

Plant Essential Oils as a Tool in the Control of Bovine Mastitis: An Update

Abstract
:1. Introduction
1.1. Bovine Mastitis
1.2. Economic Implications of Bovine Mastitis Worldwide
1.3. Treatment and Alternatives
2. Essential Oils
Antibacterial Effect
3. EOs and Their Components with Reported In Vitro Antibacterial Activity
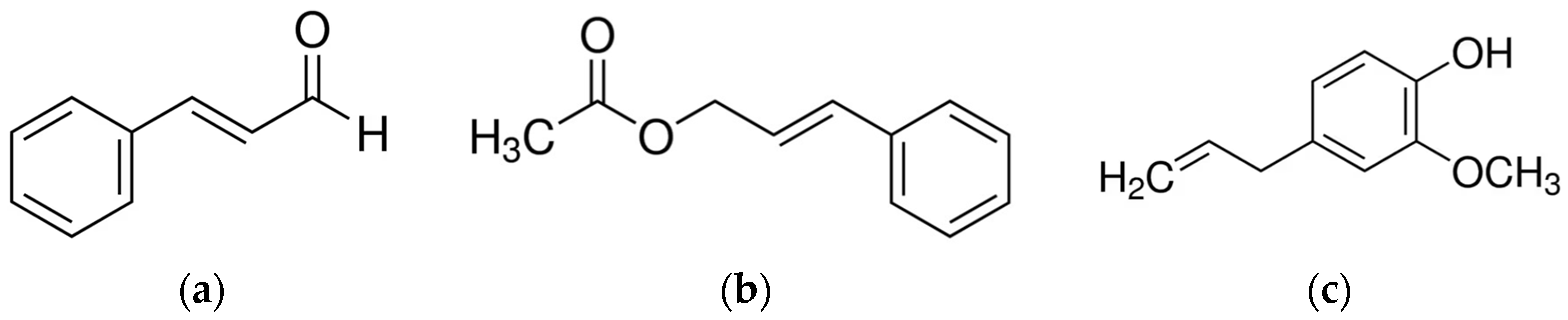
3.1. Cinnamon
3.2. Origanum
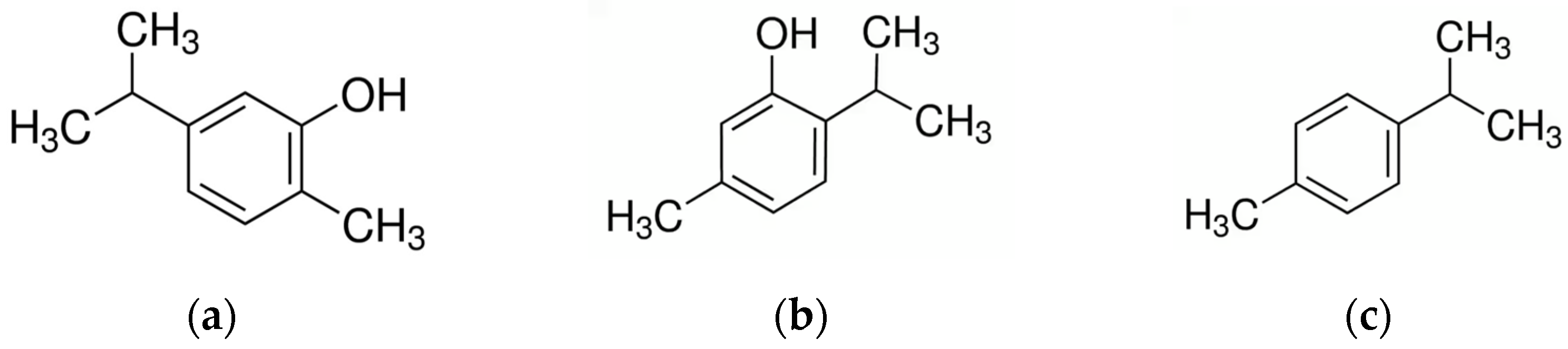
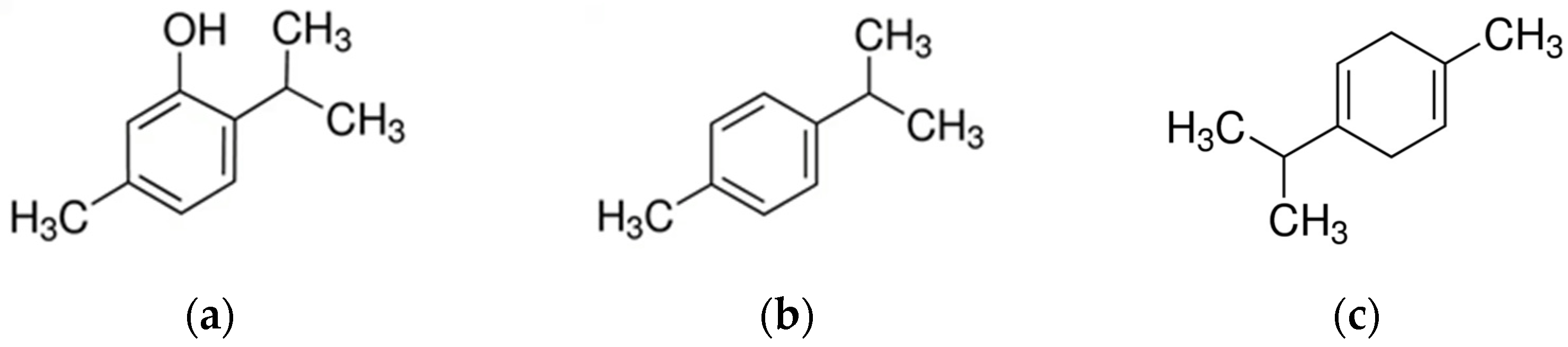
3.3. Thymus
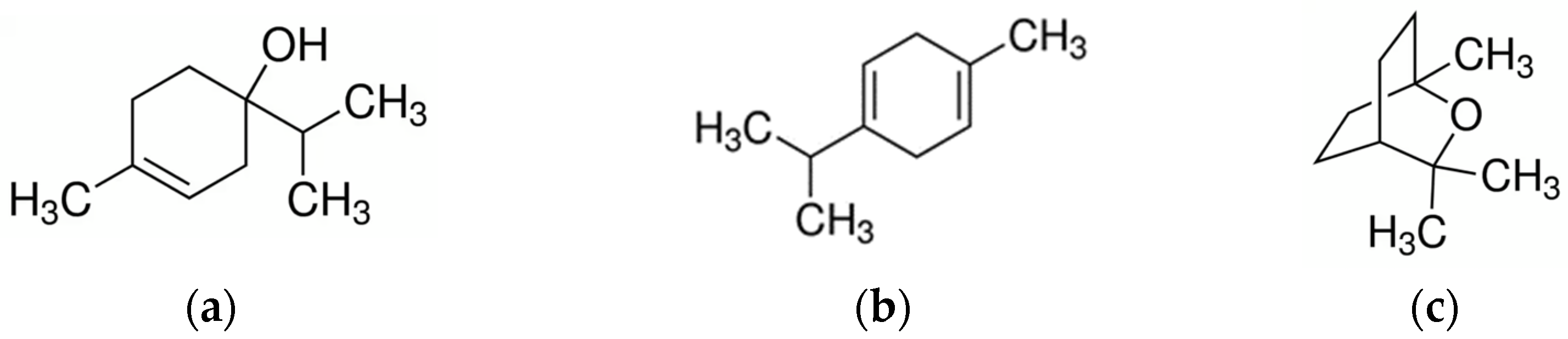
3.4. Melaleuca
3.5. Lavandula
3.6. Minthostachys verticillata
3.7. Essential Oils Derived from Other Plants
4. In Vivo Studies
5. Future Prospects and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gomes, F.; Henriques, M. Control of Bovine Mastitis: Old and Recent Therapeutic Approaches. Curr. Microbiol. 2016, 72, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Motaung, T.E.; Petrovski, K.R.; Petzer, I.-M.; Thekisoe, O.; Tsilo, T.J. Importance of Bovine Mastitis in Africa. Anim. Health Res. Rev. 2017, 18, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghamohammadi, M.; Haine, D.; Kelton, D.F.; Barkema, H.W.; Hogeveen, H.; Keefe, G.P.; Dufour, S. Herd-Level Mastitis-Associated Costs on Canadian Dairy Farms. Front. Vet. Sci. 2018, 5, 100. [Google Scholar] [CrossRef]
- Sathiyabarathi, M.; Jeyakumar, S.; Manimaran, A.; Jayaprakash, G.; Pushpadass, H.A.; Sivaram, M.; Ramesha, K.P.; Das, D.N.; Kataktalware, M.A.; Prakash, M.A.; et al. Infrared Thermography: A Potential Noninvasive Tool to Monitor Udder Health Status in Dairy Cows. Vet. World 2016, 9, 1075–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, B.; Pickering, A.C.; Rocha, L.S.; Aguilar, A.P.; Fabres-Klein, M.H.; de Oliveira Mendes, T.A.; Fitzgerald, J.R.; de Oliveira Barros Ribon, A. Diversity and Pathogenesis of Staphylococcus aureus from Bovine Mastitis: Current Understanding and Future Perspectives. BMC Vet. Res. 2022, 18, 115. [Google Scholar] [CrossRef]
- Gonçalves, J.L.; Kamphuis, C.; Martins, C.M.M.R.; Barreiro, J.R.; Tomazi, T.; Gameiro, A.H.; Hogeveen, H.; dos Santos, M.V. Bovine Subclinical Mastitis Reduces Milk Yield and Economic Return. Livest. Sci. 2018, 210, 25–32. [Google Scholar] [CrossRef]
- Sharun, K.; Dhama, K.; Tiwari, R.; Gugjoo, M.B.; Iqbal Yatoo, M.; Patel, S.K.; Pathak, M.; Karthik, K.; Khurana, S.K.; Singh, R.; et al. Advances in Therapeutic and Managemental Approaches of Bovine Mastitis: A Comprehensive Review. Vet. Q. 2021, 41, 107–136. [Google Scholar] [CrossRef]
- Abebe, R.; Hatiya, H.; Abera, M.; Megersa, B.; Asmare, K. Bovine Mastitis: Prevalence, Risk Factors and Isolation of Staphylococcus aureus in Dairy Herds at Hawassa Milk Shed, South Ethiopia. BMC Vet. Res. 2016, 12, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, J.G.; Babra, C.; Tiwari, H.; Williams, V.; De Wet, S.; Gibson, J.; Paxman, A.; Morgan, E.; Costantino, P.; Sunagar, R. Trends in Therapeutic and Prevention Strategies for Management of Bovine Mastitis: An Overview. J. Vaccines Vaccin. 2013, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lakew, B.T.; Fayera, T.; Ali, Y.M. Risk Factors for Bovine Mastitis with the Isolation and Identification of Streptococcus Agalactiae from Farms in and around Haramaya District, Eastern Ethiopia. Trop. Anim. Health Prod. 2019, 51, 1507–1513. [Google Scholar] [CrossRef]
- Kibebew, K. Bovine Mastitis: A Review of Causes and Epidemiological Point of View. J. Biol. Agric. Healthc. 2017, 7, 1–14. [Google Scholar]
- Bradley, A.J. Bovine Mastitis: An Evolving Disease. Vet. J. 2002, 164, 116–128. [Google Scholar] [CrossRef]
- Bogni, C.; Odierno, L.; Raspanti, C.; Giraudo, J.; Larriestra, A.; Reinoso, E.; Lasagno, M.; Ferrari, M.; Ducrós, E.; Frigerio, C. War against Mastitis: Current Concepts on Controlling Bovine Mastitis Pathogens. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances; FORMATEX: Badajoz, Spain, 2011; pp. 483–494. [Google Scholar]
- Reinoso, E.B. Bovine Mastitis Caused by Streptococcus Uberis. Virulence Factors and Biofilm. J. Microb. Biochem. Technol. 2017, 9, 5. [Google Scholar]
- Gilmore, K.S.; Srinivas, P.; Akins, D.R.; Hatter, K.L.; Gilmore, M.S. Growth, Development, and Gene Expression in a Persistent Streptococcus Gordonii Biofilm. Infect. Immun. 2003, 71, 4759–4766. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, I.W. The Biofilm Matrix—An Immobilized but Dynamic Microbial Environment. Trends Microbiol. 2001, 9, 222–227. [Google Scholar] [CrossRef]
- Melchior, M.B.; Vaarkamp, H.; Fink-Gremmels, J. Biofilms: A Role in Recurrent Mastitis Infections? Vet. J. 2006, 171, 398–407. [Google Scholar] [CrossRef]
