
Resveratrol, a New Allosteric Effector of Hemoglobin, Enhances Oxygen Supply Efficiency and Improves Adaption to Acute Severe Hypoxia

 , ,
, ,
Abstract
:1. Introduction
2. Results
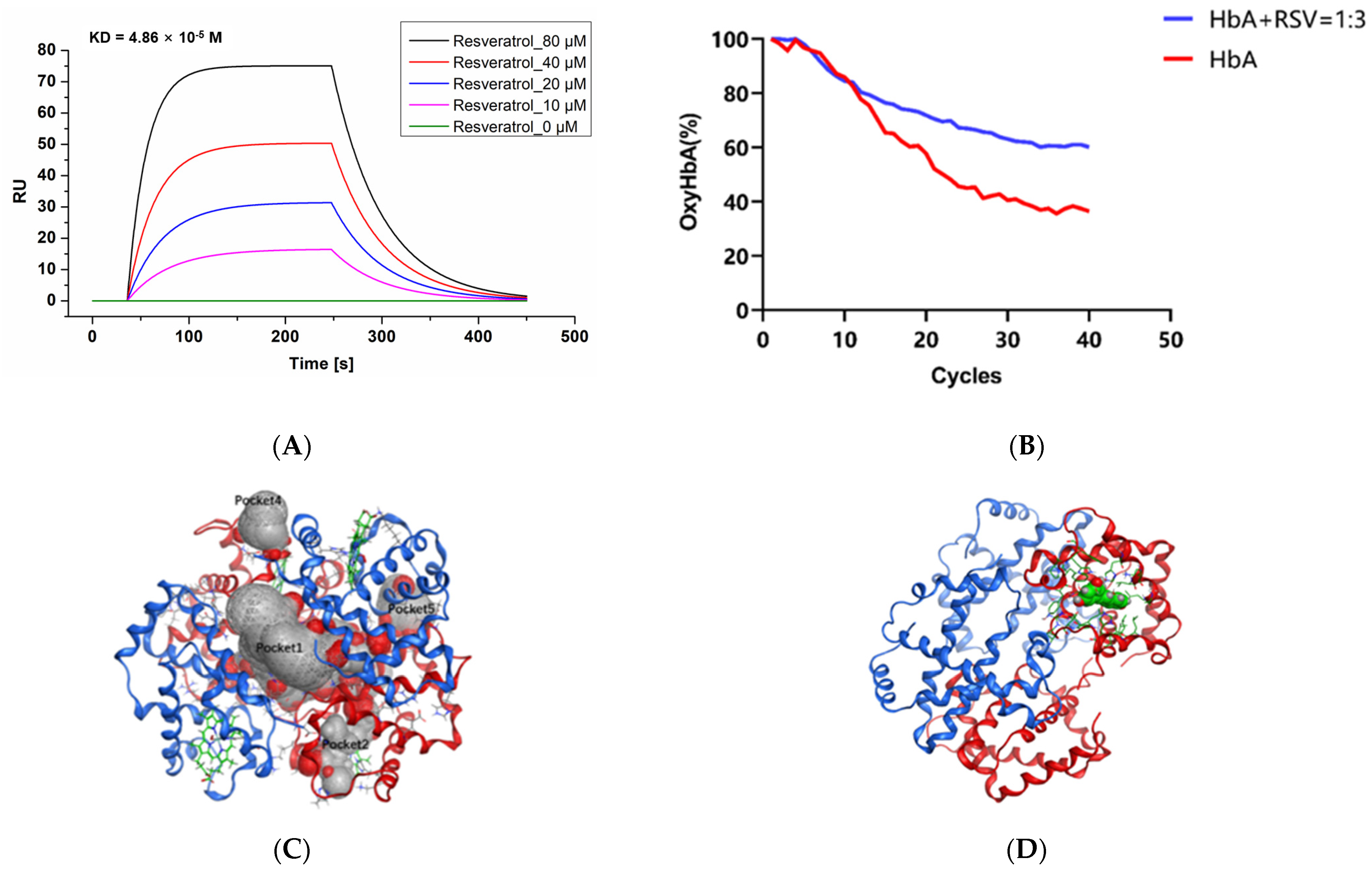
2.1. Resveratrol Binds to HbA with Concentration Gradients
2.2. Resveratrol Delays the Oxygen Release of HbA
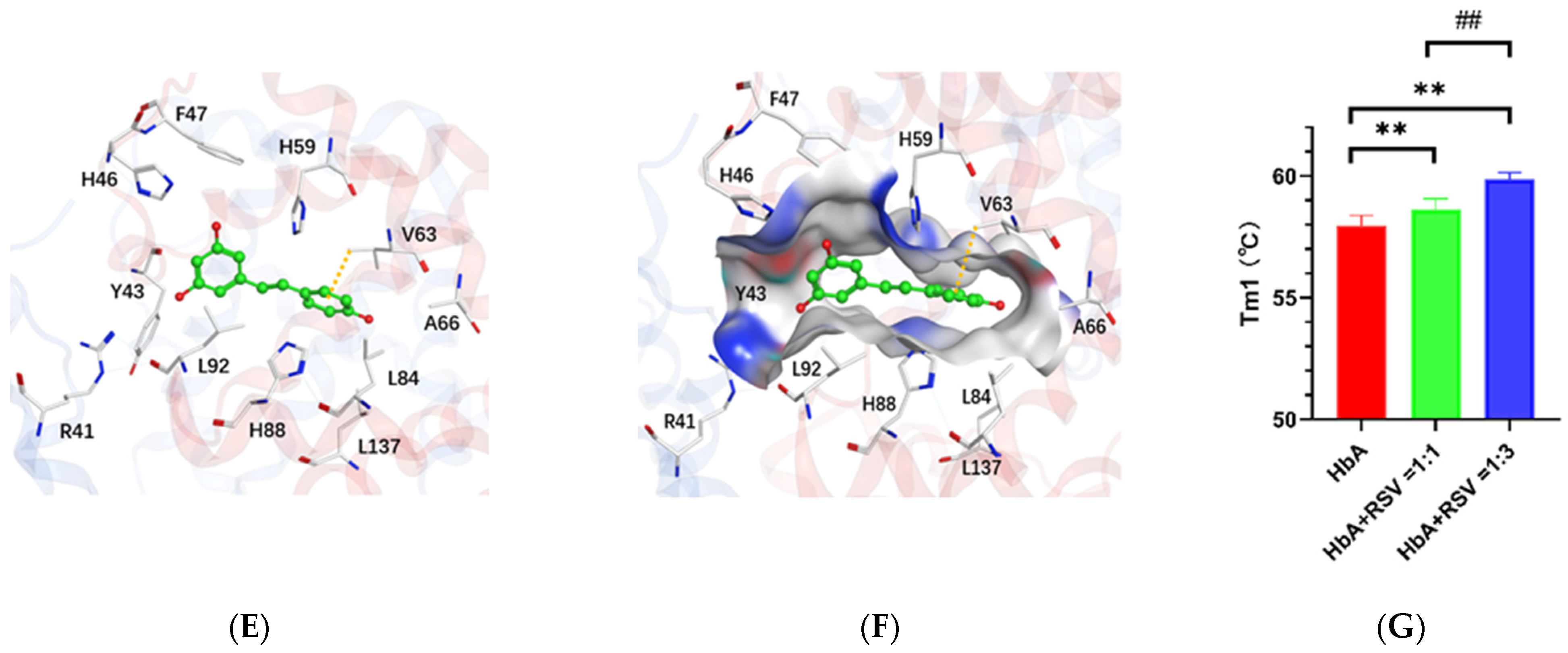
2.3. Resveratrol Binds the Heme Region of the Hemoglobin a Subunit
2.4. Resveratrol Stabilizes the Conformational Structure of HbA
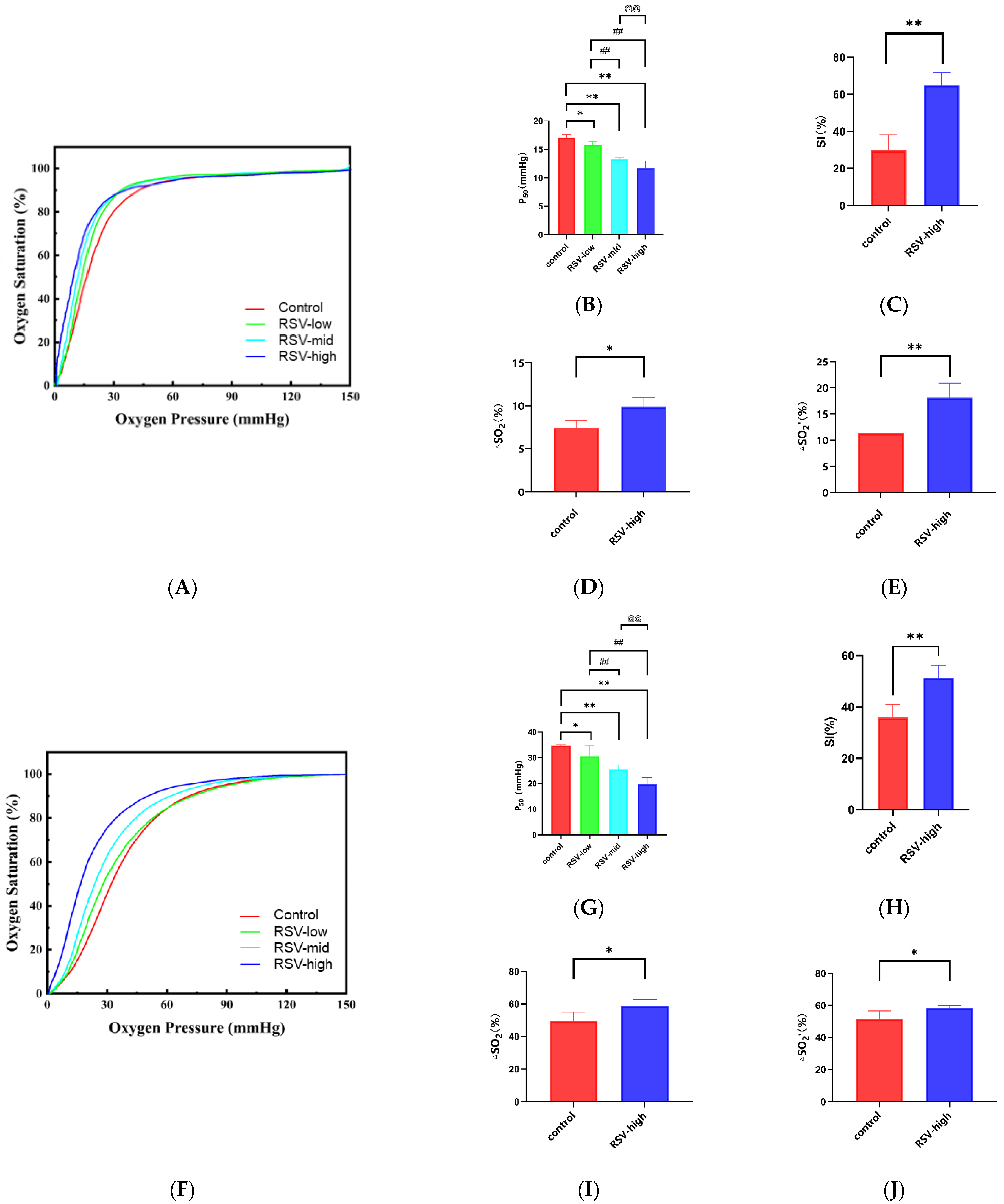
2.5. Resveratrol Enhances the Oxygen Supply Efficiency Ex Vivo
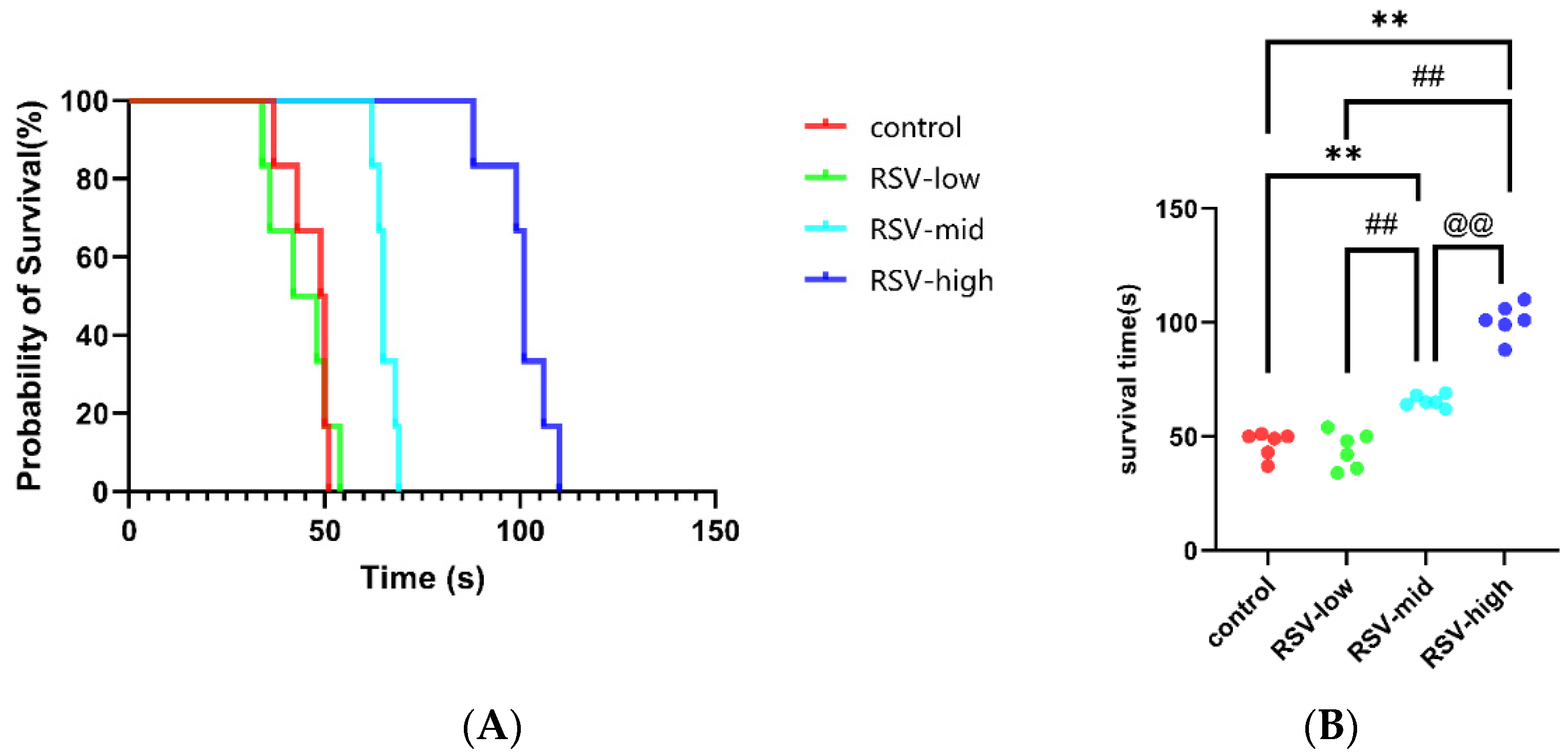
2.6. Resveratrol Significantly Increases The Survival Time of Mice under Acute Hypoxic Asphyxia
2.7. Resveratrol Enhances Adaption to Acute Severe Hypoxia
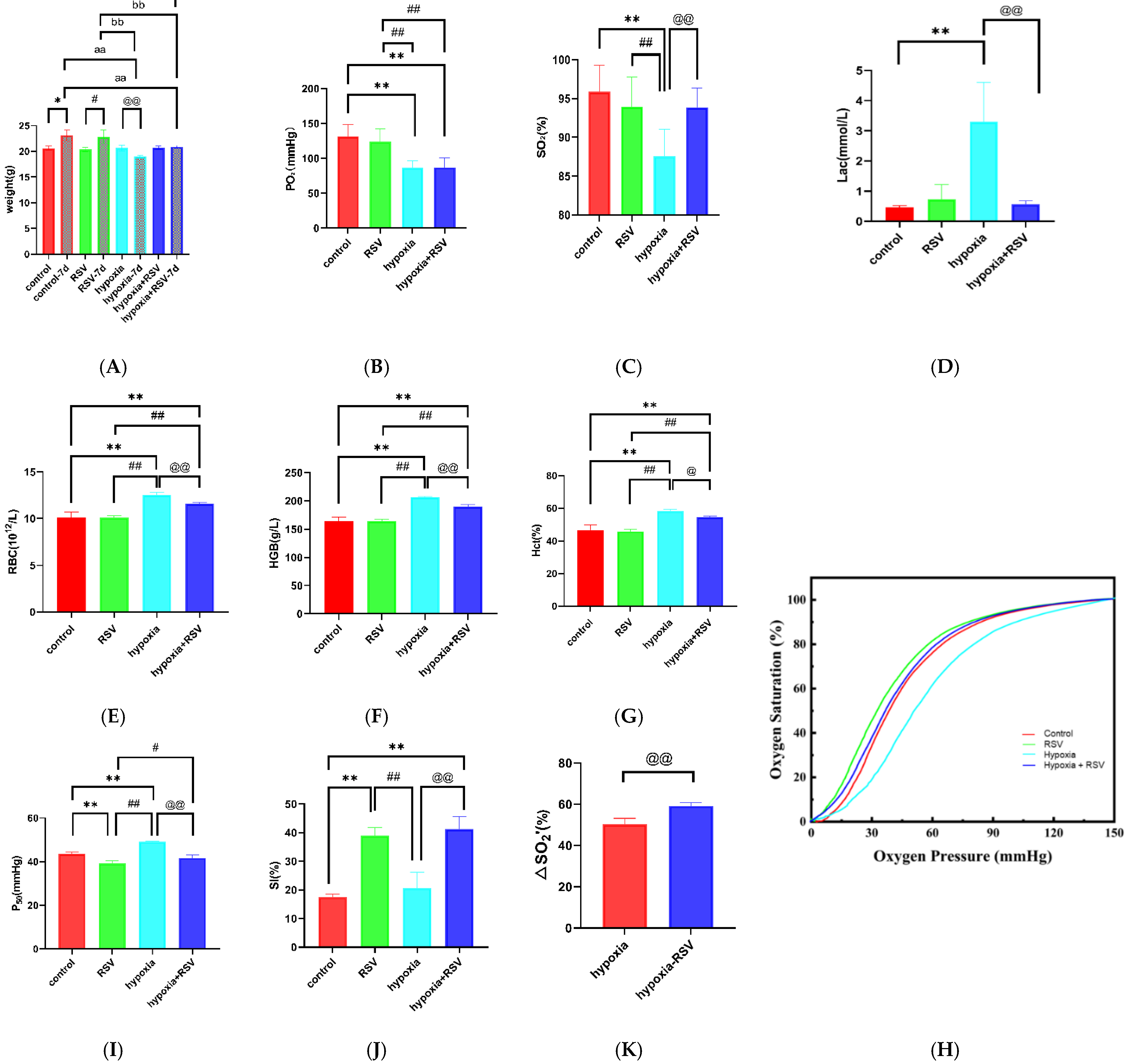
2.7.1. Resveratrol Inhibits Acute Hypoxia-Induced Weight Loss
2.7.2. Resveratrol Increased the Arterial Oxygen Saturation and Inhibited the Lactate Increase under Acute Hypoxia
2.7.3. Resveratrol Inhibits the Increases in RBCs and Hemoglobin during Acute Severe Hypoxia
2.7.4. Resveratrol Enhances the Oxygen Supply Efficiency In Vivo
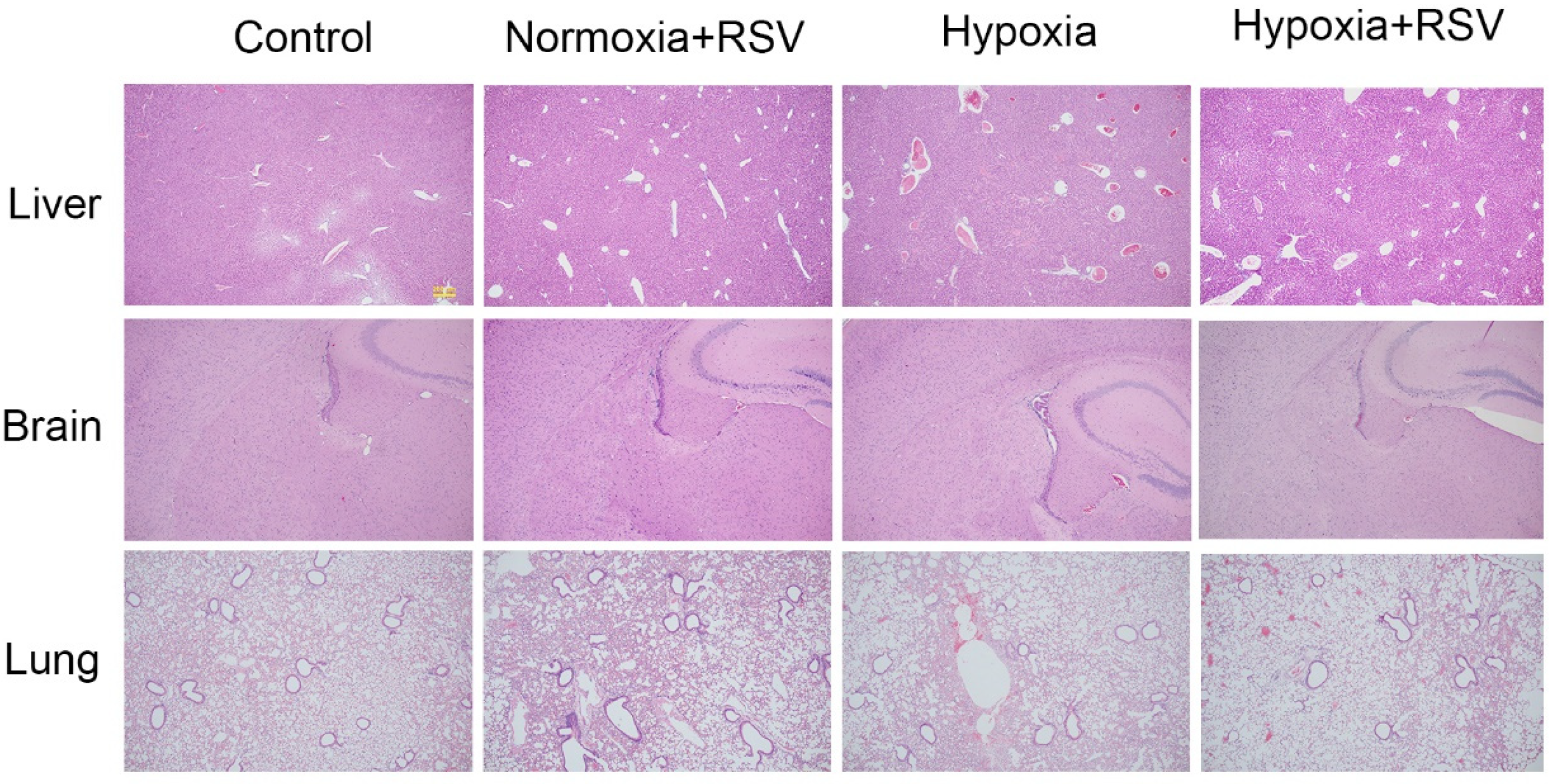
2.7.5. Resveratrol Protected the Organ Damage Caused by Acute Hypoxia
3. Discussion
3.1. Resveratrol Is an Allosteric Effector of HbA and Enhances the Oxygen Supply Efficiency of Blood Ex Vivo
3.2. Resveratrol Improves Adaption to Acute Severe Hypoxia
3.3. Enhancement of the Oxygen Supply Efficiency of Blood Improves Adaption to Acute Hypoxia
4. Methods and Materials
4.1. Ethical Considerations
4.2. Sample Preparation
4.2.1. Compounds
4.2.2. Hemoglobin of Adult and Rat Red Blood Cells
4.3. Surface Plasmon Resonance (SPR) Assays
4.4. Oxygen Dissociation Assay (ODA)
4.5. Molecular Docking
4.6. Thermal Stability Experiments
4.7. Oxygen Supply Efficiency Tests Ex Vivo
4.8. Acute Hypoxic Asphyxia Tolerance Detection
4.9. Anti-Acute Hypoxia Detection
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mairbaurl, H.; Weber, R.E. Oxygen transport by hemoglobin. Compr. Physiol. 2012, 2, 1463–1489. [Google Scholar]
- Aujla, H.; Woniak, M.; Kumar, T.; Murphy, G.J.; Investigators, R.; Rebecca, C.; Alison, D.; Renate, H.; Peter, S.; Rebecca, B. Rejuvenation of allogenic red cells: Benefits and risks. Vox Sang. 2018, 113, 509–529. [Google Scholar] [CrossRef] [Green Version]
- Lavrinenko, I.A.; Vashanov, G.A.; Cáceres, J.L.H.; Buchelnikov, A.S.; Nechipurenko, Y.D. A New Model of Hemoglobin Oxygenation. Entropy 2022, 24, 1214. [Google Scholar] [CrossRef]
