidase from Ginseng-cultivating soil bacteria and its anti-cancer property. Molecules 2017, 22, 844. [Google Scholar] [CrossRef]
- Pei, J.; Wu, T.; Yao, T.; Zhao, L.; Ding, G.; Wang, Z.Z.; Xiao, W. Biotransformation of ginsenosides Re and Rg1 into Rg2 and Rh1 by thermostable β-Glucosidase from Thermotoga thermarum. Chem. Nat. Compd. 2017, 53, 472–477. [Google Scholar] [CrossRef]
- Shin, K.C.; Kim, T.H.; Choi, J.H.; Oh, D.K. Complete biotransformation of Protopanaxadiol-type ginsenosides to 20- O-β-Glucopyranosyl-20(S)-protopanaxadiol using a novel and thermostable β-Glucosidase. J. Agric. Food Chem. 2018, 66, 2822–2829. [Google Scholar] [CrossRef]
- Siddiqi, M.Z.; Hashmi, M.S.; Oh, J.M.; Chun, S.; Im, W.T. Identification of novel glycoside hydrolases via whole genome sequencing of Niabella ginsenosidivorans for production of various minor ginsenosides. 3 Biotech 2019, 9, 258. [Google Scholar] [CrossRef]
- Siddiqi, M.Z.; Ximenes, H.A.; Song, B.K.; Park, H.Y.; Lee, W.H.; Han, H.; Im, W.T. Enhanced production of ginsenoside Rh2(S) from PPD-type major ginsenosides using BglSk cloned from Saccharibacillus kuerlensis together with two glycosidases in series. Saudi. J. Biol. Sci. 2021, 28, 4668–4676. [Google Scholar] [CrossRef]
- Kim, S.A.; Shin, K.C.; Oh, D.K. Complete biotransformation of protopanaxadiol-type ginsenosides into 20-O-β-Glucopyranosyl-20(S)-protopanaxadiol by permeabilized recombinant Escherichia coli cells coexpressing β-glucosidase and chaperone genes. J. Agric. Food Chem. 2019, 67, 8393–8401. [Google Scholar] [CrossRef]
- Li, W.; Ren, M.; Duo, L.; Li, J.; Wang, S.; Sun, Y.; Li, M.; Ren, W.; Hou, Q.; Yu, J.; et al. Fermentation characteristics of Lactococcus lactis subsp. lactis isolated from naturally fermented dairy products and screening of potential starter isolates. Front. Microbiol. 2020, 11, 1794. [Google Scholar]
- Kataoka, N.; Matsutani, M.; Matsushita, K.; Yakushi, T. Stepwise metabolic engineering of Corynebacterium glutamicum for the production of phenylalanine. Gen. Appl. Microbiol. 2022. [Google Scholar] [CrossRef]
- Li, L.; Lee, S.J.; Yuan, Q.P.; Im, W.T.; Kim, S.C.; Han, N.S. Production of bioactive ginsenoside Rg3(S) and compound K using recombinant Lactococcus lactis. J. Ginseng Res. 2018, 42, 412–418. [Google Scholar] [CrossRef]
- Cui, C.H.; Jeon, B.M.; Fu, Y.; Im, W.T.; Kim, S.C. High-density immobilization of a ginsenoside-transforming β-glucosidase for enhanced food-grade production of minor ginsenosides. Appl. Microbiol. Biotechnol. 2019, 103, 7003–7015. [Google Scholar] [CrossRef]
- Patra, P.; Das, M.; Kundu, P.; Ghosh, A. Recent advances in systems and synthetic biology approaches for developing novel cell-factories in non-conventional yeasts. Biotechnol. Adv. 2021, 47, 107695. [Google Scholar]
- Yi, X.; Alper, H.S. Considering strain variation and non-type strains for yeast metabolic engineering applications. Life 2022, 12, 510. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Li, L.; Song, W.; Li, M.; Hua, X.; Wang, Y.; Yuan, J.; Xue, Z. Natural products of pentacyclic triterpenoids: From discovery to heterologous biosynthesis. Nat. Prod. Rep. 2022. [Google Scholar] [CrossRef]