- Hussein, H.A.; El-Razik, K.A.E.-H.A.; Gomaa, A.M.; Elbayoumy, M.K.; Abdelrahman, K.A.; Hosein, H.I. Milk Amyloid A as a Biomarker for Diagnosis of Subclinical Mastitis in Cattle. Vet. World 2018, 11, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, A.; Imran, M. Diagnosis of Bovine Mastitis: From Laboratory to Farm. Trop. Anim. Health Prod. 2018, 50, 1193–1202. [Google Scholar] [CrossRef]
- Mahmmod, Y. The Future of PCR Technologies in Diagnosis of Bovine Mastitis Pathogens. Adv. Dairy Res. 2013, 2, e106. [Google Scholar] [CrossRef]
- Deb, R.; Kumar, A.; Chakraborty, S.; Verma, A.K.; Tiwari, R.; Dhama, K.; Singh, U.; Kumar, S. Trends in Diagnosis and Control of Bovine Mastitis: A Review. Pak. J. Biol. Sci. 2013, 16, 1653–1661. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Freitas, P.P.; Bexiga, R. Technological Advances in Bovine Mastitis Diagnosis: An Overview. J. Vet. Diagn. Investig. 2015, 27, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sayed, A.; Kamel, M. Bovine Mastitis Prevention and Control in the Post-Antibiotic Era. Trop. Anim. Health Prod. 2021, 53, 236. [Google Scholar] [CrossRef]
- Sharma, N.; Jeong, D.K. Stem Cell Research: A Novel Boulevard towards Improved Bovine Mastitis Management. Int. J. Biol. Sci. 2013, 9, 818–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banga, C.B.; Neser, F.W.; Garrick, D.J. The Economic Value of Somatic Cell Count in South African Holstein and Jersey Cattle. S. Afr. J. Anim. Sci. 2014, 44, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Khodaei Motlagh, M.; Kazemi, M.; Khaltabadi, F.A.H.; Yahyaei, M.; Rezaei, M.; De Rensis, F.; Taddei, S. Antibacterial Effect of Medicinal Plant Essence (Thymus Vulgaris) on Major Bacterial Mastitis Pathogen in Vitro. Int. J. Adv. Biol. Biomed. Res. 2014, 2, 286–294. [Google Scholar]
- Kabelitz, T.; Aubry, E.; van Vorst, K.; Amon, T.; Fulde, M. The Role of Streptococcus Spp. in Bovine Mastitis. Microorganisms 2021, 9, 1497. [Google Scholar] [CrossRef]
- Cheng, W.N.; Han, S.G. Bovine Mastitis: Risk Factors, Therapeutic Strategies, and Alternative Treatments—A Review. Asian-Australas. J. Anim. Sci. 2020, 33, 1699–1713. [Google Scholar] [CrossRef] [PubMed]
- van den Borne, B.H.P.; van Schaik, G.; Lam, T.J.G.M.; Nielen, M. Variation in Herd Level Mastitis Indicators between Primi- and Multiparae in Dutch Dairy Herds. Prev. Vet. Med. 2010, 96, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.K.; Thombare, N.N.; Mondal, B. Subclinical Mastitis in Dairy Animals: Incidence, Economics, and Predisposing Factors. Sci. World J. 2014, 2014, 523984. [Google Scholar] [CrossRef]
- Halasa, T.; Nielen, M.; De Roos, A.P.W.; Van Hoorne, R.; de Jong, G.; Lam, T.J.G.M.; van Werven, T.; Hogeveen, H. Production Loss Due to New Subclinical Mastitis in Dutch Dairy Cows Estimated with a Test-Day Model. J. Dairy Sci. 2009, 92, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Qayyum, A.; Khan, J.A.; Hussain, R.; Avais, M.; Ahmad, N.; Khan, M.S. Investigation of Milk and Blood Serum Biochemical Profile as an Indicator of Sub-Clinical Mastitis in Cholistani Cattle. Pak. Vet. J. 2016, 36, 275–279. [Google Scholar]
- Breen, J. The Importance of Teat Disinfection in Mastitis Control. Livestock 2019, 24, 122–128. [Google Scholar] [CrossRef]
- Down, P.M.; Bradley, A.J.; Breen, J.E.; Hudson, C.D.; Green, M.J. Current Management Practices and Interventions Prioritised as Part of a Nationwide Mastitis Control Plan. Vet. Rec. 2016, 178, 449. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.K.; Paul, S.; Hossain, M.M.; Islam, M.R.; Alam, M.G.S. Bovine Mastitis and Its Therapeutic Strategy Doing Antibiotic Sensitivity Test. Austin. J. Vet. Sci. Anim. Husb. 2017, 4, 1030. [Google Scholar]
- Biggs, A. Update on Dry Cow Therapy 1. Antibiotic v Non-Antibiotic Approaches. Practice 2017, 39, 328–333. [Google Scholar] [CrossRef]
- Du Preez, J.H. Bovine Mastitis Therapy and Why It Fails. J. S. Afr. Vet. Assoc. 2000, 71, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.G.B.; Blagitz, M.G.; Souza, F.N.; Sanchez, E.M.R.; Batista, C.F.; Bertagnon, H.G.; Diniz, S.A.; Silva, M.X.; Libera, A.M.M.P.D. Profile of Immunoglobulins, Clinical and Bacteriological Cure after Different Treatment Routes of Clinical Bovine Mastitis. Arq. Bras. Med. Veterinária E Zootec. 2018, 70, 1141–1149. [Google Scholar] [CrossRef] [Green Version]
- Hillerton, J.E.; Kliem, K.E. Effective Treatment of Streptococcus Uberis Clinical Mastitis to Minimize the Use of Antibiotics. J. Dairy Sci. 2002, 85, 1009–1014. [Google Scholar] [CrossRef]
- Fair, R.J.; Tor, Y. Antibiotics and Bacterial Resistance in the 21st Century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [Green Version]
- Regulation (EU) 2019/6 of the European Parliament and of the Council of 11 December 2018 on Veterinary Medicinal Products and Repealing Directive 2001/82/EC (Text with EEA Relevance). Off. J. Eur. Union 2018, L4, 43–167.
