

Dopamine- and Grape-Seed-Extract-Loaded Solid Lipid Nanoparticles: Interaction Studies between Particles and Differentiated SH-SY5Y Neuronal Cell Model of Parkinson’s Disease

 ,
,  ,
,  , , , , ,
, , , , ,  , ,
, , 
Abstract
:1. Introduction
2. Results
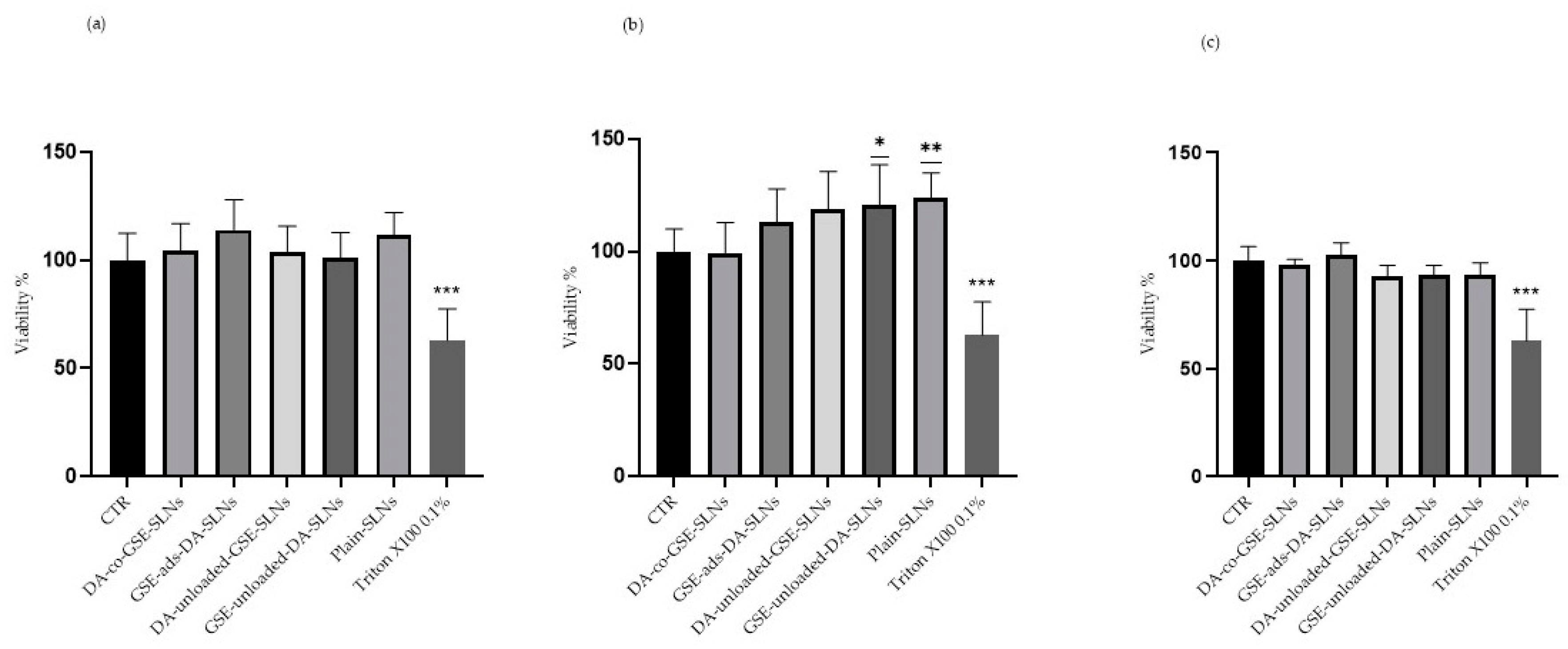
2.1. DA/GSE SLNs Show Cytobiocompatibility in SH-SY5Y-Differentiated Cell Model
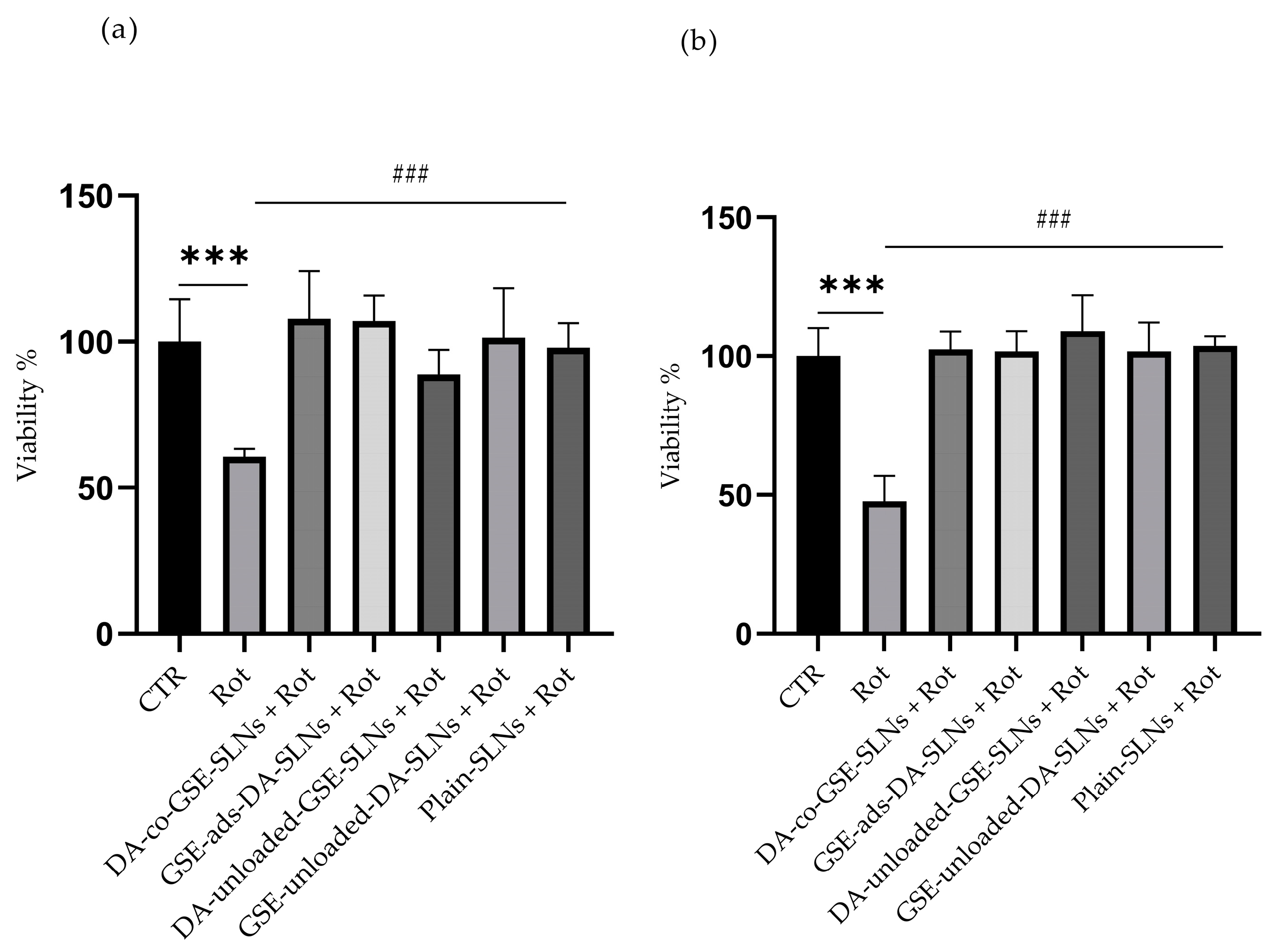
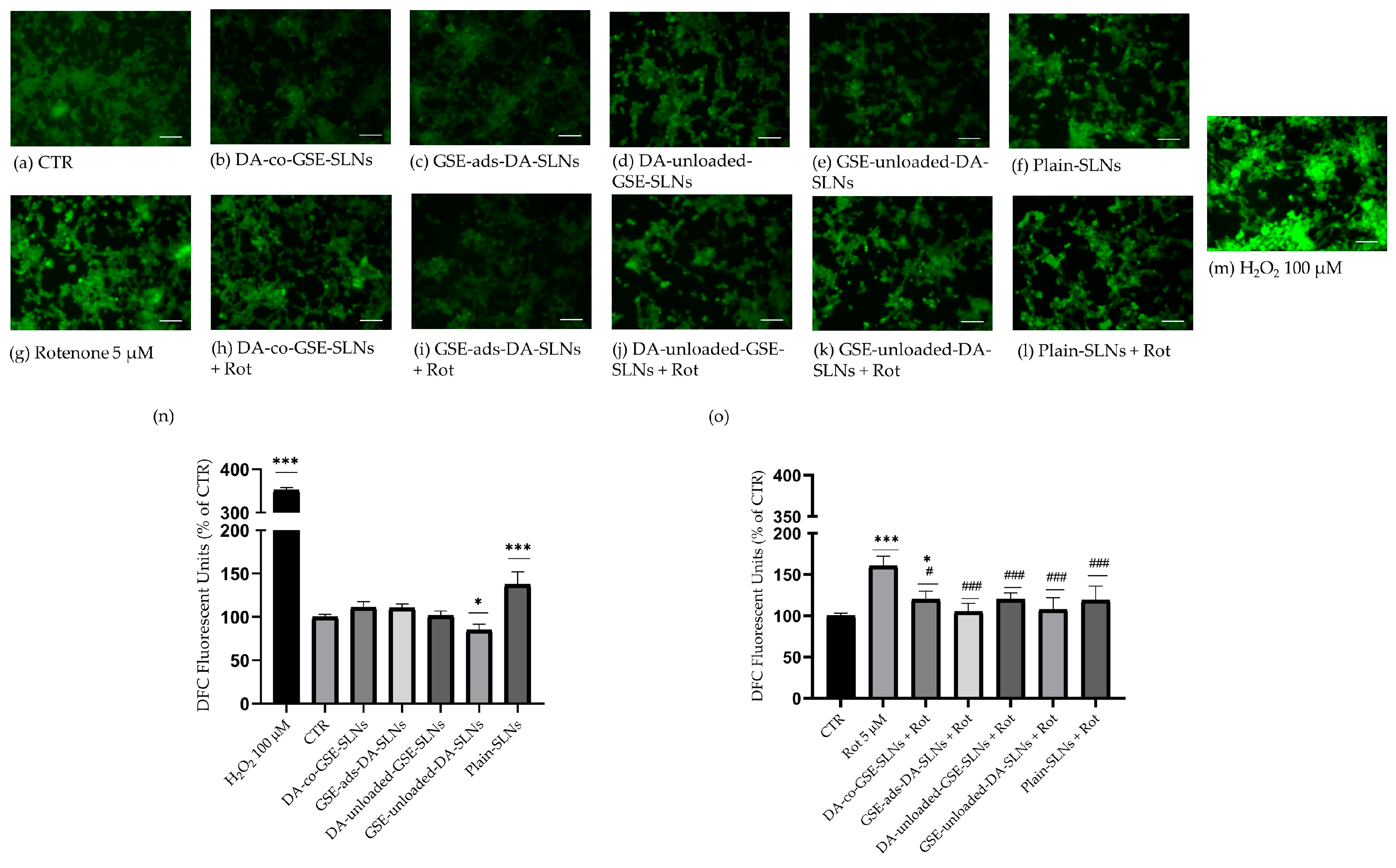
2.2. DA/GSE-SLNs Are Cytoprotective against Rotenone-Induced Toxicity and Oxidative Stress
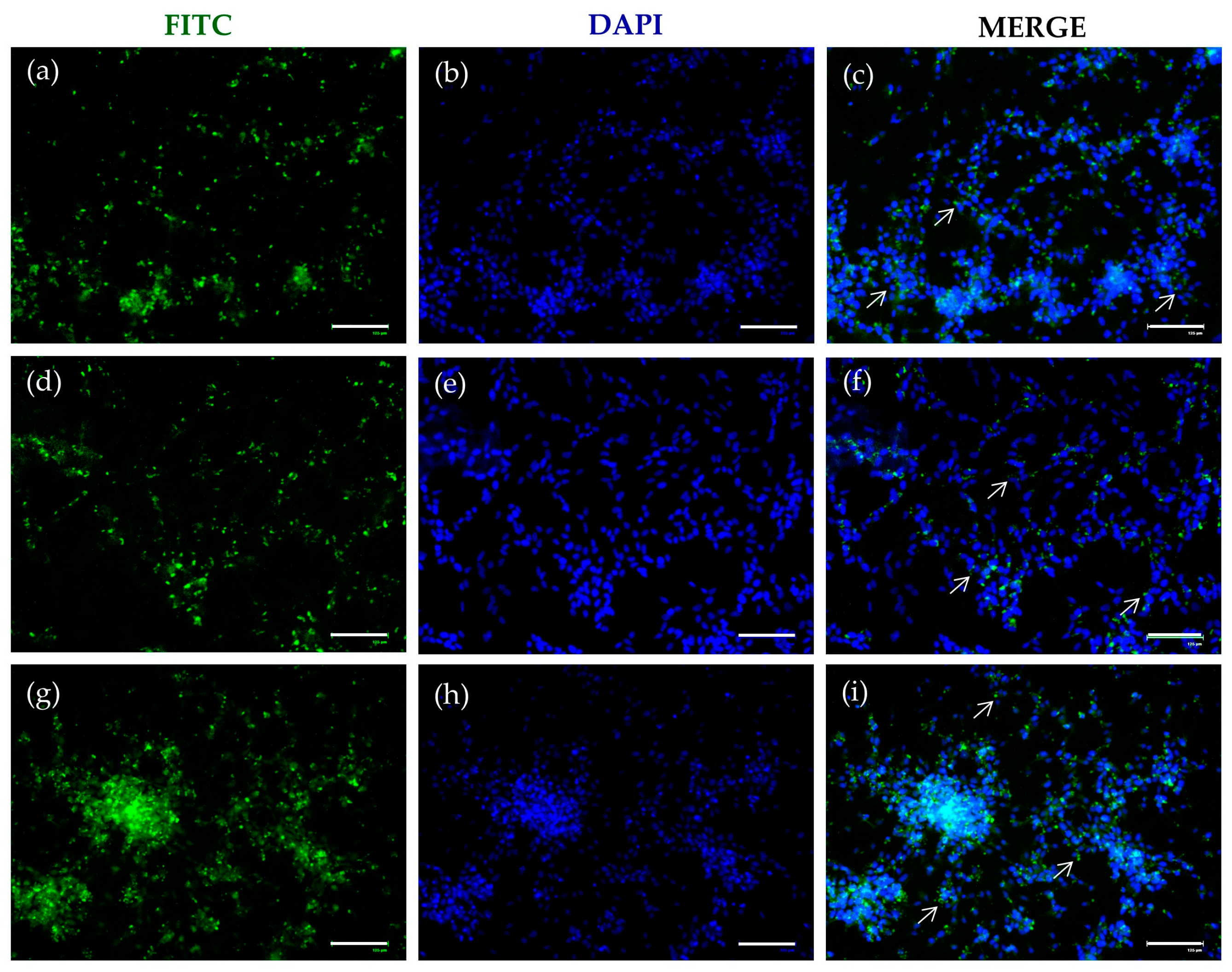
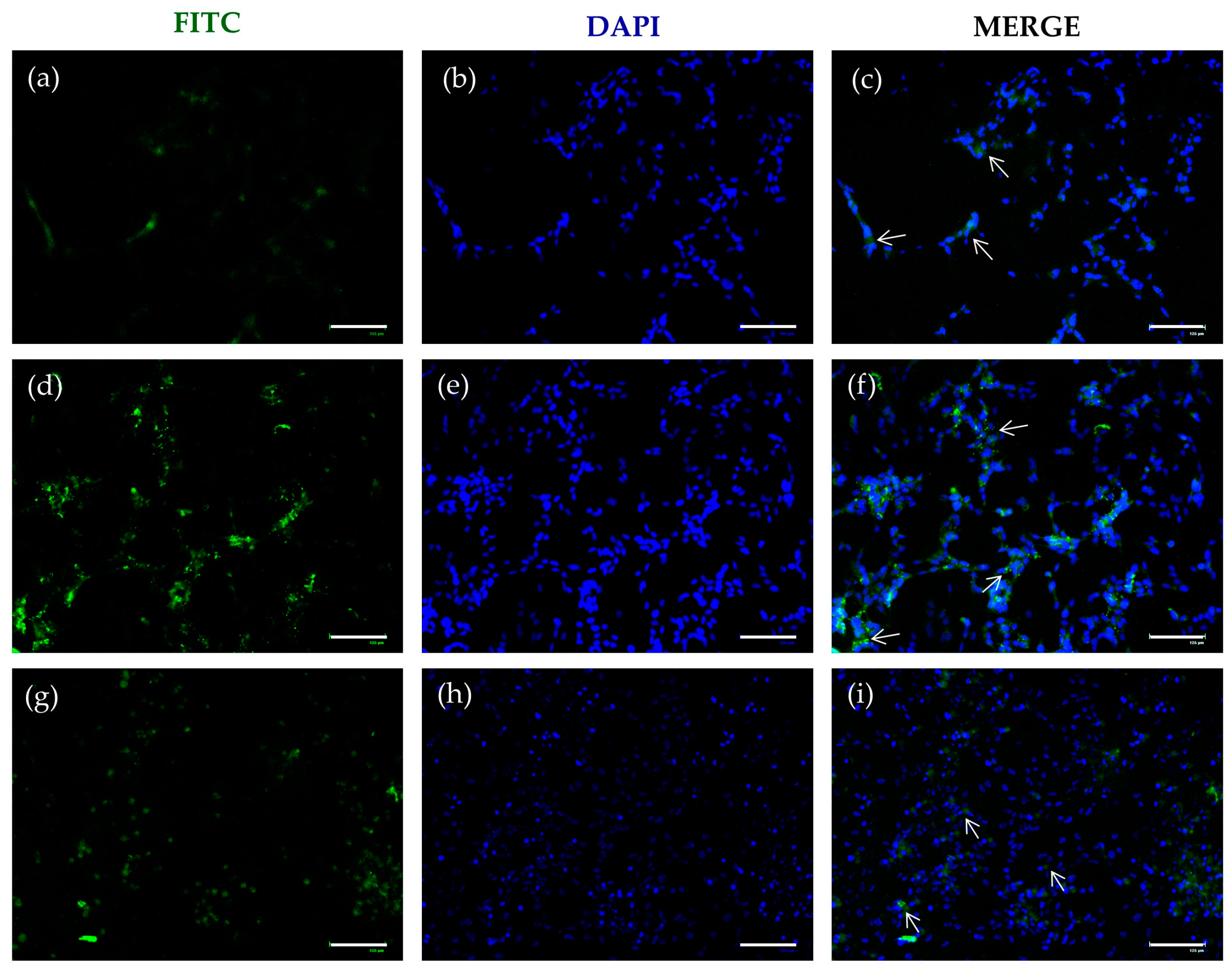
2.3. FITC-SLNs Are Detected by Fluorescence Microscopy
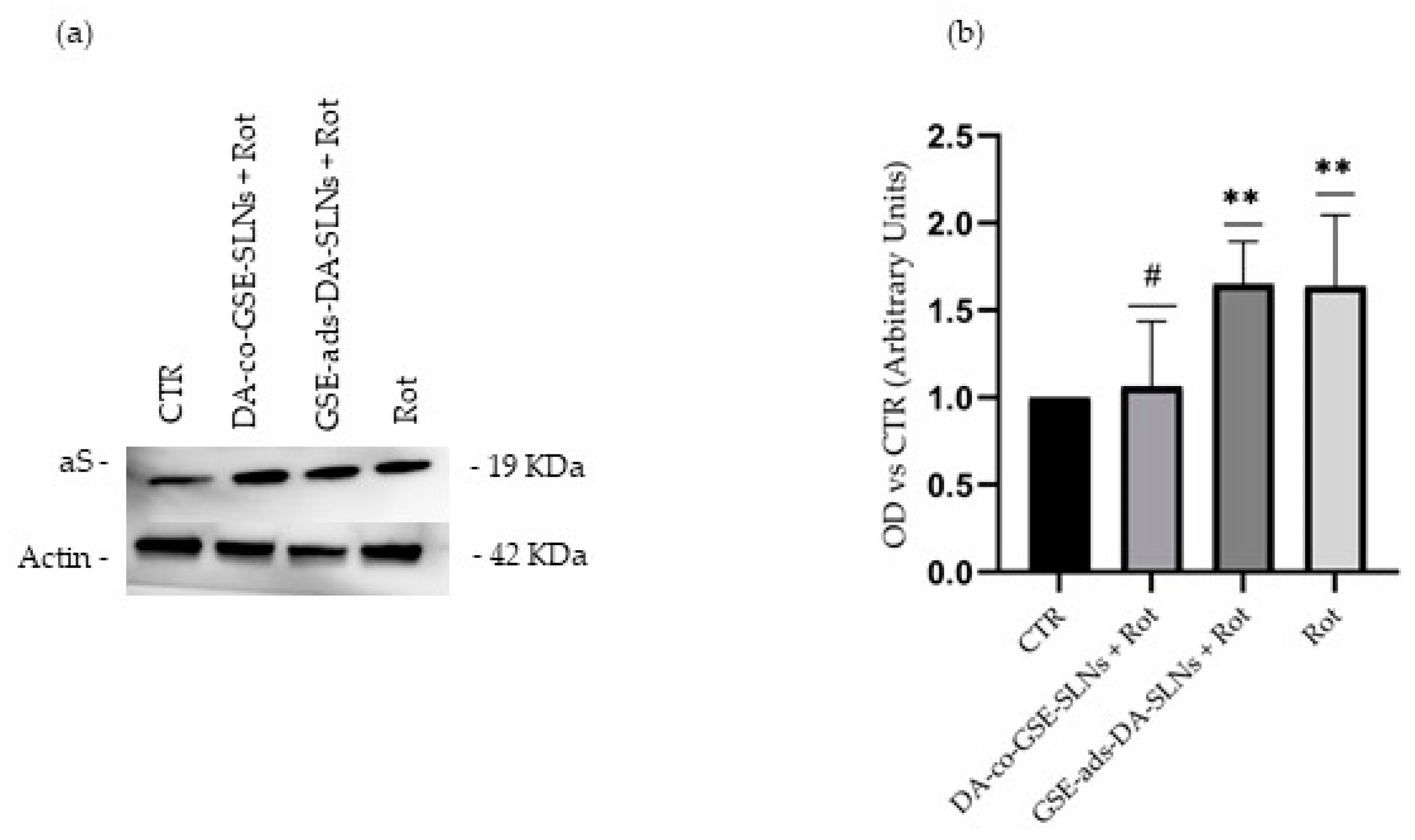
2.4. DA-co-GSE-SLNs Reduce Rotenone-Mediated Increase in Intracellular aS Levels
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. DA/GSE-SLN Preparation
- (i)