- Srinivasan, A.J.; Morkane, C.; Martin, D.S.; Welsby, L.J. Should modulation of p50 be a therapeutic target in the critically ill? Expert Rev. Hematol. 2017, 10, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Zhao, M.; Feng, L.X.; Wang, P.Y.; Li, Y.M.; Li, W.H. Salidroside attenuates acute lung injury via inhibition of inflammatory cytokine production. Biomed Pharm. 2021, 142, 111949. [Google Scholar] [CrossRef]
- Chow, T.; Browne, V.; Heileson, H.L.; Wallace, D.; Anholm, J.; Green, S.M. Ginkgo biloba and acetazolamide prophylaxis for acute mountain sickness: A randomized, placebo-controlled trial. Arch. Intern. Med. 2005, 165, 296–301. [Google Scholar] [CrossRef] [Green Version]
- Grissom, C.K.; Roach, R.C.; Sarnquist, F.H.; Hackett, P.H. Acetazolamide in the treatment of acute mountain sickness: Clinical efficacy and effect on gas exchange. Ann. Intern. Med. 1992, 116, 461–465. [Google Scholar] [CrossRef]
- Luks, A.M.; Auerbach, P.S.; Freer, L.; Grissom, C.K.; Keyes, L.E.; McIntosh, S.E.; Rodway, G.W.; Schoene, R.B.; Zafren, K.; Hackett, P.H. Wilderness Medical Society Clinical Practice Guidelines for the Prevention and Treatment of Acute Altitude Illness: 2019 Update. Wilderness Environ. Med. 2019, 30, S3–S18. [Google Scholar] [CrossRef] [Green Version]
- Kunert, M.P.; Auerbach, P.S.; Freer, L.; Grissom, C.K.; Keyes, L.E.; McIntosh, S.E.; Rodway, G.W.; Schoene, R.B.; Zafren, K.; Hackett, P.H. Low-affinity hemoglobin increases tissue PO2 and decreases arteriolar diameter and flow in the rat cremaster muscle. Microvasc. Res. 1996, 52, 58–68. [Google Scholar] [CrossRef]
- Dufu, K.; Oksenberg, D. GBT440 reverses sickling of sickled red blood cells under hypoxic conditions in vitro. Hematol. Rep. 2018, 10, 7419. [Google Scholar] [CrossRef] [Green Version]
- Dufu, K.; Patel, M.; Oksenberg, D.; Cabrales, P. GBT440 improves red blood cell deformability and reduces viscosity of sickle cell blood under deoxygenated conditions. Clin. Hemorheol. Microcirc. 2018, 70, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Kilgore, K.S.; Shwartz, C.F.; Gallagher, M.A.; Steffen, R.P.; Mosca, R.S.; Bolling, S.F. RSR13, a synthetic allosteric modifier of hemoglobin, improves myocardial recovery following hypothermic cardiopulmonary bypass. Circulation 1999, 100, II351–II356. [Google Scholar] [CrossRef]