- Caneschi, A.; Bardhi, A.; Barbarossa, A.; Zaghini, A. The Use of Antibiotics and Antimicrobial Resistance in Veterinary Medicine, a Complex Phenomenon: A Narrative Review. Antibiotics 2023, 12, 487. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization No Time to Wait: Securing the Future from Drug-Resistant Infections. Available online: https://www.who.int/publications-detail-redirect/no-time-to-wait-securing-the-future-from-drug-resistant-infections (accessed on 8 February 2023).
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Government of the United Kingdom: London, UK, 2016.
- Pulingam, T.; Parumasivam, T.; Gazzali, A.M.; Sulaiman, A.M.; Chee, J.Y.; Lakshmanan, M.; Chin, C.F.; Sudesh, K. Antimicrobial Resistance: Prevalence, Economic Burden, Mechanisms of Resistance and Strategies to Overcome. Eur. J. Pharm. Sci. 2022, 170, 106103. [Google Scholar] [CrossRef] [PubMed]
- Subirats, J.; Domingues, A.; Topp, E. Does Dietary Consumption of Antibiotics by Humans Promote Antibiotic Resistance in the Gut Microbiome? J. Food Prot. 2019, 82, 1636–1642. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-T.; Ke, C.-Y.; Wu, W.-T.; Lee, R.-P.; Tseng, Y.-H. Effective Treatment of Bovine Mastitis with Intramammary Infusion of Angelica Dahurica and Rheum Officinale Extracts. Evid.-Based Complement. Altern. Med. ECAM 2019, 2019, 7242705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orellano, M.S.; Isaac, P.; Breser, M.L.; Bohl, L.P.; Conesa, A.; Falcone, R.D.; Porporatto, C. Chitosan Nanoparticles Enhance the Antibacterial Activity of the Native Polymer against Bovine Mastitis Pathogens. Carbohydr. Polym. 2019, 213, 1–9. [Google Scholar] [CrossRef]
- Varela-Ortiz, D.F.; Barboza-Corona, J.E.; González-Marrero, J.; León-Galván, M.F.; Valencia-Posadas, M.; Lechuga-Arana, A.A.; Sánchez-Felipe, C.G.; Ledezma-García, F.; Gutiérrez-Chávez, A.J. Antibiotic Susceptibility of Staphylococcus aureus Isolated from Subclinical Bovine Mastitis Cases and in Vitro Efficacy of Bacteriophage. Vet. Res. Commun. 2018, 42, 243–250. [Google Scholar] [CrossRef]
- Langer, M.N.; Blodkamp, S.; Bayerbach, M.; Feßler, A.T.; de Buhr, N.; Gutsmann, T.; Kreienbrock, L.; Schwarz, S.; von Köckritz-Blickwede, M. Testing Cathelicidin Susceptibility of Bacterial Mastitis Isolates: Technical Challenges and Data Output for Clinical Isolates. Vet. Microbiol. 2017, 210, 107–115. [Google Scholar] [CrossRef]
- Pellegrino, M.; Berardo, N.; Giraudo, J.; Nader-Macías, M.E.F.; Bogni, C. Bovine Mastitis Prevention: Humoral and Cellular Response of Dairy Cows Inoculated with Lactic Acid Bacteria at the Dry-off Period. Benef. Microbes 2017, 8, 589–596. [Google Scholar] [CrossRef]
- Almeida, R.A.; Kerro-Dego, O.; Prado, M.E.; Headrick, S.I.; Lewis, M.J.; Siebert, L.J.; Pighetti, G.M.; Oliver, S.P. Protective Effect of Anti-SUAM Antibodies on Streptococcus Uberis Mastitis. Vet. Res. 2015, 46, 133. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Wang, L.; Chen, W. Oxytropis Glabra DC. Inhibits Biofilm Formation of Staphylococcus Epidermidis by Down-Regulating Ica Operon Expression. Curr. Microbiol. 2020, 77, 1167–1173. [Google Scholar] [CrossRef]
- Kher, M.N.; Sheth, N.R.; Bhatt, V.D. In Vitro Antibacterial Evaluation of Terminalia Chebula as an Alternative of Antibiotics against Bovine Subclinical Mastitis. Anim. Biotechnol. 2019, 30, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.; Rana, A.; Jaitak, V. Essential Oils: An Impending Substitute of Synthetic Antimicrobial Agents to Overcome Antimicrobial Resistance. Curr. Drug Targets 2019, 20, 605–624. [Google Scholar] [CrossRef] [PubMed]
- Jubair, N.; Rajagopal, M.; Chinnappan, S.; Abdullah, N.B.; Fatima, A. Review on the Antibacterial Mechanism of Plant-Derived Compounds against Multidrug-Resistant Bacteria (MDR). Evid.-Based Complement. Altern. Med. ECAM 2021, 2021, 3663315. [Google Scholar] [CrossRef]
- ISO 9235:2021(En), Aromatic Natural Raw Materials—Vocabulary. Available online: https://www.iso.org/obp/ui#iso:std:iso:9235:ed-3:v1:en (accessed on 10 February 2023).
