


Two Onnamide Analogs from the Marine Sponge Theonella conica: Evaluation of Geometric Effects in the Polyene Systems on Biological Activity


Abstract
:1. Introduction
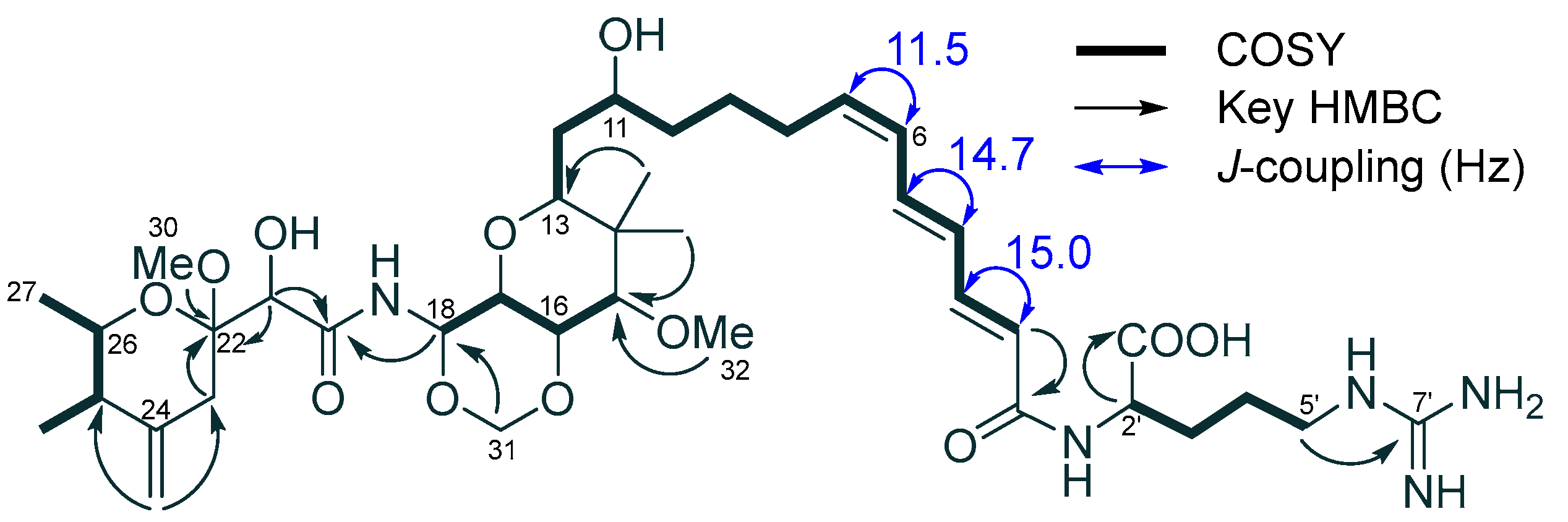
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Biological Material
3.3. Isolation
3.4. Cell Culture
3.5. Cytotoxic Test
3.6. Histone Modification Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bewley, C.A.; Faulkner, D.J. Lithistid Sponges: Star Performers or Hosts to the Stars. Angew. Chem. Int. Ed. 1998, 37, 2162–2178. [Google Scholar] [CrossRef]
- Sakemi, S.; Ichiba, T.; Kohmoto, S.; Saucy, G.; Higa, T. Isolation and Structure Elucidation of Onnamide A, a New Bioactive Metabolite of a Marine Sponge, Theonella sp. J. Am. Chem. Soc. 1988, 110, 4851–4853. [Google Scholar] [CrossRef]
- Matsunaga, S.; Fusetani, N.; Nakao, Y. Eight New Cytotoxic Metabolites Closely Related to Onnamide A from Two Marine Sponges of the Genus Theonella . Tetrahedron 1992, 48, 8369–8376. [Google Scholar] [CrossRef]
- Kobayashi, J.; Itagaki, F.; Shigemori, H.; Sasaki, T. Three New Onnamide Congeners from the Okinawan Marine Sponge Theonella sp. J. Nat. Prod. 1993, 56, 976–981. [Google Scholar] [CrossRef]
- Fusetani, N.; Sugawara, T.; Matsunaga, S. Bioactive Marine Metabolites. 41. Theopederins A-E, Potent Antitumor Metabolites from a Marine Sponge, Theonella sp. J. Org. Chem. 1992, 57, 3828–3832. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Matsunaga, S.; Fusetani, N.; Toh-e, A. Theopederins F-J: Five New Antifungal and Cytotoxic Metabolites from the Marine Sponge, Theonella Swinhoei. Tetrahedron 1999, 55, 13697–13702. [Google Scholar] [CrossRef]
- Cardani, C.; Fuganti, C.; Ghiringhelli, D.; Grasselli, P.; Pavan, M.; Valcurone, M.D. The Biosynthesis of Pederin. Tetrahedron Lett. 1973, 14, 2815–2818. [Google Scholar] [CrossRef]
- Piel, J.; Hui, D.; Wen, G.; Butzke, D.; Platzer, M.; Fusetani, N.; Matsunaga, S. Antitumor Polyketide Biosynthesis by an Uncultivated Bacterial Symbiont of the Marine Sponge Theonella Swinhoei. Proc. Natl. Acad. Sci. USA 2004, 101, 16222–16227. [Google Scholar] [CrossRef] [Green Version]
- Mosey, R.A.; Floreancig, P.E. Isolation, Biological Activity, Synthesis, and Medicinal Chemistry of the Pederin/Mycalamide Family of Natural Products. Nat. Prod. Rep. 2012, 29, 980. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin Modifications and Their Function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Pokholok, D.K.; Harbison, C.T.; Levine, S.; Cole, M.; Hannett, N.M.; Lee, T.I.; Bell, G.W.; Walker, K.; Rolfe, P.A.; Herbolsheimer, E.; et al. Genome-Wide Map of Nucleosome Acetylation and Methylation in Yeast. Cell 2005, 122, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portela, A.; Esteller, M. Epigenetic Modifications and Human Disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Hayashi-Takanaka, Y.; Kina, Y.; Nakamura, F.; Becking, L.E.; Nakao, Y.; Nagase, T.; Nozaki, N.; Kimura, H. Histone Modification Dynamics as Revealed by a Multicolor Immunofluorescence-Based Single-Cell Analysis. J. Cell. Sci. 2020, 133, jcs.243444. [Google Scholar] [CrossRef] [PubMed]
- Kupchan, S.M.; Britton, R.W.; Ziegler, M.F.; Sigel, C.W. Bruceantin, a New Potent Antileukemic Simaroubolide from Brucea Antidysenterica. J. Org. Chem. 1973, 38, 178–179. [Google Scholar] [CrossRef]
- Lindon, J.C.; Ferrige, A.G. Digitisation and Data Processing in Fourier Transform NMR. Prog. Nucl. Magn. Reson. Spectrosc. 1980, 14, 27–66. [Google Scholar] [CrossRef]
- Matsunaga, S.; Fujiki, H.; Sakata, D.; Fusetani, N. Calyculins E, F, G, and H, Additional Inhibitors of Protein Phosphatases 1 and 2a, from the Marine Sponge Discodermia Calyx. Tetrahedron 1991, 47, 2999–3006. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Nold, A.L.; Milburn, R.R.; Schindler, C.S.; Cole, K.P.; Yamaguchi, J. Total Synthesis of Marinomycins A–C and of Their Monomeric Counterparts Monomarinomycin A and Iso-Monomarinomycin A. J. Am. Chem. Soc. 2007, 129, 1760–1768. [Google Scholar] [CrossRef]
- Cao, R.; Wang, L.; Wang, H.; Xia, L.; Erdjument-Bromage, H.; Tempst, P.; Jones, R.S.; Zhang, Y. Role of Histone H3 Lysine 27 Methylation in Polycomb-Group Silencing. Science 2002, 298, 1039–1043. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Roberts, C.W.M. Targeting EZH2 in Cancer. Nat. Med. 2016, 22, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Sobel, R.E.; Cook, R.G.; Perry, C.A.; Annunziato, A.T.; Allis, C.D. Conservation of Deposition-Related Acetylation Sites in Newly Synthesized Histones H3 and H4. Proc. Natl. Acad. Sci. USA 1995, 92, 1237–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, S.; Li, G.; Son, J.; Xu, C.-F.; Margueron, R.; Neubert, T.A.; Reinberg, D. Phosphorylation of the PRC2 Component Ezh2 Is Cell Cycle-Regulated and up-Regulates Its Binding to NcRNA. Genes Dev. 2010, 24, 2615–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, P.; Tian, C.; An, L.; Nie, J.; Lu, K.; Xing, G.; Zhang, L.; He, F. Histone Methyltransferase Protein SETD2 Interacts with P53 and Selectively Regulates Its Downstream Genes. Cell Signal. 2008, 20, 1671–1678. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Calderas, L.; González-Barrios, R.; Herrera, L.A.; Cantú de León, D.; Soto-Reyes, E. The Role of the Histone Demethylase KDM4A in Cancer. Cancer Genet. 2015, 208, 215–224. [Google Scholar] [CrossRef]