- DA-coencapsulating GSE SLNs (DA-co-GSE-SLNs). The formulation “DA-co-GSE-SLNs” was obtained following the melt homogenization method [49,65]. An amount of 60 mg of Gelucire® 50/13 was melted at 70 °C and, separately, 6 mg of GSE was dispersed in the aqueous phase made of surfactant (Tween® 85, 60 mg) and 1.37 mL diluted acetic acid (0.01%, w/v). The predispersion was homogenized at 12,300 rpm with an Ultra-Turrax model T25 apparatus (Janke and Kunkel, IKA®-Werke GmbH & Co., Staufen, Germany) and allowed to equilibrate for 30 min at 70 °C. Then, 10 mg of DA was introduced in the aqueous phase, the resulting mixture was added to the melted phase at 70 °C, and the so-obtained emulsion was homogenized at 12,300 rpm for 2 min by the Ultra-Turrax system. Then, the nanosuspension was cooled at room temperature and allowed to achieve SLN DA-coencapsulating GSE. Such SLNs were centrifuged (16,000× g, 45 min, Eppendorf 5415D, Hamburg, Germany) and the pellet was harvested and re-suspended in distilled water for further studies, but the supernatant was discarded.
- (ii)
- GSEadsorbing DA-SLNs (GSE-ads-DA-SLNs). For DA SLNs adsorbing GSE (called GSE-ads-DA-SLNs), DA-loaded SLNs were prepared as previously described [32], but starting from 20 mg of DA rather than 10 mg to force DA initial cargo. After cooling down at room temperature, an aliquot of 0.5 mL of the resulting DA-SLNs was incubated with 1 mL of GSE aqueous solution (1 mg/mL concentration) at room temperature for 3 h in the dark while maintaining mild stirring (50 oscillations/min). Then, the mixture was centrifuged at 16,000× g for 45 min (Eppendorf 5415D) and the pellet was dispersed in distilled water, whereas the supernatant was discarded. For comparison, the following formulations were also tested in the biological assays: SLNs loaded with DA only, formulated without GSE (called “GSE-unloaded-DA-SLNs”), and SLNs containing GSE without DA (called “DA-unloaded-GSE-SLNs”).
- (iii)
- As the control, SLNs with neither DA nor GSE were formulated following the melt homogenization method as reported above and are indicated as plain-SLNs in this paper. For the fluorescence microscopy application, two types of fluorescent SLN carriers were obtained as follows. Firstly, to achieve the fluorescent “DA-FITC-SLNs coencapsulating GSE”, 10 mg of DA-FITC replaced 10 mg of DA and the protocol was the same as that shown in Section 2.2. (i). Another fluorescent formulation of SLNs was produced by adsorbing 1 mL of GSE aqueous solution (1 mg/mL) onto 0.5 mL of preformed DA-FITC-SLNs. The physical adsorption was carried out for 3 h at room temperature and in the dark, leading to the formation of SLNs called “GSE-ads-DA-FITC SLNs”.