- Kleinberg, L.; Grossman, S.A.; Piantadosi, S.; Pearlman, J.; Engelhard, H.; Lesser, G.; Ruffer, J.; Gerber, M. Phase I trial to determine the safety, pharmacodynamics, and pharmacokinetics of RSR13, a novel radioenhancer, in newly diagnosed glioblastoma multiforme. J. Clin. Oncol. 1999, 17, 2593–2603. [Google Scholar] [CrossRef]
- Miyake, M.; Grinberg, O.Y.; Hou, H.; Steffen, R.P.; Elkadi, H.; Swartz, H.M. The effect of RSR13, a synthetic allosteric modifier of hemoglobin, on brain tissue pO2 (measured by EPR oximetry) following severe hemorrhagic shock in rats. Adv. Exp. Med. Biol. 2003, 530, 319–329. [Google Scholar]
- Woods, J.A.; Storey, C.J.; Babcock, E.E.; Malloy, C.R. Right-shifting the oxyhemoglobin dissociation curve with RSR13: Effects on high-energy phosphates and myocardial recovery after low-flow ischemia. J. Cardiovasc. Pharm. 1998, 31, 359–363. [Google Scholar] [CrossRef]
- Henry, E.R.; Metaferia, B.; Li, Q.; Harper, J.; Best, R.B.; Glass, K.E.; Cellmer, T.; Dunkelberger, E.B.; Conrey, A.; Thein, S.L.; et al. Treatment of sickle cell disease by increasing oxygen affinity of hemoglobin. Blood 2021, 138, 1172–1181. [Google Scholar] [CrossRef]
- Stea, B.; Shaw, E.; Pinter, T.; Hackman, J.; Craig, M.; May, J.; Steffen, R.P.; Suh, J.H. Efaproxiral red blood cell concentration predicts efficacy in patients with brain metastases. Br. J. Cancer 2006, 94, 1777–1784. [Google Scholar] [CrossRef]
- Ivy, C.M.; Wearing, O.H.; Natarajan, C.; Schweizer, R.M.; Gutierrez-Pinto, N.; Velotta, J.P.; Campbell-Staton, S.C.; Petersen, E.E.; Fago, A.; Cheviron, Z.A.; et al. Genetic variation in hemoglobin is associated with evolved changes in breathing in high-altitude deer mice. J. Exp. Biol. 2022, 225, jeb243595. [Google Scholar] [CrossRef]
- Yi, R.; Sandhu, J.; Zhao, S.; Lam, G.; Loganathan, D.; Morrissey, B. Detection of efaproxiral (RSR13) and its metabolites in equine by liquid chromatography tandem mass spectrometry. J. Mass Spectrom. 2014, 49, 57–67. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Alfred, M.A.; Anil Kumar, N.V.; Lombardi, N.; Cinquanta, L.; Iriti, M.; Varoni, E.M.; Gupta, G.; Chellappan, D.K.; et al. Current trends on resveratrol bioactivities to treat periodontitis. Food Biosci. 2021, 42, 101205. [Google Scholar] [CrossRef]
- Xia, N.; Daiber, A.; Forstermann, U.; Li, H. Antioxidant effects of resveratrol in the cardiovascular system. Br. J. Pharmacol. 2017, 174, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Ren, B.; Kwah, M.X.; Liu, C.; Ma, Z.; Shanmugam, M.K.; Ding, L.; Xiang, X.; Ho, P.C.; Wang, L.; Ong, P.S.; et al. Resveratrol for cancer therapy: Challenges and future perspectives. Cancer Lett. 2021, 515, 63–72. [Google Scholar] [CrossRef]