- Luo, Y.; Jiang, Y.; Chen, L.; Li, C.; Wang, Y. Applications of protein engineering in the microbial synthesis of plant triterpenoids. Synth. Syst. Biotechnol. 2022, 8, 20–32. [Google Scholar] [CrossRef]
- Lim, S.H.; Baek, J.I.; Jeon, B.M.; Seo, J.W.; Kim, M.S.; Byun, J.Y.; Park, S.H.; Kim, S.J.; Lee, J.Y.; Lee, J.H.; et al. CRISPRi-Guided metabolic flux engineering for enhanced protopanaxadiol production in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2021, 22, 11836. [Google Scholar] [CrossRef]
- Yan, X.; Fan, Y.; Wei, W.; Wang, P.; Liu, Q.; Wei, Y.; Zhang, L.; Zhao, G.; Yue, J.; Zhou, Z. Production of bioactive ginsenoside compound K in metabolically engineered yeast. Cell Res. 2014, 24, 770–773. [Google Scholar] [CrossRef]
- Wei, W.; Wang, P.; Wei, Y.; Liu, Q.; Yang, C.; Zhao, G.; Yue, J.; Yan, X.; Zhou, Z. Characterization of Panax ginseng UDP-Glycosyltransferases catalyzing protopanaxatriol and biosyntheses of bioactive ginsenosides F1 and Rh1 in metabolically engineered yeasts. Mol. Plant. 2015, 8, 1412–1424. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.F.; Gu, A.D.; Liang, L.; Li, Y.; Gong, T.; Chen, J.J.; Chen, T.J.; Yang, J.L.; Zhu, P. Construction and optimization of microbial cell factories for sustainable production of bioactive dammarenediol-II glucosides. Green. Chem. 2019, 21, 3286–3299. [Google Scholar] [CrossRef]
- Jiang, F.; Zhou, C.; Li, Y.; Deng, H.; Gong, T.; Chen, J.; Chen, T.; Yang, J.; Zhu, P. Metabolic engineering of yeasts for green and sustainable production of bioactive ginsenosides F2 and 3β,20S-Di-O-Glc-DM. Acta Pharm. Sin. B 2022, 12, 3167–3176. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wei, Y.; Fan, Y.; Liu, Q.; Wei, W.; Yang, C.; Zhang, L.; Zhao, G.; Yue, J.; Yan, X.; et al. Production of bioactive ginsenosides Rh2 and Rg3 by metabolically engineered yeasts. Metab. Eng. 2015, 29, 97–105. [Google Scholar] [CrossRef]
- Li, C.; Yan, X.; Xu, Z.; Wang, Y.; Shen, X.; Zhang, L.; Zhou, Z.; Wang, P. Pathway elucidation of bioactive rhamnosylated ginsenosides in Panax ginseng and their de novo high-level production by engineered Saccharomyces cerevisiae. Commun. Biol. 2022, 5, 775. [Google Scholar] [CrossRef]
- Dai, L.; Li, J.; Yang, J.; Zhu, Y.; Men, Y.; Zeng, Y.; Cai, Y.; Dong, C.; Dai, Z.; Zhang, X.; et al. Use of a promiscuous glycosyltransferase from Bacillus subtilis 168 for the enzymatic synthesis of novel Protopanaxatriol-type ginsenosides. J. Agric. Food Chem. 2018, 66, 943–949. [Google Scholar] [CrossRef]
- Dai, L.; Qin, L.; Hu, Y.; Huang, J.W.; Hu, Z.; Min, J.; Sun, Y.; Guo, R.T. Structural dissection of unnatural ginsenoside-biosynthetic UDP-glycosyltransferase Bs-YjiC from Bacillus subtilis for substrate promiscuity. Biochem. Biophys. Res. Commun. 2021, 534, 73–78. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, J.; Peng, L.; Meng, L.; Diao, M.; Jiang, S.; Li, J.; Xie, N. High-yield production of protopanaxadiol from sugarcane molasses by metabolically engineered Saccharomyces cerevisiae. Microb. Cell Fact. 2022, 21, 230. [Google Scholar] [CrossRef]
- Dai, Z.; Liu, Y.; Zhang, X.; Shi, M.; Wang, B.; Wang, D.; Huang, L.; Zhang, X. Metabolic engineering of Saccharomyces cerevisiae for production of ginsenosides. Metab. Eng. 2013, 20, 146–156. [Google Scholar] [CrossRef]