- Ríos, J.-L. Chapter 1—Essential Oils: What They Are and How the Terms Are Used and Defined. In Essential Oils in Food Preservation, Flavor and Safety; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 3–10. ISBN 978-0-12-416641-7. [Google Scholar]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Maffei, M.E.; Gertsch, J.; Appendino, G. Plant Volatiles: Production, Function and Pharmacology. Nat. Prod. Rep. 2011, 28, 1359–1380. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, M.; Mehdizadeh, L. Chapter 13—Chemistry of Essential Oils and Factors Influencing Their Constituents. In Soft Chemistry and Food Fermentation; Grumezescu, A.M., Holban, A.M., Eds.; Handbook of Food Bioengineering; Academic Press: Cambridge, MA, USA, 2017; pp. 379–419. ISBN 978-0-12-811412-4. [Google Scholar]
- Panda, S.K.; Daemen, M.; Sahoo, G.; Luyten, W. Essential Oils as Novel Anthelmintic Drug Candidates. Molecules 2022, 27, 8327. [Google Scholar] [CrossRef]
- Buldain, D.; Gortari Castillo, L.; Buchamer, A.V.; Bandoni, A.; Marchetti, L.; Mestorino, N. In Vitro Synergistic Interaction between Melaleuca Armillaris Essential Oil and Erythromycin against Staphylococcus aureus Isolated from Dairy Cows. Front. Vet. Sci. 2022, 9, 1005616. [Google Scholar] [CrossRef]
- Arbab, S.; Ullah, H.; Bano, I.; Li, K.; Ul Hassan, I.; Wang, W.; Qadeer, A.; Zhang, J. Evaluation of in Vitro Antibacterial Effect of Essential Oil and Some Herbal Plant Extract Used against Mastitis Pathogens. Vet. Med. Sci. 2022, 8, 2655–2661. [Google Scholar] [CrossRef]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A Comprehensive Review of the Antibacterial, Antifungal and Antiviral Potential of Essential Oils and Their Chemical Constituents against Drug-Resistant Microbial Pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef]
- Hammer, K.A.; Carson, C.F. Antibacterial and Antifungal Activities of Essential Oils. In Lipids and Essential Oils as Antimicrobial Agents; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011; pp. 255–306. ISBN 978-0-470-97662-3. [Google Scholar]
- Oussalah, M.; Caillet, S.; Lacroix, M. Mechanism of Action of Spanish Oregano, Chinese Cinnamon, and Savory Essential Oils against Cell Membranes and Walls of Escherichia coli O157:H7 and Listeria Monocytogenes. J. Food Prot. 2006, 69, 1046–1055. [Google Scholar] [CrossRef]
- Turina, A.d.V.; Nolan, M.V.; Zygadlo, J.A.; Perillo, M.A. Natural Terpenes: Self-Assembly and Membrane Partitioning. Biophys. Chem. 2006, 122, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Saad, N.Y.; Muller, C.D.; Lobstein, A. Major Bioactivities and Mechanism of Action of Essential Oils and Their Components. Flavour Fragr. J. 2013, 28, 269–279. [Google Scholar] [CrossRef]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pagès, J.-M.; Amaral, L.; Bolla, J.-M. Geraniol Restores Antibiotic Activities against Multidrug-Resistant Isolates from Gram-Negative Species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, J.S. Mitochondrial Membrane Permeabilization: The Sine qua Non for Cell Death. BioEssays 2006, 28, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Petretto, G.L.; Chessa, M.; Piana, A.; Masia, M.D.; Foddai, M.; Mangano, G.; Culeddu, N.; Afifi, F.U.; Pintore, G. Chemical and Biological Study on the Essential Oil of Artemisia caerulescens L. ssp. Densiflora (Viv.). Nat. Prod. Res. 2013, 27, 1709–1715. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Przychodna, M.; Sopata, S.; Bodalska, A.; Fecka, I. Thymol and Thyme Essential Oil-New Insights into Selected Therapeutic Applications. Molecules 2020, 25, 4125. [Google Scholar] [CrossRef]
- Rani, S.; Singh, H.; Ram, C. Efficacy and Mechanism of Carvacrol with Octanoic Acid against Mastitis Causing Multi-Drug-Resistant Pathogens. Braz. J. Microbiol. 2022, 53, 385–399. [Google Scholar] [CrossRef]
- Corona-Gómez, L.; Hernández-Andrade, L.; Mendoza-Elvira, S.; Suazo, F.M.; Ricardo-González, D.I.; Quintanar-Guerrero, D. In Vitro Antimicrobial Effect of Essential Tea Tree Oil(Melaleuca Alternifolia), Thymol, and Carvacrol on Microorganisms Isolated from Cases of Bovine Clinical Mastitis. Int. J. Vet. Sci. Med. 2022, 10, 72–79. [Google Scholar] [CrossRef]
- Dal Pozzo, M.; Santurio, D.F.; Rossatto, L.; Vargas, A.C.; Alves, S.H.; Loreto, E.S.; Viegas, J. Activity of Essential Oils from Spices against Staphylococcus Spp. Isolated from Bovine Mastitis. Arq. Bras. Med. Veterinária E Zootec. 2011, 63, 1229–1232. [Google Scholar] [CrossRef] [Green Version]
- Budri, P.E.; Silva, N.C.C.; Bonsaglia, E.C.R.; Fernandes Júnior, A.; Araújo Júnior, J.P.; Doyama, J.T.; Gonçalves, J.L.; Santos, M.V.; Fitzgerald-Hughes, D.; Rall, V.L.M. Effect of Essential Oils of Syzygium Aromaticum and Cinnamomum Zeylanicum and Their Major Components on Biofilm Production in Staphylococcus aureus Strains Isolated from Milk of Cows with Mastitis. J. Dairy Sci. 2015, 98, 5899–5904. [Google Scholar] [CrossRef]
- Dal Pozzo, M.; Loreto, É.S.; Santurio, D.F.; Alves, S.H.; Rossatto, L.; de Vargas, A.C.; Viegas, J.; da Costa, M.M. Antibacterial Activity of Essential Oil of Cinnamon and Trans-Cinnamaldehyde against Staphylococcus Spp. Isolated from Clinical Mastitis of Cattle and Goats. Acta Sci. Vet. 2012, 40, 1–5. [Google Scholar]
- Abd El-Aziz, N.K.; Ammar, A.M.; El-Naenaeey, E.Y.M.; El Damaty, H.M.; Elazazy, A.A.; Hefny, A.A.; Shaker, A.; Eldesoukey, I.E. Antimicrobial and Antibiofilm Potentials of Cinnamon Oil and Silver Nanoparticles against Streptococcus Agalactiae Isolated from Bovine Mastitis: New Avenues for Countering Resistance. BMC Vet. Res. 2021, 17, 136. [Google Scholar] [CrossRef] [PubMed]
- Benbelkacem, I.; Selles, S.M.A.; Aissi, M.; Khaldi, F.; Ghazi, K. In Vitro Assessment of Antifungal and Antistaphylococcal Activities of Cinnamomum Aromaticum Essential Oil against Subclinical Mastitis Pathogens. Veterinaria 2019, 68, 31. [Google Scholar]
- Zhu, H.; Du, M.; Fox, L.; Zhu, M.-J. Bactericidal Effects of Cinnamon Cassia Oil against Bovine Mastitis Bacterial Pathogens. Food Control 2016, 66, 291–299. [Google Scholar] [CrossRef]
- Szweda, P.; Zalewska, M.; Pilch, J.; Kot, B.; Milewski, S. Essential Oils as Potential Anti-Staphylococcal Agents. Acta Vet. Beogr. 2018, 68, 95–107. [Google Scholar]
- Barreiros, Y.; de Meneses, A.C.; Alves, J.L.F.; Mumbach, G.D.; Ferreira, F.A.; Machado, R.A.F.; Bolzan, A.; de Araujo, P.H.H. Xanthan Gum-Based Film-Forming Suspension Containing Essential Oils: Production and in Vitro Antimicrobial Activity Evaluation against Mastitis-Causing Microorganisms. LWT 2022, 153, 112470. [Google Scholar] [CrossRef]
- Grzesiak, B.; Głowacka, A.; Krukowski, H.; Lisowski, A.; Lassa, H.; Sienkiewicz, M. The In Vitro Efficacy of Essential Oils and Antifungal Drugs Against Prototheca Zopfii. Mycopathologia 2016, 181, 609–615. [Google Scholar] [CrossRef]