- Deng, B.; Liu, W.; Pu, L.; Wang, X.; Duan, R.; Wang, T.; Wang, Z.; Du, L.; Gao, Z.; Chen, Z. Quantitative Proteomics Reveals the Effects of Resveratrol on High-Altitude Polycythemia Treatment. Proteomics 2020, 20, e1900423. [Google Scholar] [CrossRef]
- Galtieri, A.; Tellone, E.; Ficarra, S.; Russo, A.; Bellocco, E.; Barreca, D.; Scatena, R.; Laganà, G.; Leuzzi, U.; Giardina, B. Resveratrol treatment induces redox stress in red blood cells: A possible role of caspase 3 in metabolism and anion transport. Biol. Chem. 2010, 391, 1057–1065. [Google Scholar] [CrossRef]
- Sticozzi, C.; Cervellati, F.; Muresan, X.M.; Cervellati, C.; Valacchi, G. Resveratrol prevents cigarette smoke-induced keratinocytes damage. Food Funct. 2014, 5, 2348–2356. [Google Scholar] [CrossRef]
- Tellone, E.; De Rosa, M.C.; Pirolli, D.; Russo, A.; Giardina, B.; Galtieri, A.; Ficarra, S. Molecular interactions of hemoglobin with resveratrol: Potential protective antioxidant role and metabolic adaptations of the erythrocyte. Biol. Chem. 2014, 395, 347–354. [Google Scholar] [CrossRef]
- de la Lastra, C.A.; Villegas, I. Resveratrol as an anti-inflammatory and anti-aging agent: Mechanisms and clinical implications. Mol. Nutr. Food Res. 2005, 49, 405–430. [Google Scholar] [CrossRef]
- Cao, B.; Qin, J.; Pan, B.; Qazi, I.H.; Ye, J.; Fang, Y.; Zhou, G. Oxidative Stress and Oocyte Cryopreservation: Recent Advances in Mitigation Strategies Involving Antioxidants. Cells 2022, 11, 3573. [Google Scholar] [CrossRef]
- Lu, Z.; Zhang, Y.; Liu, H.; Yuan, J.; Zheng, Z.; Zou, G. Transport of a cancer chemopreventive polyphenol, resveratrol: Interaction with serum albumin and hemoglobin. J Fluoresc. 2007, 17, 580–587. [Google Scholar] [CrossRef]
- Jiang, Y.L. Design, synthesis and spectroscopic studies of resveratrol aliphatic acid ligands of human serum albumin. Bioorgan. Med. Chem. 2008, 16, 6406–6414. [Google Scholar] [CrossRef]
- Nakagawa, A.; Ferrari, M.; Schleifer, G.; Cooper, M.K.; Liu, C.; Yu, B.; Berra, L.; Klings, E.S.; Safo, R.S.; Chen, Q.; et al. A Triazole Disulfide Compound Increases the Affinity of Hemoglobin for Oxygen and Reduces the Sickling of Human Sickle Cells. Mol. Pharm. 2018, 15, 1954–1963. [Google Scholar] [CrossRef]