- Zhao, F.L.; Bai, P.; Nan, W.H.; Li, D.H.; Zhang, C.B.; Lu, C.Z.; Qi, H.S.; Lu, W. A modular engineering strategy for high-level production of protopanaxadiol from ethanol by Saccharomyces cerevisiae. AIChE J. 2018, 65, 866–874. [Google Scholar] [CrossRef]
- Choi, B.H.; Kang, H.J.; Kim, S.C.; Lee, P.C. Organelle engineering in yeast: Enhanced production of protopanaxadiol through manipulation of peroxisome proliferation in Saccharomyces cerevisiae. Microorganisms 2022, 10, 650. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Ri, H.C.; An, Z.; Wang, X.; Zhou, J.N.; Zheng, D.; Wu, H.; Wang, P.; Yang, J.; et al. Deletion and tandem duplications of biosynthetic genes drive the diversity of triterpenoids in Aralia elata. Nat. Commun. 2022, 13, 2224. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Yang, G.Y.; Chen, X.; Liu, Q.; Zhang, X.; Deng, Z.; Feng, Y. Biosynthesis of plant-derived ginsenoside Rh2 in yeast via repurposing a key promiscuous microbial enzyme. Metab. Eng. 2017, 42, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xu, S.; Gao, X.; Li, M.; Li, D.; Lu, W. Enhanced protopanaxadiol production from xylose by engineered Yarrowia lipolytica. Microb. Cell Fact. 2019, 18, 83. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, Y.; Zhang, C.; Sun, J.; Zhou, Z.; Lu, W. Production of triterpene ginsenoside compound K in the non-conventional yeast Yarrowia lipolytica. J. Agric. Food. Chem. 2019, 67, 2581–2588. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Gao, X.; Liu, X.; Wang, Y.; Yang, S.; Wang, F.; Ren, Y. Enhancing biosynthesis of a ginsenoside precursor by self-assembly of two key enzymes in Pichia pastoris. J. Agric. Food Chem. 2016, 64, 3380–3385. [Google Scholar] [CrossRef]
- Sun, Z.; Meng, H.; Li, J.; Wang, J.; Li, Q.; Wang, Y.; Zhang, Z.S. Identification of novel knockout targets for improving terpenoids biosynthesis in Saccharomyces cerevisiae. PLoS ONE 2014, 9, e112615. [Google Scholar] [CrossRef]
- Dong, L.; Cheng, R.; Xiao, L. Diversity and composition of bacterial endophytes among plant parts of Panax notoginseng. Chin. Med. 2018, 13, 41. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Metagenomics methods for the study of plant-associated microbial communities: A review. J. Microbiol. Methods 2020, 170, 105860. [Google Scholar] [CrossRef]
- Tang, K.; Zhang, Y.; Lin, D.; Han, Y.; Chen, C.A.; Wang, D.; Lin, Y.S.; Sun, J.; Zheng, Q.; Jiao, N. Cultivation-independent and cultivation-dependent analysis of microbes in the shallow-sea hydrothermal system off kueishantao island, Taiwan: Unmasking heterotrophic bacterial diversity and functional capacity. Front. Microbiol. 2018, 9, 279. [Google Scholar] [CrossRef]
- Misra, B.B.; Langefeld, C.D.; Olivier, M. Integrated omics: Tools, advances, and future approaches. J. Mol. Endocrinol. 2019, 62, R21–R45. [Google Scholar] [CrossRef]