- Pașca, C.; Mărghitaș, L.A.; Dezmirean, D.; Bobiș, O.; Bonta, V.; Mărgăoan, R.; Chirilă, F.; Fit, N. The Assessment of the Antibacterial Activity of Some Plant Extracts on Normal and Pathogenic Microflora from Milk. Sci. Pap. Anim. Sci. Biotechnol. 2015, 48, 166–172. [Google Scholar]
- Noori, S.; Rahchamani, R.; Bayat Kohsar, J.; Bahri Binabaj, F. Antibacterial Effect of Lavandula Stoechas and Origanum Majorana Essential Oils against Staphylococcus aureus, Streptococcus Agalactiae, and Escherichia coli. Iran. J. Vet. Sci. Technol. 2019, 11, 35–66. [Google Scholar]
- Tortorano, A.M.; Prigitano, A.; Dho, G.; Piccinini, R.; Daprà, V.; Viviani, M.A. In Vitro Activity of Conventional Antifungal Drugs and Natural Essences against the Yeast-like Alga Prototheca. J. Antimicrob. Chemother. 2008, 61, 1312–1314. [Google Scholar] [CrossRef] [Green Version]
- Buldain, D.; Gortari Castillo, L.; Buchamer, A.V.; Aliverti, F.; Bandoni, A.; Marchetti, L.; Mestorino, N. Melaleuca Armillaris Essential Oil in Combination With Rifaximin Against Staphylococcus aureus Isolated of Dairy Cows. Front. Vet. Sci. 2020, 7, 344. [Google Scholar] [CrossRef] [PubMed]
- Cerioli, M.F.; Moliva, M.V.; Cariddi, L.N.; Reinoso, E.B. Effect of the Essential Oil of Minthostachys Verticillata (Griseb.) Epling and Limonene on Biofilm Production in Pathogens Causing Bovine Mastitis. Front. Vet. Sci. 2018, 5, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montironi, I.D.; Cariddi, L.N.; Reinoso, E.B. Evaluation of the Antimicrobial Efficacy of Minthostachys Verticillata Essential Oil and Limonene against Streptococcus Uberis Strains Isolated from Bovine Mastitis. Rev. Argent. Microbiol. 2016, 48, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-Y.; Damte, D.; Lee, S.-J.; Kim, J.-C.; Park, S.-C. Antimicrobial Activity of Lemongrass and Oregano Essential Oil against Standard Antibiotic Resistant Staphylococcus aureus and Field Isolates from Chronic Mastitis Cow. Int. J. Phytomed. 2012, 4, 134. [Google Scholar]
- Ksouri, S.; Djebir, S.; Bentorki, A.A.; Gouri, A.; Hadef, Y.; Benakhla, A. Antifungal Activity of Essential Oils Extract from Origanum floribundum Munby, Rosmarinus officinalis L. and Thymus ciliatus Desf. against Candida albicans Isolated from Bovine Clinical Mastitis. J. Mycol. Médicale 2017, 27, 245–249. [Google Scholar] [CrossRef]
- Grzesiak, B.; Kołodziej, B.; Głowacka, A.; Krukowski, H. The Effect of Some Natural Essential Oils Against Bovine Mastitis Caused by Prototheca Zopfii Isolates In Vitro. Mycopathologia 2018, 183, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Kovačević, Z.; Kladar, N.; Čabarkapa, I.; Radinović, M.; Maletić, M.; Erdeljan, M.; Božin, B. New Perspective of Origanum vulgare L. and Satureja montana L. Essential Oils as Bovine Mastitis Treatment Alternatives. Antibiotics 2021, 10, 1460. [Google Scholar] [CrossRef] [PubMed]
- Tomanić, D.; Božin, B.; Kladar, N.; Stanojević, J.; Čabarkapa, I.; Stilinović, N.; Apić, J.; Božić, D.D.; Kovačević, Z. Environmental Bovine Mastitis Pathogens: Prevalence, Antimicrobial Susceptibility, and Sensitivity to Thymus vulgaris L., Thymus serpyllum L., and Origanum vulgare L. Essential Oils. Antibiotics 2022, 11, 1077. [Google Scholar] [CrossRef]
- Nardoni, S.; Pisseri, F.; Pistelli, L.; Najar, B.; Luini, M.; Mancianti, F. In Vitro Activity of 30 Essential Oils against Bovine Clinical Isolates of Prototheca Zopfii and Prototheca Blaschkeae. Vet. Sci. 2018, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Corona-Gómez, L.; Hernández-Andrade, L.; Mendoza-Elvira, S.; Ricardo-González, D.I.; Pérez-Gerardo, M.D.; Quintanar-Guerrero, D. Effect of the in Vitro Activity of a Component Derived from Thymus vulgaris Essential Oil and Tilmicosin on Different Strains of Staphylococcus sp, Streptococcus sp and Escherichia coli Isolated from Bovine Mastitis. J. Essent. Oil Res. 2022, 34, 270–277. [Google Scholar] [CrossRef]
- Elhouda, B.A.N.; Kheira, G.; Samia, M. Escherichia coli and Staphylococcus aureus Responsible for Bovine Mastitis Sensitivity to the Essential Oil of Algerian Thymus fontanesii Boiss. et Reut. Acta Sci. Nat. 2020, 7, 26–32. [Google Scholar] [CrossRef]
- Kovačević, Z.; Radinović, M.; Čabarkapa, I.; Kladar, N.; Božin, B. Natural Agents against Bovine Mastitis Pathogens. Antibiotics 2021, 10, 205. [Google Scholar] [CrossRef]
- Mullen, K.A.E.; Lee, A.R.; Lyman, R.L.; Mason, S.E.; Washburn, S.P.; Anderson, K.L. Short Communication: An in Vitro Assessment of the Antibacterial Activity of Plant-Derived Oils. J. Dairy Sci. 2014, 97, 5587–5591. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, N.G.; Croda, J.; Simionatto, S. Antibacterial Mechanisms of Cinnamon and Its Constituents: A Review. Microb. Pathog. 2018, 120, 198–203. [Google Scholar] [CrossRef]
- Rao, P.V.; Gan, S.H. Cinnamon: A Multifaceted Medicinal Plant. Evid. Based Complement. Alternat. Med. 2014, 2014, e642942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neculai-Valeanu, A.S.; Ariton, A.M.; Mădescu, B.M.; Rîmbu, C.M.; Creangă, Ş. Nanomaterials and Essential Oils as Candidates for Developing Novel Treatment Options for Bovine Mastitis. Animals 2021, 11, 1625. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Berkay Yılmaz, Y.; Antika, G.; Salehi, B.; Tumer, T.B.; Kulandaisamy Venil, C.; Das, G.; Patra, J.K.; Karazhan, N.; Akram, M.; et al. Phytochemical Constituents, Biological Activities, and Health-Promoting Effects of the Genus Origanum. Phytother. Res. PTR 2021, 35, 95–121. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Al-Khalifa, A.R.; Aouf, A.; Boukhebti, H.; Farouk, A. Effect of Nanoencapsulation on Volatile Constituents, and Antioxidant and Anticancer Activities of Algerian Origanum Glandulosum Desf. Essential Oil. Sci. Rep. 2020, 10, 2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- La Storia, A.; Ercolini, D.; Marinello, F.; Di Pasqua, R.; Villani, F.; Mauriello, G. Atomic Force Microscopy Analysis Shows Surface Structure Changes in Carvacrol-Treated Bacterial Cells. Res. Microbiol. 2011, 162, 164–172. [Google Scholar] [CrossRef]
- Di Pasqua, R.; Betts, G.; Hoskins, N.; Edwards, M.; Ercolini, D.; Mauriello, G. Membrane Toxicity of Antimicrobial Compounds from Essential Oils. J. Agric. Food Chem. 2007, 55, 4863–4870. [Google Scholar] [CrossRef] [PubMed]