- Cano, E.; Hazzalin, C.A.; Mahadevan, L.C. Anisomycin-Activated Protein Kinases P45 and P55 but Not Mitogen-Activated Protein Kinases ERK-1 and -2 Are Implicated in the Induction of c-Fos and c-Jun. Mol. Cell. Biol. 1994, 14, 7352–7362. [Google Scholar]
- Zinck, R.; Cahill, M.A.; Kracht, M.; Sachsenmaier, C.; Hipskind, R.A.; Nordheim, A. Protein Synthesis Inhibitors Reveal Differential Regulation of Mitogen-Activated Protein Kinase and Stress-Activated Protein Kinase Pathways That Converge on Elk-1. Mol. Cell. Biol. 1995, 15, 4930–4938. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-H.; Nishimura, S.; Matsunaga, S.; Fusetani, N.; Horinouchi, S.; Yoshida, M. Inhibition of Protein Synthesis and Activation of Stress-Activated Protein Kinases by Onnamide A and Theopederin B, Antitumor Marine Natural Products. Cancer Sci. 2005, 96, 357–364. [Google Scholar] [CrossRef]
- Iordanov, M.S.; Pribnow, D.; Magun, J.L.; Dinh, T.H.; Pearson, J.A.; Chen, S.L.; Magun, B.E. Ribotoxic Stress Response: Activation of the Stress-Activated Protein Kinase JNK1 by Inhibitors of the Peptidyl Transferase Reaction and by Sequence-Specific RNA Damage to the Alpha-Sarcin/Ricin Loop in the 28S RRNA. Mol. Cell. Biol. 1997, 17, 3373–3381. [Google Scholar] [CrossRef] [Green Version]
- Vind, A.C.; Genzor, A.V.; Bekker-Jensen, S. Ribosomal Stress-Surveillance: Three Pathways Is a Magic Number. Nucleic Acids Res. 2020, 48, 10648–10661. [Google Scholar] [CrossRef]
- Garreau de Loubresse, N.; Prokhorova, I.; Holtkamp, W.; Rodnina, M.V.; Yusupova, G.; Yusupov, M. Structural Basis for the Inhibition of the Eukaryotic Ribosome. Nature 2014, 513, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Schneider-Poetsch, T.; Eyler, D.E.; Jewett, J.C.; Bhat, S.; Rawal, V.H.; Green, R.; Liu, J.O. Inhibition of Eukaryotic Translation Elongation by the Antitumor Natural Product Mycalamide B. RNA 2011, 17, 1578–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepe, V.; Ummarino, R.; D’Auria, M.V.; Taglialatela-Scafati, O.; Marino, S.D.; D’Amore, C.; Renga, B.; Chini, M.G.; Bifulco, G.; Nakao, Y.; et al. Preliminary Structure-Activity Relationship on Theonellasterol, a New Chemotype of FXR Antagonist, from the Marine Sponge Theonella Swinhoei. Mar. Drugs 2012, 10, 2448–2466. [Google Scholar] [CrossRef] [PubMed]
- Sepe, V.; D’Amore, C.; Ummarino, R.; Renga, B.; D’Auria, M.V.; Novellino, E.; Sinisi, A.; Taglialatela-Scafati, O.; Nakao, Y.; Limongelli, V.; et al. Insights on Pregnane-X-Receptor Modulation. Natural and Semisynthetic Steroids from Theonella Marine Sponges. Eur. J. Med. Chem. 2014, 73, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, F.; Maejima, H.; Kawamura, M.; Arai, D.; Okino, T.; Zhao, M.; Ye, T.; Lee, J.; Chang, Y.-T.; Fusetani, N.; et al. Kakeromamide A, a New Cyclic Pentapeptide Inducing Astrocyte Differentiation Isolated from the Marine Cyanobacterium Moorea Bouillonii. Bioorg. Med. Chem. Lett. 2018, 28, 2206–2209. [Google Scholar] [CrossRef]