- Hong, C.E.; Kim, J.U.; Lee, J.W.; Bang, K.H.; Jo, I.H. Metagenomic analysis of bacterial endophyte community structure and functions in Panax ginseng at different ages. 3 Biotech 2019, 9, 300. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Lin, Y.; Wang, Y.; Li, X.; Han, Y.; Wang, K.; Sun, C.; Wang, Y.; Zhang, M. Transcriptome analysis identifies strong candidate genes for ginsenoside biosynthesis and reveals its underlying molecular mechanism in Panax ginseng CA Meyer. Sci. Rep. 2019, 9, 615. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Kim, H.Y.; Kim, H.S.; Jung, S.H. Transcriptome analysis of Panax ginseng response to high light stress. J. Ginseng Res. 2020, 44, 312–320. [Google Scholar] [CrossRef]
- Sun, W.; Qin, L.; Xue, H.; Yu, Y.; Ma, Y.; Wang, Y.; Li, C. Novel trends for producing plant triterpenoids in yeast. Crit. Rev. Biotechnol. 2019, 39, 618–632. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wei, Y.M.; Zhou, X.W.; Lin, J.; Sun, X.F.; Tang, K.X. Agrobacterium tumefaciens-mediated genetic transformation of the taxol-producing endophytic fungus Ozonium sp. EFY21. Genet. Mol. Res. 2013, 12, 2913–2922. [Google Scholar] [CrossRef]
- Noushahi, H.A.; Khan, A.H.; Noushahi, U.F.; Hussain, M.; Javed, T.; Zafar, M.; Batool, M.; Ahmed, U.; Liu, K.; Harrison, M.T.; et al. Biosynthetic pathways of triterpenoids and strategies to improve their biosynthetic efficiency. Plant Growth Regul. 2022, 97, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wei, W.; Ye, W.; Li, X.; Zhao, W.; Yang, C.; Li, C.; Yan, X.; Zhou, Z. Synthesizing ginsenoside Rh2 in Saccharomyces cerevisiae cell factory at high-efficiency. Cell Discov. 2019, 5, 5. [Google Scholar] [CrossRef] [Green Version]




| Structure | Name | R1 | R2 | R3 | R4 | Cellular Mechanisms | Ref. |
|---|---|---|---|---|---|---|---|
| Protopanaxadiol (PPD) Type | |||||||
 | Ra1 | Glc2-Glc | - | Glc6-Ara(p)4-Xyl | - | Not reported | |
| Rb1 | Glc2-Glc | - | Glc6-Glc | - | Inhibition of invasion and migration | [17,21] | |
| Rb2 | Glc2-Glc | - | Glc6-Ara(p) | - | Inhibition of metastasis and proliferation | [17] | |
| Rc | Glc2-Glc | - | Glc6-Ara(f) | - | Anti-proliferative activity | [19] | |
| Rd | Glc2-Glc | - | Glc | - | Inhibit proliferation; Inhibit angiogenesis | [4] | |
| Rg3 | Glc2-Glc | - | H | - | Repression of cell proliferation and induce apoptosis | [20,25] | |
| Rh2 | Glc | - | H | - | Modulation of cell cycle; Regulation of inflammatory response molecules | [20,27] | |
| F2 | Glc | - | Glc | - | Inhibit proliferation | [4] | |
| CK | H | - | Glc | - | Modulation of growth factors and regulation of transcription factors; Induce apoptosis | [5,23] | |
| Protopanaxatriol (PPT)-type | |||||||
 | Re | OH | Glc2-Rha | Glc | - | Not reported | |
| Rf | OH | Glc2-Glc | H | - | Cell cycle arrest and apoptosis | [19] | |
| Rg1 | OH | Glc | Glc | - | Induce apoptosis; Repression of cell proliferation | [4,21] | |
| Rg2 | OH | Glc2-Rha | H | - | Induce apoptosis; Repression of cell proliferation | [4] | |
| Rh1 | OH | Glc | H | - | Regulation of gene coding for metalloproteinase; Repression of cell proliferation | [5,28] | |
| F1 | OH | H | Glc | - | Modulation of death receptor | [29] | |
| Notoginsenoside R1 | H | Glc2-Xyl Glc | Glc | - | Regulation of inflammatory response molecules | [19] | |
| Ocotillol-type | |||||||
 | Majonoside R2 | OH | Glc2-Xyl | - | - | Not reported | |
| Vinaginsenoside R1 | OH | Ac-Glc2-Rha | - | - | Not reported | ||