- Salvo, A.; La Torre, G.L.; Rotondo, A.; Cicero, N.; Gargano, R.; Mangano, V.; Casale, K.E.; Dugo, G. Multiple Analytical Approaches for the Organic and Inorganic Characterization of Origanum vulgare L. Samples. Nat. Prod. Res. 2019, 33, 2815–2822. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, T.; Wang, X.; Shen, M.; Yan, X.; Fan, S.; Wang, L.; Wang, X.; Xu, X.; Sui, H.; et al. Traditional Uses, Chemical Constituents and Biological Activities of Plants from the Genus Thymus. Chem. Biodivers. 2019, 16, e1900254. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Salehi, B.; Varoni, E.M.; Sharopov, F.; Yousaf, Z.; Ayatollahi, S.A.; Kobarfard, F.; Sharifi-Rad, M.; Afdjei, M.H.; Sharifi-Rad, M.; et al. Plants of the Melaleuca Genus as Antimicrobial Agents: From Farm to Pharmacy. Phytother. Res. 2017, 31, 1475–1494. [Google Scholar] [CrossRef]
- Taga, I.; Lan, C.Q.; Altosaar, I. Plant Essential Oils and Mastitis Disease: Their Potential Inhibitory Effects on Pro-Inflammatory Cytokine Production in Response to Bacteria Related Inflammation. Nat. Prod. Commun. 2012, 7, 675–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, K.; Yang, T.; Feng, B.; Zhu, X.; Chen, Y.; Huo, Y.; Zhao, G. The Protective Roles of Tea Tree Oil Extracts in Bovine Mammary Epithelial Cells and Polymorphonuclear Leukocytes. J. Anim. Sci. Biotechnol. 2020, 11, 62. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, Y.; Zhou, J.; Lu, L.; Wang, X.; Liang, Y.; Loor, J.J.; Gou, D.; Xu, H.; Yang, Z. Tea Tree Oil Prevents Mastitis-Associated Inflammation in Lipopolysaccharide-Stimulated Bovine Mammary Epithelial Cells. Front. Vet. Sci. 2020, 7, 496. [Google Scholar] [CrossRef]
- Dobros, N.; Zawada, K.D.; Paradowska, K. Phytochemical Profiling, Antioxidant and Anti-Inflammatory Activity of Plants Belonging to the Lavandula Genus. Molecules 2023, 28, 256. [Google Scholar] [CrossRef]
- Turgut, A.C.; Emen, F.M.; Canbay, H.S.; Demirdöğen, R.E.; Çam, N.; Kılıç, D.; Yeşilkaynak, T. Chemical characterization of Lavandula angustifolia Mill. which is a phytocosmetic species and investigation of its antimicrobial effect in cosmetic products. J. Turk. Chem. Soc. Sect. Chem. 2017, 4, 283–298. [Google Scholar] [CrossRef] [Green Version]
- Danh, L.T.; Triet, N.D.A.; Han, L.T.N.; Zhao, J.; Mammucari, R.; Foster, N. Antioxidant Activity, Yield and Chemical Composition of Lavender Essential Oil Extracted by Supercritical CO2. J. Supercrit. Fluids 2012, 70, 27–34. [Google Scholar] [CrossRef]
- Montironi, I.D.; Reinoso, E.B.; Paullier, V.C.; Siri, M.I.; Pianzzola, M.J.; Moliva, M.; Campra, N.; Bagnis, G.; Ferreira LaRocque-de-Freitas, I.; Decote-Ricardo, D.; et al. Minthostachys Verticillata Essential Oil Activates Macrophage Phagocytosis and Modulates the Innate Immune Response in a Murine Model of Enterococcus Faecium Mastitis. Res. Vet. Sci. 2019, 125, 333–344. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Setzer, W.N. Biological Activities and Safety of Citrus Spp. Essential Oils. Int. J. Mol. Sci. 2018, 19, 1966. [Google Scholar] [CrossRef] [Green Version]
- Tisserand, R.; Young, R. Essential Oil Safety: A Guide for Health Care Professionals; Elsevier Health Sciences: Amsterdam, The Netherlands, 2013; ISBN 978-0-7020-5434-1. [Google Scholar]
- Tibenda, J.J.; Yi, Q.; Wang, X.; Zhao, Q. Review of Phytomedicine, Phytochemistry, Ethnopharmacology, Toxicology, and Pharmacological Activities of Cymbopogon Genus. Front. Pharmacol. 2022, 13, 997918. [Google Scholar] [CrossRef] [PubMed]
- Jisha, S.; Suja, R.S.; Bibu, J.K.; Surya, S.; Ambily, R.; Reni, J.; Thresia; Vidya, V.K.; Gayathri, P.; Usha, P.T.A. In Vitro Antimicrobial Activity of Lemongrass (Cymbopogon flexuosus) Oil and Citral against Methicillin Resistant Staphylococcus aureus (MRSA) from Bovine Mastitis. J. Vet. Anim. Sci. 2020, 51, 184–188. [Google Scholar]
- Ragab, A.; Dapgh, A.; ELsayed, H. The Antibacterial Activity of Some Essential Oils Nano Particles on Escherichia coli Infection in Dairy Farms. Alex. J. Vet. Sci. 2022, 72, 22. [Google Scholar] [CrossRef]
- Şarer, E.; Toprak, S.Y.; Otlu, B.; Durmaz, R. Composition and Antimicrobial Activity of the Essential Oil from Mentha spicata L. subsp. Spicata. J. Essent. Oil Res. 2011, 23, 105–108. [Google Scholar] [CrossRef]
- Mahendran, G.; Rahman, L.-U. Ethnomedicinal, Phytochemical and Pharmacological Updates on Peppermint (Mentha × piperita L.)—A Review. Phytother. Res. 2020, 34, 2088–2139. [Google Scholar] [CrossRef]
- Borges, R.S.; Ortiz, B.L.S.; Pereira, A.C.M.; Keita, H.; Carvalho, J.C.T. Rosmarinus Officinalis Essential Oil: A Review of Its Phytochemistry, Anti-Inflammatory Activity, and Mechanisms of Action Involved. J. Ethnopharmacol. 2019, 229, 29–45. [Google Scholar] [CrossRef]
- Ghorbani, A.; Esmaeilizadeh, M. Pharmacological Properties of Salvia Officinalis and Its Components. J. Tradit. Complement. Med. 2017, 7, 433–440. [Google Scholar] [CrossRef]
- Zarooni, S.; Rahchamani, R.; Ghanbari, F.; Khanahmadi, A. Antibacterial Effect of Satureja Hortensis and Salvia Officinalis Essential Oils against Major Bovine Mastitis Bacteria. Iran. J. Vet. Sci. Technol. 2021, 13, 75–81. [Google Scholar] [CrossRef]
- Haro-González, J.N.; Castillo-Herrera, G.A.; Martínez-Velázquez, M.; Espinosa-Andrews, H. Clove Essential Oil (Syzygium aromaticum L. Myrtaceae): Extraction, Chemical Composition, Food Applications, and Essential Bioactivity for Human Health. Molecules 2021, 26, 6387. [Google Scholar] [CrossRef] [PubMed]
- de Faria, M.J.M.; Braga, C.A.d.S.B.; de Paula, J.R.; André, M.C.D.P.B.; Vaz, B.G.; de Carvalho, T.C.; Romão, W.; da Costa, H.B.; da Conceição, E.C. Antimicrobial Activity of Copaifera Spp. against Bacteria Isolated from Milk of Cows with Mastitis. Ciênc. Anim. Bras. 2017, 18. [Google Scholar] [CrossRef] [Green Version]
- Tomanić, D.Z.; Stanojević, J.B.; Galić, I.M.; Ružić, Z.N.; Kukurić, T.B.; Tešin, N.B.; Prpa, B.P.; Kovačević, Z.R. Review of Trends in Essential Oils as Alternatives to Antibiotics in Bovine Mastitis Treatment. Zb. Matice Srp. Za Prir. Nauke 2022, 142, 47–60. [Google Scholar] [CrossRef]
- Campra, N.A.; Reinoso, E.B.; Montironi, I.D.; Moliva, M.V.; Raviolo, J.; Ruiz Moreno, F.; Marin, C.; Camacho, N.M.; Paredes, A.J.; Morán, M.C.; et al. Spray-Drying-Microencapsulated Minthostachys Verticillata Essential Oil and Limonene as Innovative Adjuvant Strategy to Bovine Mastitis Vaccines. Res. Vet. Sci. 2022, 149, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jiang, Y.; Yang, Y.; Wang, H.; Ye, J.; Liu, D.; Chen, Y.; Lian, C.; Wang, R.; Gao, Y.; et al. Houttuynia Essential Oil and Its Self-Microemulsion Preparation Protect Against LPS-Induced Murine Mastitis by Restoring the Blood–Milk Barrier and Inhibiting Inflammation. Front. Immunol. 2022, 13, 842189. [Google Scholar] [CrossRef]
- Hamlaoui, M.W.; Hadef, A.; Meklati, F.R.; Alleg, M.; Ghouila, B. In Vivo Effect of Origanum Vulgare Essential Oil on Microbiological Quality of Raw Milk from Subclinical Mastitis Affected Cows. Agric. For. 2021, 67, 83–98. [Google Scholar]