- Okamoto, Y.; Sugisaki, S.; Suga, K.; Umakoshi, H. Development of Time-course Oxygen Binding Analysis for Hemoglobin-based Oxygen Carriers. Anal. Sci. 2017, 33, 953–956. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.P.; Siu, V.; Silva-Garcia, A.; Xu, Q.; Li, Z.; Oksenberg, D. Development and validation of an oxygen dissociation assay, a screening platform for discovering, and characterizing hemoglobin-oxygen affinity modifiers. Drug Des. Devel. Ther. 2018, 12, 1599–1607. [Google Scholar] [CrossRef] [Green Version]
- Hopp, M.T.; Rathod, D.C.; Winn, K.H.; Ambast, S.; Imhof, D. Novel insights into heme binding to hemoglobin. Biol. Chem. 2022, 403, 1055–1066. [Google Scholar] [CrossRef]
- Miele, A.E.; Bellelli, A.; Brunori, M. Hemoglobin allostery: New views on old players. J. Mol. Biol. 2013, 425, 1515–1526. [Google Scholar] [CrossRef]
- Okonjo, K.O. Bohr effect of native and chemically modified hemoglobins: Quantitative analyses based on the Wyman equation. Biophys. Chem. 2017, 226, 34–42. [Google Scholar] [CrossRef]
- Kavanagh, B.D.; Khandelwal, S.R.; Schmidt-Ullrich, R.K.; Roberts, J.D.; Shaw, E.G.; Pearlman, A.D.; Venitz, J.; Dusenbery, K.E.; Abraham, D.J.; Gerber, M.J. A phase I study of RSR13, a radiation-enhancing hemoglobin modifier: Tolerance of repeated intravenous doses and correlation of pharmacokinetics with pharmacodynamics. Int. J. Radiat. Oncol. Biol. Phys. 2001, 49, 1133–1139. [Google Scholar] [CrossRef]
- Schneider, M.A.; Linecker, M.; Fritsch, R.; Muehlematter, U.J.; Stocker, D.; Pestalozzi, B.; Samaras, P.; Jetter, A.; Kron, P.; Petrowsky, H.; et al. Phase Ib dose-escalation study of the hypoxia-modifier Myo-inositol trispyrophosphate in patients with hepatopancreatobiliary tumors. Nat. Commun. 2021, 12, 3807. [Google Scholar] [CrossRef]
- Kassa, T.; Wood, F.; Strader, M.B.; Alayash, A.I. Antisickling Drugs Targeting betaCys93 Reduce Iron Oxidation and Oxidative Changes in Sickle Cell Hemoglobin. Front. Physiol. 2019, 10, 931. [Google Scholar] [CrossRef] [Green Version]
- Woyke, S.; Mair, N.; Ortner, A.; Haller, T.; Ronzani, M.; Rugg, C.; Ströhle, M.; Wintersteiger, R.; Gatterer, H. Dose- and Sex-Dependent Changes in Hemoglobin Oxygen Affinity by the Micronutrient 5-Hydroxymethylfurfural and alpha-Ketoglutaric Acid. Nutrients 2021, 13, 3448. [Google Scholar] [CrossRef]
- Dominelli, P.B.; Wiggins, C.C.; Baker, S.E.; Shepherd, J.R.A.; Roberts, S.K.; Roy, T.K.; Curry, T.B.; Hoyer, J.D.; Oliveira, J.L.; Joyner, M.J. Influence of high affinity haemoglobin on the response to normoxic and hypoxic exercise. J. Physiol. 2020, 598, 1475–1490. [Google Scholar] [CrossRef]