4.3. In Vitro Properties of DA/GSE SLNs
4.4. Cell Culture and Treatments
4.5. DA/GSE-SLN Cytobiocompatibility Evaluation
4.6. Evaluation of DA/GSE-SLN Cytoprotective Effects
4.7. FITC-DA/GSE-SLN Fluorescence Microscopy
4.8. DA/GSE-SLN Antioxidant Property Assay
4.9. Western Blotting
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dickson, D.W. Neuropathology of Parkinson Disease. Park. Relat. Disord. 2018, 46, S30–S33. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Benito, M.; Granado, N.; García-Sanz, P.; Michel, A.; Dumoulin, M.; Moratalla, R. Modeling Parkinson’s Disease with the Alpha-Synuclein Protein. Front. Pharmacol. 2020, 11, 356. [Google Scholar] [CrossRef] [PubMed]
- Speranza, L.; di Porzio, U.; Viggiano, D.; de Donato, A.; Volpicelli, F. Dopamine: The Neuromodulator of Long-Term Synaptic Plasticity, Reward and Movement Control. Cells 2021, 10, 735. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.Y.-Y.; Ho, P.W.-L.; Liu, H.-F.; Leung, C.-T.; Li, L.; Chang, E.E.S.; Ramsden, D.B.; Ho, S.-L. The Interplay of Aging, Genetics and Environmental Factors in the Pathogenesis of Parkinson’s Disease. Transl. Neurodegener. 2019, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Rojo, A.I.; Manda, G.; Boscá, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef]
- Mao, Q.; Qin, W.; Zhang, A.; Ye, N. Recent Advances in Dopaminergic Strategies for the Treatment of Parkinson’s Disease. Acta Pharmacol. Sin. 2020, 41, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Seven, E.S.; Seven, Y.B.; Zhou, Y.; Poudel-Sharma, S.; Diaz-Rucco, J.J.; Kirbas Cilingir, E.; Mitchell, G.S.; Van Dyken, J.D.; Leblanc, R.M. Crossing the Blood–Brain Barrier with Carbon Dots: Uptake Mechanism and in Vivo Cargo Delivery. Nanoscale Adv. 2021, 3, 3942–3953. [Google Scholar] [CrossRef]
- Yeragani, V.; Tancer, M.; Chokka, P.; Baker, G. Arvid Carlsson, and the Story of Dopamine. Indian J. Psychiatry 2010, 52, 87–88. [Google Scholar] [CrossRef]
- Caelsson, A.; Lindqvist, M.; Magnusson, T.; Waldeck, B. On the Presence of 3-Hydroxytyramine in Brain. Science 1958, 127, 471. [Google Scholar] [CrossRef]
- Haddad, F.; Sawalha, M.; Khawaja, Y.; Najjar, A.; Karaman, R. Dopamine and Levodopa Prodrugs for the Treatment of Parkinson’s Disease. Molecules 2017, 23, 40. [Google Scholar] [CrossRef]
- Leta, V.; Klingelhoefer, L.; Longardner, K.; Campagnolo, M.; Levent, H.Ç.; Aureli, F.; Metta, V.; Bhidayasiri, R.; Chung-Faye, G.; Falup-Pecurariu, C.; et al. Gastrointestinal Barriers to Levodopa Transport and Absorption in Parkinson’s Disease. Eur. J. Neurol. 2023, 30, 1465–1480. [Google Scholar] [CrossRef] [PubMed]
- Gupta, H.V.; Lyons, K.E.; Wachter, N.; Pahwa, R. Long Term Response to Levodopa in Parkinson’s Disease. J. Park. Dis. 2019, 9, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.-W.; Park, T.-E. Recent Advances with Liposomes as Drug Carriers for Treatment of Neurodegenerative Diseases. Biomed. Eng. Lett. 2021, 11, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. The Blood-Brain Barrier: Bottleneck in Brain Drug Development. NeuroRX 2005, 2, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Johnston, T.H.; Fox, S.H.; Brotchie, J.M. Advances in the Delivery of Treatments for Parkinson’s Disease. Expert Opin. Drug Deliv. 2005, 2, 1059–1073. [Google Scholar] [CrossRef] [PubMed]
- Priya, S.; Desai, V.M.; Singhvi, G. Surface Modification of Lipid-Based Nanocarriers: A Potential Approach to Enhance Targeted Drug Delivery. ACS Omega 2023, 8, 74–86. [Google Scholar] [CrossRef]
- Mishra, D.K.; Shandilya, R.; Mishra, P.K. Lipid Based Nanocarriers: A Translational Perspective. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 2023–2050. [Google Scholar] [CrossRef] [PubMed]
- Rezigue, M. Lipid and Polymeric Nanoparticles: Drug Delivery Applications. In Integrative Nanomedicine for New Therapies; Springer Nature: Berlin, Germany, 2020; pp. 167–230. [Google Scholar]
- Santonocito, D.; Sarpietro, M.G.; Carbone, C.; Panico, A.; Campisi, A.; Siciliano, E.A.; Sposito, G.; Castelli, F.; Puglia, C. Curcumin Containing PEGylated Solid Lipid Nanoparticles for Systemic Administration: A Preliminary Study. Molecules 2020, 25, 2991. [Google Scholar] [CrossRef]
- Sandhu, S.K.; Kumar, S.; Raut, J.; Singh, M.; Kaur, S.; Sharma, G.; Roldan, T.L.; Trehan, S.; Holloway, J.; Wahler, G.; et al. Systematic Development and Characterization of Novel, High Drug-Loaded, Photostable, Curcumin Solid Lipid Nanoparticle Hydrogel for Wound Healing. Antioxidants 2021, 10, 725. [Google Scholar] [CrossRef]
- Kakkar, V.; Singh, S.; Singla, D.; Kaur, I.P. Exploring Solid Lipid Nanoparticles to Enhance the Oral Bioavailability of Curcumin. Mol. Nutr. Food Res. 2011, 55, 495–503. [Google Scholar] [CrossRef]
- Reis, S.; Neves, A.R.; Lúcio, M.; Martins, S.; Lima, J.L. Novel Resveratrol Nanodelivery Systems Based on Lipid Nanoparticles to Enhance Its Oral Bioavailability. Int. J. Nanomed. 2013, 8, 177–187. [Google Scholar] [CrossRef]