- Cho, B.-W.; Cha, C.-N.; Lee, S.-M.; Kim, M.-J.; Park, J.-Y.; Yoo, C.-Y.; Son, S.-E.; Kim, S.; Lee, H.-J. Therapeutic Effect of Oregano Essential Oil on Subclinical Bovine Mastitis Caused by Staphylococcus aureus and Escherichia coli. Korean J. Vet. Res. 2015, 55, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Pașca, C.; Mărghitaș, L.A.; Dezmirean, D.S.; Matei, I.A.; Bonta, V.; Pașca, I.; Chirilă, F.; Cîmpean, A.; Fiț, N.I. Efficacy of Natural Formulations in Bovine Mastitis Pathology: Alternative Solution to Antibiotic Treatment. J. Vet. Res. 2020, 64, 523–529. [Google Scholar] [CrossRef]
- Rasool, S.; Farooq, M.U.; Ahmad, S.S.; Avais, M.; Abbas, S.; Ambreen, N. Effect of Essential Oils of Eucalyptus Globulus and Lavandula Hybrida as Teat Dips to Control Subclinical Mastitis in Friesian Dairy Cattle. Eur. J. Med. Vet. Sci.-Nov. 2021, 2, 0100014EJMVS. [Google Scholar]
- Kovačević, Z.; Tomanić, D.; Čabarkapa, I.; Šarić, L.; Stanojević, J.; Bijelić, K.; Galić, I.; Ružić, Z.; Erdeljan, M.; Kladar, N. Chemical Composition, Antimicrobial Activity, and Withdrawal Period of Essential Oil-Based Pharmaceutical Formulation in Bovine Mastitis Treatment. Int. J. Environ. Res. Public. Health 2022, 19, 16643. [Google Scholar] [CrossRef]
- Tomanić, D.; Kladar, N.; Radinović, M.; Stančić, I.; Erdeljan, M.; Stanojević, J.; Galić, I.; Bijelić, K.; Kovačević, Z. Intramammary Ethno-Veterinary Formulation in Bovine Mastitis Treatment for Optimization of Antibiotic Use. Pathogens 2023, 12, 259. [Google Scholar] [CrossRef] [PubMed]
- Nehme, R.; Andrés, S.; Pereira, R.B.; Ben Jemaa, M.; Bouhallab, S.; Ceciliani, F.; López, S.; Rahali, F.Z.; Ksouri, R.; Pereira, D.M.; et al. Essential Oils in Livestock: From Health to Food Quality. Antioxidants 2021, 10, 330. [Google Scholar] [CrossRef] [PubMed]
- Tomanić, D.; Božin, B.; Čabarkapa, I.; Kladar, N.; Radinović, M.; Maletić, M.; Kovačević, Z. Chemical Composition, Antioxidant and Antibacterial Activity of Two Different Essential Oils Against Mastitis Associated Pathogens. Acta Vet. 2022, 72, 45–58. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Essential Oil | Main Component(s) | Tested Microorganisms | MIC | MBC | Zone Inhibition (mm) | Biofilm (%) | Reference |
|---|---|---|---|---|---|---|---|
| - | Carvacrol | S. aureus | 0.38 ± 0.00 mg/mL * | 0.75 ± 0.00 mg/mL * | 32.7 ± 3.01 | [74] | |
| - | Carvacrol | E. coli | 0.75 ± 0.00 mg/mL * | 1.25 ± 0.43 mg/mL * | 31.7 ± 0.58 | [74] | |
| - | Carvacrol | Klebsiella pneumoniae | 0.75 ± 0.00 mg/mL * | 1.5 ± 0.00 mg/mL * | 13.7 ± 0.58 | [74] | |
| - | Carvacrol | C. albicans | 0.1–0.2 mg/mL | 0.4–0.8 mg/mL | [75] | ||
| - | Carvacrol | Staphylococcus spp., Streptococcus spp. | 0.02–0.4 mg/mL | 0.2–0.78 mg/mL | [75] | ||
| - | Carvacrol | E. coli, Klebsiella pneumoniae | 0.02–0.2 mg/mL | 0.1–0.7 mg/mL | [75] | ||
| - | Carvacrol | Staphylococcus spp. | 584 µg/mL | 732 µg/mL | [76] | ||
| - | Cinnamaldehyde | S. aureus | 0.199 mg/mL | 69.4/44.9 | [77] | ||
| - | trans-Cinnamaldehyde | S. aureus | 0.3 ± 0.00 mg/mL * | 0.66 ± 0.00 mg/mL * | 25.3 ± 1.53 | [74] | |
| - | trans-Cinnamaldehyde | E. coli | 0.6 ± 0.00 mg/mL * | 1.32 ± 0.00 mg/mL * | 20 ± 1.00 | [74] | |
| - | trans-Cinnamaldehyde | Klebsiella pneumoniae | 0.6 ± 0.00 mg/mL * | 1.32 ± 0.00 mg/mL * | 11.0 ± 1.00 | [74] | |
| - | trans-Cinnamaldehyde | Staphylococcus spp. | 598 mg/L | 2238 mg/L | [78] | ||
| Cinnamon | / | S. agalactiae | 0.01 ± 0.01 μg/mL ** | 20.57–38.29 | 50% | [79] | |
| Cinnamomum aromaticum | e-Cinnamaldehyde (94.67%) | Staphylococcus spp. | 0.625 μL/mL | 1.25–10 μL/mL | [80] | ||
| Cinnamomum aromaticum | e-Cinnamaldehyde (94.67%) | Candida spp., Cryptococcus spp., Rhodotorula glutinis | 0.625–1.25 μL/mL | 1.25–2.5 μL/mL | [80] | ||
| Cinnamomum cassia | Cinnamaldehyde (59.96%) | S. aureus, S. epidermidis, S. xylosus | 0.0125% v/v | 0.05% v/v | 29.6, 19.1, 33.3 | [81] | |
| Cinnamomum cassia | Cinnamaldehyde (59.96%) | S. hyicus | 0.00625% v/v | 0.025% v/v | 27.0 | [81] | |
| Cinnamomum cassia | Cinnamaldehyde (59.96%) | E. coli | 0.025% v/v | 0.10% v/v | 30.7 | [81] | |
| Cinnamomum cassia | / | S. aureus, S. epidermidis, S. xylosus | 0.078–1.250% v/v | 0.039–1.250% v/v | [82] | ||
| Cinnamomum cassia | Cinnamaldehyde (98.29%) | C. albicans | 1 wt% | 5.0–25.0 | [83] | ||
| Cinnamomum cassia | Cinnamaldehyde (98.29%) | E. coli | 0.5–1 wt% | 3.0–21.0 | [83] | ||
| Cinnamomum cassia | Cinnamaldehyde (98.29%) | S. aureus | 0.5 wt% | 7.0–32.0 | [83] | ||
| Cinnamomum zeylanicum | Cinnamaldehyde (86.59%) | S. aureus | 0.243 mg/mL | 74.7/45.3 | [77] | ||
| Cinnamomum zeylanicum Ness | Cinnamaldehyde (76.8%), Methoxycinnamaldehyde (11.7%) | Prototheca zopfii | 0.2–0.4 μL/mL | [84] | |||
| Cinnamomum zeylanicum | Isoeugenol (96%) | Staphylococcus spp. | 2032 mg/L | 4263 mg/L | [78] | ||
| Lavandula angustifolia | / | Staphylococcus spp., Kytococcus sedentarius, Bacillus cereus | 15.17 ± 5.06 | [85] | |||
| Lavandula stoechas | 17-Pentatriacontene (42.15%), Linalyl acetate (26.82%), Eucalyptol (18.87%) | E. coli | 3.12% v/v | 6.25% v/v | [86] | ||
| Lavandula stoechas | 17-Pentatriacontene (42.15%), Linalyl acetate (26.82%), Eucalyptol (18.87%) | S. aureus | 4.37% v/v | 8.75% v/v | [86] | ||
| Lavandula stoechas | 17-Pentatriacontene (42.15%), Linalyl acetate (26.82%), Eucalyptol (18.87%) | S. agalactiae | 3.75% v/v | 7.50% v/v | [86] | ||
| Melaleuca alternifolia | Terpinen-4-ol (39.1%), γ-Terpinene (21.1%), α-Terpinene (9.2%) | C. albicans | 6.25 mg/mL | 6.25–12.50 mg/mL | [75] | ||
| Melaleuca alternifolia | Terpinen-4-ol (39.1%), γ-Terpinene (21.1%), α-Terpinene (9.2%) | Staphylococcus spp., Streptococcus spp. | 3.13–25 mg/mL | 6.25–52 mg/mL | [75] | ||
| Melaleuca alternifolia | Terpinen-4-ol (39.1%), γ-Terpinene (21.1%), α-Terpinene (9.2%) | E. coli, Klebsiella pneumoniae | 0.78–3.13 mg/mL | 6.25–12.50 mg/mL | [75] | ||
| Melaleuca alternifolia | / | Prototheca zopfii, P. wickerhamii | 0.03–0.12% | [87] | |||
| Melaleuca alternifolia | / | Candida parapsilosis, Candida krusei | 0.06–0.25% | [87] | |||
| Melaleuca armillaris | 1,8-Cineole (72.3%), Limonene (7.8%) | S. aureus | 6.25–25 µL/mL | 12.5–50 µL/mL | [88] | ||
| Melaleuca armillaris | 1,8-Cineole (72.3%), Limonene (7.8%) | S. aureus | 3.1–25 µL/mL | 12.5–50 µL/mL | [88] | ||
| Minthostachys verticillata | Pulegone (74.96%), Menthone (20.38%) | E. coli | 0.90–14.51 mg/mL | / | 36.51–89.60 | [89] | |
| Minthostachys verticillata | Pulegone (74.96%), Menthone (20.38%) | Enterococcus faecium | 3.63 mg/mL | 29 mg/mL | 36.51–89.60 | [89] | |
| Minthostachys verticillata | Pulegone (74.96%), Menthone (20.38%) | Bacillus pumilus | 1.8–29 mg/mL | / | 36.51–89.60 | [89] | |