- Carpenter, A.E.; Jones, T.R.; Lamprecht, M.R.; Clarke, C.; Kang, I.H.; Friman, O.; Guertin, D.A.; Chang, J.H.; Lindquist, R.A.; Moffat, J.; et al. CellProfiler: Image Analysis Software for Identifying and Quantifying Cell Phenotypes. Genome Biol. 2006, 7, R100. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δC | δH Mult. (J in Hz) | COSY | HMBC |
|---|---|---|---|---|
| 1 | 168.5 | |||
| 2 | 120.8 | 5.74 d (11.6) | H-3 | C-1, C-4 |
| 3 | 141.7 | 6.46 t (11.6) | H-2, H-4 | C-1, C-5 |
| 4 | 128.4 | 7.45 dd (15.0, 11.6) | H-3, H-5 | |
| 5 | 142.4 | 6.43 dd (15.0, 10.7) | H-4, H-6 | |
| 6 | 131.9 | 6.24 dd (15.0, 10.7) | H-5, H-7 | |
| 7 | 140.5 | 5.95 dt (15.0, 6.9) | H-6, H-8 | C-5 |
| 8 | 34.1 | 2.13 m, 2.23 m | H-7, H-9 | |
| 9 | 31.6 | 1.30 m, 1.47 m | H-8, H-10 | |
| 10 | 37.0 | 1.29 m,1.43 m | H-11, H-9 | |
| 11 | 71.2 | 3.65 m | H-10, H-12 | |
| 12 | 37.5 | 1.54 m | H-11, H-13 | |
| 13 | 78.8 | 3.47 dd (8.9, 3.4) | H-12 | |
| 14 | 42.5 | |||
| 15 | 80.7 | 3.66 d (9.9) | H-16 | C-32 |
| 16 | 75.8 | 4.17 dd (9.9, 6.6) | H-15, H-17 | C-15, C-17, C-18, C-31 |
| 17 | 71.0 | 3.99 dd (9.4, 6.6) | H-16 | |
| 18 | 75.0 | 5.80 d (9.4) | H-17 | C-20, C-31 |
| 20 | 174.6 | |||
| 21 | 74.1 | 4.24 s | C-20, C-23, C-24 | |
| 22 | 101.5 | |||
| 23 | 34.9 | 2.41 d (14.3), 2.32 d (14.3) | C-22, C-24, C-25, C-29 | |
| 24 | 148.4 | |||
| 25 | 43.2 | 2.20 m | H-26, H-28 | C-24 |
| 26 | 71.0 | 3.88 qd (6.5, 2.5) | H-25, H-27 | C-25 |
| 27 | 18.3 | 1.18 d (6.5) | H-26, H-28 | C-25,26 |
| 28 | 12.5 | 0.97 d (7.0) | H-25, H-27 | C-24, C-25, C-26 |
| 29 | 110.2 | 4.80 brs, 4.64 brs | C-23, 25 | |
| 30 | 48.8 | 3.23 s | C-22 | |
| 31 | 87.8 | 5.21d (6.8), 4.79 d (6.8) | C-18 | |
| 32 | 62.1 | 3.56 s | C-15 | |
| 33 | 14.3 | 0.86 s | C-13, C-14, C-15, C-34 | |
| 34 | 23.7 | 1.00 s | C-13, C-14, C-15, C-33 | |
| 1′ | 178.6 | |||
| 2′ | 55.2 | 4.35 dd (7.3, 5.2) | H-3′ | C-1, C-1′, C-3′ |
| 3′ | 31.6 | 1.89 m, 1.74 m | H-2′, H-4′ | C-1′ |
| 4′ | 26.3 | 1.65 m | H-3′, H-5′ | C-3′, C-5′ |
| 5′ | 42.3 | 3.19 m, 3.26 m | H-4′ | C-7′ |
| 7′ | 158.7 |
| Position | δC | δH Mult. (J in Hz) | COSY | HMBC |
|---|---|---|---|---|
| 1 | 168.5 | |||
| 2 | 120.8 | 6.11 d (15.0) | H-3 | C-1, C-4 |
| 3 | 141.7 | 7.25 dd (15.0, 11.2) | H-2, H-4 | C-5 |
| 4 | 128.4 | 6.35 dd (14.7, 11.2) | H-3, H-5 | |
| 5 | 142.4 | 6.94 dd (14.7, 11.5) | H-4, H-6 | |