- Trapani, A.; Mandracchia, D.; Tripodo, G.; Di Gioia, S.; Castellani, S.; Cioffi, N.; Ditaranto, N.; Esteban, M.A.; Conese, M. Solid Lipid Nanoparticles Made of Self-Emulsifying Lipids for Efficient Encapsulation of Hydrophilic Substances. AIP Conf. Proc. 2019, 2145, 020004. [Google Scholar]
- Surguchov, A.; Surguchev, A. Synucleins: New Data on Misfolding, Aggregation and Role in Diseases. Biomedicines 2022, 10, 3241. [Google Scholar] [CrossRef] [PubMed]
- Dorszewska, J.; Kowalska, M.; Prendecki, M.; Piekut, T.; Kozłowska, J.; Kozubski, W. Oxidative Stress Factors in Parkinson’s Disease. Neural Regen. Res. 2021, 16, 1383–1391. [Google Scholar] [CrossRef]
- Leitão, A.D.G.; Rudolffi-Soto, P.; Chappard, A.; Bhumkar, A.; Lau, D.; Hunter, D.J.B.; Gambin, Y.; Sierecki, E. Selectivity of Lewy Body Protein Interactions along the Aggregation Pathway of α-Synuclein. Commun. Biol. 2021, 4, 1124. [Google Scholar] [CrossRef]
- Goedert, M.; Spillantini, M.G.; Del Tredici, K.; Braak, H. 100 Years of Lewy Pathology. Nat. Rev. Neurol. 2013, 9, 13–24. [Google Scholar] [CrossRef]
- Duarte-Jurado, A.P.; Gopar-Cuevas, Y.; Saucedo-Cardenas, O.; Loera-Arias, M.D.J.; Montes-de-Oca-Luna, R.; Garcia-Garcia, A.; Rodriguez-Rocha, H. Antioxidant Therapeutics in Parkinson’s Disease: Current Challenges and Opportunities. Antioxidants 2021, 10, 453. [Google Scholar] [CrossRef]
- Murphy, M.P. Antioxidants as Therapies: Can We Improve on Nature? Free Radic. Biol. Med. 2014, 66, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, A.M.; Meléndez, C.M.; Uribe, D.; Pedroza-Díaz, J. Synergistic Effects of Natural Compounds and Conventional Chemotherapeutic Agents: Recent Insights for the Development of Cancer Treatment Strategies. Heliyon 2022, 8, e09519. [Google Scholar] [CrossRef]
- De Giglio, E.; Bakowsky, U.; Engelhardt, K.; Caponio, A.; La Pietra, M.; Cometa, S.; Castellani, S.; Guerra, L.; Fracchiolla, G.; Poeta, M.L.; et al. Solid Lipid Nanoparticles Containing Dopamine and Grape Seed Extract: Freeze-Drying with Cryoprotection as a Formulation Strategy to Achieve Nasal Powders. Molecules 2023, 28, 7706. [Google Scholar] [CrossRef]
- Trapani, A.; Castellani, S.; Guerra, L.; De Giglio, E.; Fracchiolla, G.; Corbo, F.; Cioffi, N.; Passantino, G.; Poeta, M.L.; Montemurro, P.; et al. Combined Dopamine and Grape Seed Extract-Loaded Solid Lipid Nanoparticles: Nasal Mucosa Permeation, and Uptake by Olfactory Ensheathing Cells and Neuronal SH-SY5Y Cells. Pharmaceutics 2023, 15, 881. [Google Scholar] [CrossRef] [PubMed]
- Aresta, A.; Calvano, C.D.; Trapani, A.; Cellamare, S.; Zambonin, C.G.; De Giglio, E. Development and Analytical Characterization of Vitamin(s)-Loaded Chitosan Nanoparticles for Potential Food Packaging Applications. J. Nanoparticle Res. 2013, 15, 1592. [Google Scholar] [CrossRef]
- Tripodo, G.; Perteghella, S.; Grisoli, P.; Trapani, A.; Torre, M.L.; Mandracchia, D. Drug Delivery of Rifampicin by Natural Micelles Based on Inulin: Physicochemical Properties, Antibacterial Activity and Human Macrophages Uptake. Eur. J. Pharm. Biopharm. 2019, 136, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Magalingam, K.B.; Radhakrishnan, A.K.; Somanath, S.D.; Md, S.; Haleagrahara, N. Influence of Serum Concentration in Retinoic Acid and Phorbol Ester Induced Differentiation of SH-SY5Y Human Neuroblastoma Cell Line. Mol. Biol. Rep. 2020, 47, 8775–8788. [Google Scholar] [CrossRef] [PubMed]
- Shipley, M.M.; Mangold, C.A.; Szpara, M.L. Differentiation of the SH-SY5Y Human Neuroblastoma Cell Line. J. Vis. Exp. 2016, e53193. [Google Scholar]
- Greco, M.; Munir, A.; Musarò, D.; Coppola, C.; Maffia, M. Restoring Autophagic Function: A Case for Type 2 Diabetes Mellitus Drug Repurposing in Parkinson’s Disease. Front. Neurosci. 2023, 17, 1244022. [Google Scholar] [CrossRef] [PubMed]
- Topal, G.R.; Mészáros, M.; Porkoláb, G.; Szecskó, A.; Polgár, T.F.; Siklós, L.; Deli, M.A.; Veszelka, S.; Bozkir, A. ApoE-Targeting Increases the Transfer of Solid Lipid Nanoparticles with Donepezil Cargo across a Culture Model of the Blood–Brain Barrier. Pharmaceutics 2020, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, L.; Dimmito, M.P.; Cacciatore, I.; Toto, E.C.; Di Rienzo, A.; Palmerio, F.; Puca, V.; Di Filippo, E.S.; Fulle, S.; Di Stefano, A. Solid Lipid Nanoparticles for Efficient Delivery of Capsaicin-Rich Extract: Potential Neuroprotective Effects in Parkinson’s Disease. J. Drug Deliv. Sci. Technol. 2024, 91, 105097. [Google Scholar] [CrossRef]
- Torres, J.; Pereira, J.M.; Marques-Oliveira, R.; Costa, I.; Gil-Martins, E.; Silva, R.; Remião, F.; Peixoto, A.F.; Sousa Lobo, J.M.; Silva, A.C. An in Vitro Evaluation of the Potential Neuroprotective Effects of Intranasal Lipid Nanoparticles Containing Astaxanthin Obtained from Different Sources: Comparative Studies. Pharmaceutics 2023, 15, 1035. [Google Scholar] [CrossRef]
- Amiri, M.; Jafari, S.; Kurd, M.; Mohamadpour, H.; Khayati, M.; Ghobadinezhad, F.; Tavallaei, O.; Derakhshankhah, H.; Sadegh Malvajerd, S.; Izadi, Z. Engineered Solid Lipid Nanoparticles and Nanostructured Lipid Carriers as New Generations of Blood–Brain Barrier Transmitters. ACS Chem. Neurosci. 2021, 12, 4475–4490. [Google Scholar] [CrossRef]