- Signore, A.V.; Tift, M.S.; Hoffmann, F.G.; Schmitt, T.L.; Moriyama, H.; Storz, J.F. Evolved increases in hemoglobin-oxygen affinity and the Bohr effect coincided with the aquatic specialization of penguins. Proc. Natl. Acad. Sci. USA 2021, 118, e2023936118. [Google Scholar] [CrossRef]
- Jendroszek, A.; Malte, H.; Overgaard, C.B.; Beedholm, K.; Natarajan, C.; Weber, R.E.; Storz, J.F.; Fago, A. Allosteric mechanisms underlying the adaptive increase in hemoglobin-oxygen affinity of the bar-headed goose. J. Exp. Biol. 2018, 221, jeb185470. [Google Scholar] [CrossRef] [Green Version]
- Bellelli, A.; Tame, J. Hemoglobin allostery and pharmacology. Mol. Asp. Med. 2022, 84, 101037. [Google Scholar] [CrossRef]
- Tarasev, M.; Ferranti, M.; Herppich, A.; Hines, P. GBT1118, a voxelotor analog, protects red blood cells from damage during severe hypoxia. Am. J. Transl. Res. 2022, 14, 240–251. [Google Scholar]
- Yalcin, O.; Cabrales, P. Increased hemoglobin O2 affinity protects during acute hypoxia. Am. J. Physiol. -Heart Circ. Physiol. 2012, 303, H271–H281. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Li, X.; Liu, J.; Fan, X.; You, G.; Zhao, L.; Zhou, H.; Li, J.; Lei, H. Investigation of the differences between the Tibetan and Han populations in the hemoglobin-oxygen affinity of red blood cells and in the adaptation to high-altitude environments. Hematology 2018, 23, 309–313. [Google Scholar] [CrossRef] [Green Version]
- Lv, R.; Du, L.; Zhang, L.; Zhang, Z. Polydatin attenuates spinal cord injury in rats by inhibiting oxidative stress and microglia apoptosis via Nrf2/HO-1 pathway. Life Sci. 2019, 217, 119–127. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Y.; Wang, X.; Fan, X.; Zhang, Y.; Li, X.; Gong, Y.; Han, L. SIRT1 activation attenuates cardiac fibrosis by endothelial-to-mesenchymal transition. Biomed. Pharmacother. 2019, 118, 109227. [Google Scholar] [CrossRef]
- Xu, S.; Gao, Y.; Zhang, Q.; Wei, S.; Chen, Z.; Dai, X.; Zeng, Z.; Zhao, K. SIRT1/3 Activation by Resveratrol Attenuates Acute Kidney Injury in a Septic Rat Model. Oxidative Med. Cell. Longev. 2016, 2016, 7296092. [Google Scholar] [CrossRef] [Green Version]
- Abraham, S.K.; Khandelwal, N.; Hintzsche, H.; Stopper, H. Antigenotoxic effects of resveratrol: Assessment of in vitro and in vivo response. Mutagenesis 2016, 31, 27–33. [Google Scholar]
- Murdoch, D.R. High Life: A History of High Altitude Physiology and Medicine; Oxford University Press: Oxford, UK, 1999; p. 1631. [Google Scholar]
- West, J.B. Early history of high-altitude physiology. Ann. N. Y. Acad. Sci. 2016, 1365, 33–42. [Google Scholar] [CrossRef]