| 6 | 131.9 | 6.13 t (11.5) | H-5, H-7 | |
| 7 | 140.5 | 5.67 dt (11.5, 7.7) | H-6, H-8 | |
| 8 | 34.1 | 2.30 m | H-7, H-9 | |
| 9 | 31.6 | 1.50 m, 1.58 m | H-8, H-10 | |
| 10 | 37.0 | 1.32 m | H-11, H-9 | |
| 11 | 71.2 | 3.65 m | H-10, H-12 | |
| 12 | 37.5 | 1.54 m | H-11, H-13 | C-17 |
| 13 | 78.8 | 3.49 m | H-12 | |
| 14 | 42.5 | |||
| 15 | 80.7 | 3.66 d (9.8) | H-16 | C-32 |
| 16 | 75.8 | 4.17 dd (9.8, 6.6) | H-15, H-17 | C-15, C-17, C-18, C-31 |
| 17 | 71.0 | 3.98 dd (9.1, 6.6) | H-16 | |
| 18 | 75.0 | 5.83 d (9.1) | H-17 | C-20 |
| 20 | 174.6 | |||
| 21 | 74.1 | 4.24 s | C-20, 23, 24 | |
| 22 | 101.5 | |||
| 23 | 34.9 | 2.41 d (14.3), 2.32 d (14.3) | C-22, 24, 25, 29 | |
| 24 | 148.4 | |||
| 25 | 43.2 | 2.20 m | H-26, H-28 | |
| 26 | 71.0 | 3.87 qd (6.5, 2.6) | H-25, H-27 | |
| 27 | 18.3 | 1.17 d (6.5) | H-26 | C-25,26 |
| 28 | 12.5 | 0.97 d (7.1) | H-25 | C-24, 25, 26 |
| 29 | 110.2 | 4.79 s, 4.64 brs | C-23, 25 | |
| 30 | 48.8 | 3.24 s | C-22 | |
| 31 | 87.8 | 5.23d (6.9), 4.80 d (6.9) | C-18 | |
| 32 | 62.1 | 3.56 s | C-15 | |
| 33 | 14.3 | 0.86 s | C-13, 14, 15, 34 | |
| 34 | 23.7 | 1.00 s | C-13, 14, 15, 33 | |
| 1′ | 178.6 | |||
| 2′ | 55.2 | 4.38 dd (7.3, 5.2) | H-3′ | C-1′ |
| 3′ | 31.6 | 1.89 m, 1.75 m | H-2′, H-4′ | |
| 4′ | 26.3 | 1.64 m | H-3′, H-5′ | C-2′, 3′ |
| 5′ | 42.3 | 3.18 m, 3.22 m | H-4′ | C-7′ |
| 7′ | 158.7 |
| Cell Line | IC50 (µM) | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| HeLa | 0.17 | 0.15 | 0.066 | 0.038 | 0.057 | 0.54 |
| P388 | 1.8 | 4.8 | 0.62 | 0.31 | 0.57 | 5.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakamura, F.; Kimura, H.; Fusetani, N.; Nakao, Y. Two Onnamide Analogs from the Marine Sponge Theonella conica: Evaluation of Geometric Effects in the Polyene Systems on Biological Activity. Molecules 2023, 28, 2524. https://doi.org/10.3390/molecules28062524
Nakamura F, Kimura H, Fusetani N, Nakao Y. Two Onnamide Analogs from the Marine Sponge Theonella conica: Evaluation of Geometric Effects in the Polyene Systems on Biological Activity. Molecules. 2023; 28(6):2524. https://doi.org/10.3390/molecules28062524
Chicago/Turabian StyleNakamura, Fumiaki, Hiroshi Kimura, Nobuhiro Fusetani, and Yoichi Nakao. 2023. "Two Onnamide Analogs from the Marine Sponge Theonella conica: Evaluation of Geometric Effects in the Polyene Systems on Biological Activity" Molecules 28, no. 6: 2524. https://doi.org/10.3390/molecules28062524
APA StyleNakamura, F., Kimura, H., Fusetani, N., & Nakao, Y. (2023). Two Onnamide Analogs from the Marine Sponge Theonella conica: Evaluation of Geometric Effects in the Polyene Systems on Biological Activity. Molecules, 28(6), 2524. https://doi.org/10.3390/molecules28062524