- Dal Magro, R.; Ornaghi, F.; Cambianica, I.; Beretta, S.; Re, F.; Musicanti, C.; Rigolio, R.; Donzelli, E.; Canta, A.; Ballarini, E.; et al. ApoE-Modified Solid Lipid Nanoparticles: A Feasible Strategy to Cross the Blood-Brain Barrier. J. Control. Release 2017, 249, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, P.; Mahjub, R.; Mohammadi, M.; Derakhshandeh, K.; Ghaleiha, A.; Mahboobian, M.M. Pharmacokinetics and Brain Distribution Studies of Perphenazine-Loaded Solid Lipid Nanoparticles. Drug Dev. Ind. Pharm. 2021, 47, 146–152. [Google Scholar] [CrossRef]
- Zheng, W.; Han, X.; Han, B.; Li, G.; Gan, J.; Wang, T.; Xu, B.; He, J.; Du, W.; Cao, X.; et al. LAR Downregulation Protects the Astrocytic U251 and Cocultured SH-SY5Y Cells in a Rotenone-Induced Parkinson’s Disease Cell Model. Int. J. Mol. Sci. 2023, 24, 11111. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Gutiérrez, M.T.; Serrano-García, N.; Orozco-Ibarra, M. Rotenone-Induced Model of Parkinson’s Disease: Beyond Mitochondrial Complex I Inhibition. Mol. Neurobiol. 2023, 60, 1929–1948. [Google Scholar] [CrossRef] [PubMed]
- Tikhonova, M.A.; Tikhonova, N.G.; Tenditnik, M.V.; Ovsyukova, M.V.; Akopyan, A.A.; Dubrovina, N.I.; Amstislavskaya, T.G.; Khlestkina, E.K. Effects of Grape Polyphenols on the Life Span and Neuroinflammatory Alterations Related to Neurodegenerative Parkinson Disease-like Disturbances in Mice. Molecules 2020, 25, 5339. [Google Scholar] [CrossRef] [PubMed]
- Tambe, M.A.; de Rus Jacquet, A.; Strathearn, K.E.; Hensel, J.A.; Colón, B.D.; Chandran, A.; Yousef, G.G.; Grace, M.H.; Ferruzzi, M.G.; Wu, Q.; et al. Protective Effects of Polyphenol-Rich Extracts against Neurotoxicity Elicited by Paraquat or Rotenone in Cellular Models of Parkinson’s Disease. Antioxidants 2023, 12, 1463. [Google Scholar] [CrossRef]
- Fandy, S.J. Grape seed extract’s neuroprotective effect and parkinson’s disease: A systematic review. Malang Neurol. J. 2023, 9, 44–51. [Google Scholar] [CrossRef]
- Trapani, A.; Guerra, L.; Corbo, F.; Castellani, S.; Sanna, E.; Capobianco, L.; Monteduro, A.G.; Manno, D.E.; Mandracchia, D.; Di Gioia, S.; et al. Cyto/Biocompatibility of Dopamine Combined with the Antioxidant Grape Seed-Derived Polyphenol Compounds in Solid Lipid Nanoparticles. Molecules 2021, 26, 916. [Google Scholar] [CrossRef]
- Kon, T.; Tomiyama, M.; Wakabayashi, K. Neuropathology of Lewy Body Disease: Clinicopathological Crosstalk between Typical and Atypical Cases. Neuropathology 2020, 40, 30–39. [Google Scholar] [CrossRef]
- Sian-Hulsmann, J.; Monoranu, C.; Strobel, S.; Riederer, P. Lewy Bodies: A Spectator or Salient Killer? CNS Neurol. Disord.-Drug Targets 2015, 14, 947–955. [Google Scholar] [CrossRef]
- Chartier, S.; Duyckaerts, C. Is Lewy Pathology in the Human Nervous System Chiefly an Indicator of Neuronal Protection or of Toxicity? Cell Tissue Res. 2018, 373, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Won, S.J.; Fong, R.; Butler, N.; Sanchez, J.; Zhang, Y.; Wong, C.; Tambou Nzoutchoum, O.; Huynh, A.; Pan, J.; Swanson, R.A. Neuronal Oxidative Stress Promotes α-Synuclein Aggregation in Vivo. Antioxidants 2022, 11, 2466. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Perera, G.; Bhadbhade, M.; Halliday, G.M.; Dzamko, N. Autophagy Activation Promotes Clearance of α-Synuclein Inclusions in Fibril-Seeded Human Neural Cells. J. Biol. Chem. 2019, 294, 14241–14256. [Google Scholar] [CrossRef] [PubMed]
- Xilouri, M.; Brekk, O.R.; Stefanis, L. Alpha-Synuclein and Protein Degradation Systems: A Reciprocal Relationship. Mol. Neurobiol. 2013, 47, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.-C.; Liu, P.-F.; Chang, C.-H.; Lin, Y.-C.; Chen, Y.-J.; Shu, C.-W. The Interplay of Autophagy and Oxidative Stress in the Pathogenesis and Therapy of Retinal Degenerative Diseases. Cell Biosci. 2022, 12, 1. [Google Scholar] [CrossRef]
- Zhu, Q.; Zhuang, X.; Chen, J.; Yuan, N.; Chen, Y.; Cai, C.; Tan, J.; Su, H.; Lu, J. Lycorine, a Natural Alkaloid, Promotes the Degradation of Alpha-Synuclein via PKA-Mediated UPS Activation in Transgenic Parkinson’s Disease Models. Phytomedicine 2021, 87, 153578. [Google Scholar] [CrossRef] [PubMed]
- Fowler, A.J.; Moussa, C.E.-H. Activating Autophagy as a Therapeutic Strategy for Parkinson’s Disease. CNS Drugs 2018, 32, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Illes-Toth, E.; Rempel, D.L.; Gross, M.L. Exploration of Resveratrol as a Potent Modulator of α-Synuclein Fibril Formation. ACS Chem. Neurosci. 2024, 15, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Chau, E.; Kim, H.; Shin, J.; Martinez, A.; Kim, J.R. Inhibition of Alpha-Synuclein Aggregation by AM17, a Synthetic Resveratrol Derivative. Biochem. Biophys. Res. Commun. 2021, 574, 85–90. [Google Scholar] [CrossRef]
- Gallardo-Fernández, M.; Hornedo-Ortega, R.; Cerezo, A.B.; Troncoso, A.M.; Garcia-Parrilla, M.C. Hydroxytyrosol and Dopamine Metabolites: Anti-Aggregative Effect and Neuroprotective Activity against α-Synuclein-Induced Toxicity. Food Chem. Toxicol. 2023, 171, 113542. [Google Scholar] [CrossRef]