- Mangin, O. High oxygen affinity hemoglobins. Rev. Méd. Interne 2017, 38, 106–112. [Google Scholar] [CrossRef]
- Wagner, P.D.; Wagner, H.E.; Groves, B.M.; Cymerman, A.; Houston, C.S. Hemoglobin P50 during A Simulated Ascent of Mt. Everest, Operation Everest II. High Alt. Med. Biol. 2007, 8, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Sasagawa, K.; Imai, K.; Kobayashi, M. Influence of Allosteric Effectors and Temperature on Oxygen Binding Properties and the Bohr Effect of Bovine Hemoglobin. Zool. Sci. 2006, 23, 565–572. [Google Scholar] [CrossRef]
- Evans, B.A.; Ansari, A.K.; Kamyszek, R.W.; Salvagno, M.; Welsby, J.; Fuller, M.; Welsby, I. Modulation of red blood cell oxygen affinity with a novel allosteric modifier of hemoglobin is additive to the Bohr effect. Blood Cells Mol. Dis. 2021, 87, 102520. [Google Scholar] [CrossRef]
- Chu, Z.; Wang, Y.; You, G.; Wang, Q.; Ma, N.; Li, B.; Zhao, L.; Zhou, H. The P50 value detected by the oxygenation-dissociation analyser and blood gas analyser. Artif. Cells Nanomed. Biotechnol. 2020, 48, 867–874. [Google Scholar] [CrossRef]
- Song, T.; Yang, Y.; Wei, H.; Xie, X.; Lu, J.; Zeng, Q.; Peng, J.; Zhou, Y.; Jiang, S.; Peng, J. Zfp217 mediates m6A mRNA methylation to orchestrate transcriptional and post-transcriptional regulation to promote adipogenic differentiation. Nucleic Acids Res. 2019, 47, 6130–6144. [Google Scholar] [CrossRef] [Green Version]
- Pochron, M.; Siu, V.; Oksenberg, D.; Dufu, K. Central physiologic mechanisms which augment oxygen release (bohr effect and 2,3-dpg binding) are preserved in the presence of voxelotor at the therapeutic target of 30% hb modification:ps1522. HemaSphere 2019, 3, 701–702. [Google Scholar] [CrossRef]
- Wang, Z.L.; Gao, H.M.; Wang, S.; Zhang, M.; Chen, K.; Zhang, Y.Q.; Wang, H.D.; Han, B.Y.; Xu, L.L.; Song, T.Q.; et al. Dissection of the general two-step di-C-glycosylation pathway for the biosynthesis of (iso)schaftosides in higher plants. Proc. Natl. Acad. Sci. USA 2020, 117, 30816–30823. [Google Scholar] [CrossRef]
- Labute, P. LowModeMD—Implicit low-mode velocity filtering applied to conformational search of macrocycles and protein loops. J. Chem. Inf. Model. 2010, 50, 792–800. [Google Scholar] [CrossRef]
- Paoli, M.; Liddington, R.; Tame, J.; Wilkinson, A.; Dodson, G. Crystal structure of T state haemoglobin with oxygen bound at all four haems. J. Mol. Biol. 1996, 256, 775–792. [Google Scholar] [CrossRef]