| Oleanolic acid type | |||||||
 | RO | GlcUA-Glc | - | - | Glc | Not reported | |
| ROA | GlcUA-Glc | - | - | Glc6-Glc | Not reported | ||
| Host | Endophytic Strains | Type of Compounds/ Biotransformation Pathway | Major Media | Titer (mg/mL) | References |
| Ginsenoside production by native endoyphtes | |||||
| Aralia elata | Penicillium sp. | Rb2, Re | PDA liquid | 2.049 | [41] |
| P. ginseng | Fusarium sp. | Total ginsenoside | PDA liquid | 0.181 | [40] |
| Aspergillus sp. | 0.144 | ||||
| Verticillium sp. | 0.144 | ||||
| P. notoginseng | Fusarium sp. PN8 | Rb1, Rd, and Rg3 | PDA liquid | 1.061 | [39] |
| Aspergillus sp. PN17 | Re, Rd, and Rg3 | 0.583 | |||
| P. ginseng | Agrobacterium sp. | Rg3 | LL medium | 62.20 mg L−1 | [42] |
| Rh2 | 18.60 mg L−1 | ||||
| Biotransformation of major to rare ginsenosides by endophytes | |||||
| P. ginseng | Arthrinium sp. | Rb1 → Rd → F2 → CK | PDA liquid | NA | [47] |
| Burkholderia sp. | Rb1 → Rd → Rg3 | PDA liquid | NA | [48] | |
| Flavobacterium sp. | Rb1 → Gyp-XVII | PDA liquid | NA | [49] | |
| Platycodon grandiflorum | Luteibacter sp. | Rb1 → Rd → F2 | LL medium | 0.06692 | [50] |
| Rb1 → Rd → F2 → CK | 0.03323 | ||||
| Rb2 → CO → CY → CK | |||||
| Rc →CMc1 → CMe → CK | |||||
| Rg1 → Rh1 | NA | ||||
| P. notoginseng | Fusarium oxysporum or Fusarium sp. | Rb1 → CK | LB medium | 0.02 | [38] |
| Rb1 → F2 | 0.025 | ||||
| Nodulisporium sp. | Re → 6-O-[α-L-Rhamnopyranosyl-(1→2)-β-D-glucopyranosyl]-20-O-β-glucopyranosyl-dammarane-3,6,12,20,24,25-hexaol | 0.125 | |||
| Vinaginsenoside R13 | 0.09 | ||||
| Fusarium oxysporum Nodulisporium sp. Bacillus sp. | Rg1 → Vinaginsenoside R22 | 0.065 | |||
| Nodulisporium sp. | Rh1 → Pseudo-ginsenoside RT4 | 0.075 | |||
| Fusarium oxysporum | Rh1 → PPT | 0.02 | |||
| Brevundimonas sp. | Rh1 → Rg1 | 0.15 | |||
| Rh1 → Vinaginsenoside R15 | 0.05 | ||||
| Bacillus sp. | Rh1 → (20S)-3-O-β-D-glucopyranosyl-6-O-β-D-glucopyranosylprotopanaxatriol | 0.07 | |||
| P. notoginseng | Enterobacter chengduensis | Rg1 → F1 | PDA medium | 13.24%; | [46] |
| Trichoderma koningii | Rb1 → Rd | 40.00% | |||
| Rb1 → Rg3 | 32.31%; | ||||
| Penicillium chermesinum | Rb1 → Rd | 74.24% | |||
| P. quinquefolius | Bacillus sp. G9y | Rc → Rd | Beef extract peptone | 100% | [51] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, L.L.; Huy, N.Q.; Tung, N.H. Microorganisms for Ginsenosides Biosynthesis: Recent Progress, Challenges, and Perspectives. Molecules 2023, 28, 1437. https://doi.org/10.3390/molecules28031437
Chu LL, Huy NQ, Tung NH. Microorganisms for Ginsenosides Biosynthesis: Recent Progress, Challenges, and Perspectives. Molecules. 2023; 28(3):1437. https://doi.org/10.3390/molecules28031437
Chicago/Turabian StyleChu, Luan Luong, Nguyen Quang Huy, and Nguyen Huu Tung. 2023. "Microorganisms for Ginsenosides Biosynthesis: Recent Progress, Challenges, and Perspectives" Molecules 28, no. 3: 1437. https://doi.org/10.3390/molecules28031437
APA StyleChu, L. L., Huy, N. Q., & Tung, N. H. (2023). Microorganisms for Ginsenosides Biosynthesis: Recent Progress, Challenges, and Perspectives. Molecules, 28(3), 1437. https://doi.org/10.3390/molecules28031437