| Minthostachys verticillata | Pulegone (51.7%), Menthone (37.8%) | Streptococcus uberis | 14.3–114.5 mg/mL | 114.5–229 mg/mL | 23.50–88.25 | [90] | |
| Oregano | Carvacrol (68.78%), o-cymene (9.80%) | S. aureus (MRSA) | 100–4000 μg/mL | 2.5 ± 0.5–9.5 ± 1.5 | [91] | ||
| Origanum floribundum Munby. | Thymol (50.47%), p-Cymene (24.22%), γ-Terpinene (11.27%) | C. albicans | 17.18–23.14% µg/mL (MIC 80%) | [92] | |||
| Origanum majorana L. | trans-Sabinene hydrate (38.10%), Borneol (11.10%), Sabinene (7.60%) | Prototheca zopfii | 0.25–0.5 µL/mL | [93] | |||
| Origanum majorana | 3-Cyclohexene-1-ol,4-methyl-1-(1-methylethyl)-,(R)-(44.84%), α-Terpineol (6.83%), p-Cymene (6.75%) | E. coli | 1.56% v/v | 3.12% v/v | [86] | ||
| Origanum majorana | 3-Cyclohexene-1-ol,4-methyl-1-(1-methylethyl)-,(R)-(44.84%), α-Terpineol (6.83%), p-Cymene (6.75%) | S. aureus | 0.62% v/v | 1.25% v/v | [86] | ||
| Origanum majorana | 3-Cyclohexene-1-ol,4-methyl-1-(1-methylethyl)-,(R)-(44.84%), α-Terpineol (6.83%), p-Cymene (6.75%) | S. agalactiae | 1.87% v/v | 3.75% v/v | [86] | ||
| Origanum vulgare | Carvacrol (92%) | Staphylococcus spp. | 1600 µg/mL | 2288 µg/mL | [76] | ||
| Origanum vulgare L. | Carvacrol (80.35%), p-Cymene (4.82%), Thymol (4.21%) | Streptococcus spp., E. coli, Cronobacter sakazakii, Klebsiella oxytoca, Staphylococcus spp. | 0.78–6.25 mg/mL, | 1.56–12.5 mg/mL | [94] | ||
| Origanum vulgare L. | Carvacrol (77.29%), p-Cymene (8.85%), γ-Terpinene (4.96%) | Prototheca zopfii | 0.5–1 µL/mL | [93] | |||
| Origanum vulgare L. | Carvacrol (78.94%), Thymol (4.87%), p-Cymene (4.52%) | Proteus mirabilis | 3.125 ± 1.35 mg/mL * | 3.125 ± 1.35 mg/mL * | [95] | ||
| Origanum vulgare L. | Carvacrol (78.94%), Thymol (4.87%), p-Cymene (4.52%) | Serratia marcescens | 3.125 ± 1.91 mg/mL * | 6.25 ± 3.83 mg/mL * | [95] | ||
| Origanum vulgare | Carvacrol (90.50%) | C. albicans | 0.3 wt% | 3.0–25.0 | [83] | ||
| Origanum vulgare | Carvacrol (90.50%) | E. coli | 0.5 wt% | 3.0–23.5 | [83] | ||
| Origanum vulgare | Carvacrol (90.50%) | S. aureus | 0.3–0.5 wt% | 5.0–24.5 | [83] | ||
| Origanum vulgare | Carvacrol (65.9%), p-Cymene (9.3%) | P. zopfii, P. blaschkeae | 0.75/1% | [96] | |||
| - | Thymol | Staphylococcus spp. | 429.68 ± 123.53 µg/mL | 859.38 ± 247.05 µg/mL | [97] | ||
| - | Thymol | Streptococcus spp. | 664.06 ± 370.58 µg/mL | 1328.13 ± 741.16 µg/mL | [97] | ||
| - | Thymol | E. coli | 976.55 ± 887.91 µg/mL | 1953.1 ± 1775.82 µg/mL | [97] | ||
| - | Thymol | C. albicans | 0.05–0.4 mg/mL | 0.4–1.6 mg/mL | 43.8 ± 0.2 | [75] | |
| - | Thymol | Staphylococcus spp., Streptococcus spp. | 0.1–0.2 mg/mL | 0.2–0.4 mg/mL | 42.2 ± 5.5 | [75] | |
| - | Thymol | E. coli, Klebsiella pneumoniae | 0.1–0.2 mg/mL | 0.4 mg/mL | 31.2 ± 0.7 | [75] | |
| - | Thymol | S. aureus | 0.75 ± 0.00 mg/mL * | 1.5 ± 0.00 mg/mL * | 24.3 ± 2.00 | [74] | |
| - | Thymol | E. coli | 0.38 ± 0.00 mg/mL * | 0.63 ± 0.21 mg/mL * | 20.7 ± 1.16 | [74] | |
| - | Thymol | Klebsiella pneumoniae | 0.75 ± 0.00 mg/mL * | 1.5 ± 0.00 mg/mL * | 13.5 ± 0.50 | [74] | |
| - | Thymol | Staphylococcus spp. | 427 µg/mL | 856 µg/mL | [76] | ||
| Thymus ciliatus Desf. | Thymol (62.41%), p-Cymene (15.51%), Carvacrol (6.12%) | C. albicans | 15.02–20.96 µg/mL (MIC 80%) | [92] | |||
| Thymus fontanesii Boiss. Et Reut. | Carvacrol (62.25%) | E. coli | 1/1600 µL/mL | 0.625 µL/mL | 25.33 ± 1.53 | [98] | |
| Thymus fontanesii Boiss. Et Reut. | Carvacrol (62.25%) | S. aureus | 1/1600 µL/mL | 0.625 µL/mL | 35 ± 0.7 | [98] | |
| Thymus serpyllum | Thymol (54.17%), γ-Terpinene (22.18%), p-Cymene (16.66%) | Streptococcus spp. | 0.78–3.125 mg/mL | 1.56–6.25 mg/mL | [99] | ||
| Thymus serpyllum | Thymol (54.17%), γ-Terpinene (22.18%), p-Cymene (16.66%) | E. coli, Enterobacter sakazakii | 6.25 mg/mL | 12.5 mg/mL | [99] | ||
| Thymus serpyllum | Thymol (54.17%), γ-Terpinene (22.18%), p-Cymene (16.66%) | Klebsiella oxytoca, Staphylococcus spp. | 3.125 mg/mL | 6.25 mg/mL | [99] | ||
| Thymus serpyllum L. | Thymol (55.11%), γ-Terpinene (22.31%), p-Cymene (16.66%) | Proteus mirabilis | 3.125 ± 1.35 mg/mL * | 6.25 ± 2.7 mg/mL * | [95] | ||
| Thymus serpyllum L. | Thymol (55.11%), γ-Terpinene (22.31%), p-Cymene (16.66%) | Serratia marcescens | 1.56 ± 0.96 mg/mL * | 3.125 ± 1.91 mg/mL * | [95] | ||
| Thymus vulgaris | Thymol (45.22%), p-Cymene (23.83%) | Streptococcus spp. | 0.39–1.56 mg/mL | 0.78–6.25 mg/mL | [99] | ||
| Thymus vulgaris | Thymol (45.22%), p-Cymene (23.83%) | E. coli, Enterobacter sakazakii | 3.125 mg/mL | 6.25 mg/mL | [99] | ||
| Thymus vulgaris | Thymol (45.22%), p-Cymene (23.83%) | Klebsiella oxytoca, Staphylococcus spp. | 1.56 mg/mL/6.25 mg/mL | 6.25 mg/mL/12.5 mg/mL | [99] | ||
| Thymus vulgaris L. | Thymol (52.96%), p-Cymene (17.73%), γ-Terpinene (5.97%) | Prototheca zopfii | 0.25–1 µL/mL | [93] | |||
| Thymus vulgaris L. | Thymol (38.1%), p-Cymene (29.1%), γ-Terpinene (5.2%) | Prototheca zopfii | 0.6–1.0 μL/mL | [84] | |||
| Thymus vulgaris | Thymol (52.6%), p-Cymene (15.3%) | P. zopfii, P. blaschkeae | 0.75/1% | [96] | |||
| Thymus vulgaris | / | S. aureus, S. chromogenes, S. uberis | 2% v/v | [100] | |||
| Thymus vulgaris | / | S. aureus | 5% | 5% | 17.83–37.5 | [26] | |
| Thymus vulgaris | / | Streptococcus agalactiae | 4% | 5% | 13.5–30.83 | [26] | |
| Thymus vulgaris | / | E. coli | 1% | 1% | 14.66–34.33 | [26] | |
| Thymus vulgaris | γ-Terpinene (64%) | Staphylococcus spp. | 1564 µg/mL | 2370 µg/mL | [76] | ||
| Thymus vulgaris L. | Thymol (46.37%), p-Cymene (23.83%), γ-Terpinene (3.46%) | Proteus mirabilis | 3.125 ± 0.00 mg/mL * | 6.25 ± 2.7 mg/mL * | [95] | ||
| Thymus vulgaris L. | Thymol (46.37%), p-Cymene (23.83%), γ-Terpinene (3.46%) | Serratia marcescens | 1.56 ± 0.96 mg/mL * | 3.125 ± 1.91 mg/mL * | [95] | ||
| Thymus vulgaris | / | S. aureus, S. epidermidis, S. xylosus | 0.010–0.625 v/v% | 0.010–0.625 v/v% | [82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caneschi, A.; Bardhi, A.; Barbarossa, A.; Zaghini, A. Plant Essential Oils as a Tool in the Control of Bovine Mastitis: An Update. Molecules 2023, 28, 3425. https://doi.org/10.3390/molecules28083425
Caneschi A, Bardhi A, Barbarossa A, Zaghini A. Plant Essential Oils as a Tool in the Control of Bovine Mastitis: An Update. Molecules. 2023; 28(8):3425. https://doi.org/10.3390/molecules28083425
Chicago/Turabian StyleCaneschi, Alice, Anisa Bardhi, Andrea Barbarossa, and Anna Zaghini. 2023. "Plant Essential Oils as a Tool in the Control of Bovine Mastitis: An Update" Molecules 28, no. 8: 3425. https://doi.org/10.3390/molecules28083425
APA StyleCaneschi, A., Bardhi, A., Barbarossa, A., & Zaghini, A. (2023). Plant Essential Oils as a Tool in the Control of Bovine Mastitis: An Update. Molecules, 28(8), 3425. https://doi.org/10.3390/molecules28083425