- Volkamer, A.; Griewel, A.; Grombacher, T.; Rarey, M. Analyzing the topology of active sites: On the prediction of pockets and subpockets. J. Chem. Inf. Model. 2010, 50, 2041–2052. [Google Scholar] [CrossRef]
- Labute, P. The generalized Born/volume integral implicit solvent model: Estimation of the free energy of hydration using London dispersion instead of atomic surface area. J. Comput. Chem. 2008, 29, 1693–1698. [Google Scholar] [CrossRef]
- Li, Y.Y.; Cai, R.J.; Yang, J.Y.; Hendrickson, T.L.; Xiang, Y.; Javid, B. Clinically Relevant Mutations of Mycobacterial GatCAB Inform Regulation of Translational Fidelity. mBio 2021, 12, e0110021. [Google Scholar] [CrossRef]
- Wu, Y.; Li, C.; Xia, S.; Tian, X.; Kong, Y.; Wang, Z.; Gu, C.; Zhang, R.; Tu, C.; Xie, Y.; et al. Identification of Human Single-Domain Antibodies against SARS-CoV-2. Cell Host Microbe 2020, 27, 891–898. [Google Scholar] [CrossRef]
- Yonetani, T.; Park, S.; Tsuneshige, A.; Imai, K.; Kanaori, K. Global Allostery Model of Hemoglobin. J. Biol. Chem. 2002, 277, 34508–34520. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| Ka (association constant) | 3.97 × 102 |
| Kd (dissociation constant) | 1.93 × 10−2 |
| KD (dissociation equilibrium constant) | 4.86 × 10−5 M |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, Z.; Li, W.; You, G.; Chen, Y.; Qin, D.; Shu, P.; Wang, Y.; Wang, Y.; Zhao, L.; Zhou, H. Resveratrol, a New Allosteric Effector of Hemoglobin, Enhances Oxygen Supply Efficiency and Improves Adaption to Acute Severe Hypoxia. Molecules 2023, 28, 2050. https://doi.org/10.3390/molecules28052050
Chu Z, Li W, You G, Chen Y, Qin D, Shu P, Wang Y, Wang Y, Zhao L, Zhou H. Resveratrol, a New Allosteric Effector of Hemoglobin, Enhances Oxygen Supply Efficiency and Improves Adaption to Acute Severe Hypoxia. Molecules. 2023; 28(5):2050. https://doi.org/10.3390/molecules28052050
Chicago/Turabian StyleChu, Zongtang, Weidan Li, Guoxing You, Yuzhi Chen, Dong Qin, Peilin Shu, Yujing Wang, Ying Wang, Lian Zhao, and Hong Zhou. 2023. "Resveratrol, a New Allosteric Effector of Hemoglobin, Enhances Oxygen Supply Efficiency and Improves Adaption to Acute Severe Hypoxia" Molecules 28, no. 5: 2050. https://doi.org/10.3390/molecules28052050
APA StyleChu, Z., Li, W., You, G., Chen, Y., Qin, D., Shu, P., Wang, Y., Wang, Y., Zhao, L., & Zhou, H. (2023). Resveratrol, a New Allosteric Effector of Hemoglobin, Enhances Oxygen Supply Efficiency and Improves Adaption to Acute Severe Hypoxia. Molecules, 28(5), 2050. https://doi.org/10.3390/molecules28052050