- Abdel-Salam, O.M.; El-Shamarka, M.E.S.; Omara, E.A. Grape Seed Extract Exerts an Anti-Apoptotic Effect and Attenuates the Decrease in Striatal Tyrosine Hydroxylase in Rotenone-Treated Mice. React. Oxyg. Species 2019, 7, 30–46. [Google Scholar] [CrossRef]
- Shrotriya, S.; Deep, G.; Lopert, P.; Patel, M.; Agarwal, R.; Agarwal, C. Grape Seed Extract Targets Mitochondrial Electron Transport Chain Complex III and Induces Oxidative and Metabolic Stress Leading to Cytoprotective Autophagy and Apoptotic Death in Human Head and Neck Cancer Cells. Mol. Carcinog. 2015, 54, 1734–1747. [Google Scholar] [CrossRef]
- Carta, F.; Vullo, D.; Maresca, A.; Scozzafava, A.; Supuran, C.T. Mono-/Dihydroxybenzoic Acid Esters and Phenol Pyridinium Derivatives as Inhibitors of the Mammalian Carbonic Anhydrase Isoforms I, II, VII, IX, XII and XIV. Bioorg. Med. Chem. 2013, 21, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Cometa, S.; Bonifacio, M.A.; Trapani, G.; Di Gioia, S.; Dazzi, L.; De Giglio, E.; Trapani, A. In Vitro Investigations on Dopamine Loaded Solid Lipid Nanoparticles. J. Pharm. Biomed. Anal. 2020, 185, 113257. [Google Scholar] [CrossRef] [PubMed]
- Denora, N.; Laquintana, V.; Trapani, A.; Lopedota, A.; Latrofa, A.; Gallo, J.M.; Trapani, G. Translocator Protein (TSPO) Ligand−Ara-C (Cytarabine) Conjugates as a Strategy to Deliver Antineoplastic Drugs and to Enhance Drug Clinical Potential. Mol. Pharm. 2010, 7, 2255–2269. [Google Scholar] [CrossRef] [PubMed]
- Trapani, A.; Laquintana, V.; Lopedota, A.; Franco, M.; Latrofa, A.; Talani, G.; Sanna, E.; Trapani, G.; Liso, G. Evaluation of New Propofol Aqueous Solutions for Intravenous Anesthesia. Int. J. Pharm. 2004, 278, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Greco, M.; Spinelli, C.C.; De Riccardis, L.; Buccolieri, A.; Di Giulio, S.; Musarò, D.; Pagano, C.; Manno, D.; Maffia, M. Copper Dependent Modulation of A-synuclein Phosphorylation in Differentiated Shsy5y Neuroblastoma Cells. Int. J. Mol. Sci. 2021, 22, 2038. [Google Scholar] [CrossRef] [PubMed]
- Nozhat, Z.; Khalaji, M.S.; Hedayati, M.; Kia, S.K. Different Methods for Cell Viability and Proliferation Assay: Essential Tools in Pharmaceutical Studies. Anti-Cancer Agents Med. Chem. 2022, 22, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Keston, A.S.; Brandt, R. The Fluorometric Analysis of Ultramicro Quantities of Hydrogen Peroxide. Anal. Biochem. 1965, 11, 1–5. [Google Scholar] [CrossRef]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the Probe 2′,7′-Dichlorofluorescin as an Indicator of Reactive Oxygen Species Formation and Oxidative Stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | Size (nm) | PDI a | Zeta Potential (mV) | A.E. DA (%) | A.E. GSE (%) | A.E. FITC (%) |
|---|---|---|---|---|---|---|
| DA-co-GSE-SLNs | 187 ± 4 ** | 0.49 ± 0.04 | −4.1 ± 0.1 ** | 62 ± 4 | 10 ± 0 | - |
| GSE-ads-DA-SLNs | 287 ± 15 ** | 0.53 ± 0.01 | −7.8 ± 0.4 ** | 65 ± 6 | 57 ± 8 | - |
| DA-FITC-co-GSE-SLNs | 297 ± 25 ** | 0.59 ± 0.04 | −3.6 ± 0.1 ** | 82 ± 4 | 23 ± 1 | 94 ± 6 |
| GSE-ads-DA-FITC-SLNs | 266 ± 12 ** | 0.57± 0.04 | −8.1 ± 0.1 ** | 35 ± 1 | 42 ± 2 | 96 ± 1 |
| Plain-SLNs b | 141 ± 11 | 0.35 ± 0.17 | −9.7 ± 0.8 | - | - | - |
| CTR | DA-co-GSE-SLNs | GSE-ads-DA-SLNs | DA-Unloaded-GSE-SLNs | GSE-Unloaded-DA-SLNs | Plain-SLNs | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SLN volume (µL) | - | - | - | 30 | 15 | 10 | 30 | 15 | 10 | 30 | 15 | 10 | 30 | 15 | 10 | 30 | 15 | 10 |
| [DA] (µM) | - | - | - | 150 | 75 | 50 | 150 | 75 | 50 | - | - | - | 150 | 75 | 50 | - | - | - |
| GSE amount (µg/mL) | - | - | - | 12 | 6 | 4 | 0.02 | 0.01 | 0.0067 | 12.6 | 6.3 | 4.2 | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mallamaci, R.; Musarò, D.; Greco, M.; Caponio, A.; Castellani, S.; Munir, A.; Guerra, L.; Damato, M.; Fracchiolla, G.; Coppola, C.; et al. Dopamine- and Grape-Seed-Extract-Loaded Solid Lipid Nanoparticles: Interaction Studies between Particles and Differentiated SH-SY5Y Neuronal Cell Model of Parkinson’s Disease. Molecules 2024, 29, 1774. https://doi.org/10.3390/molecules29081774
Mallamaci R, Musarò D, Greco M, Caponio A, Castellani S, Munir A, Guerra L, Damato M, Fracchiolla G, Coppola C, et al. Dopamine- and Grape-Seed-Extract-Loaded Solid Lipid Nanoparticles: Interaction Studies between Particles and Differentiated SH-SY5Y Neuronal Cell Model of Parkinson’s Disease. Molecules. 2024; 29(8):1774. https://doi.org/10.3390/molecules29081774
Chicago/Turabian StyleMallamaci, Rosanna, Debora Musarò, Marco Greco, Antonello Caponio, Stefano Castellani, Anas Munir, Lorenzo Guerra, Marina Damato, Giuseppe Fracchiolla, Chiara Coppola, and et al. 2024. "Dopamine- and Grape-Seed-Extract-Loaded Solid Lipid Nanoparticles: Interaction Studies between Particles and Differentiated SH-SY5Y Neuronal Cell Model of Parkinson’s Disease" Molecules 29, no. 8: 1774. https://doi.org/10.3390/molecules29081774
APA StyleMallamaci, R., Musarò, D., Greco, M., Caponio, A., Castellani, S., Munir, A., Guerra, L., Damato, M., Fracchiolla, G., Coppola, C., Cardone, R. A., Rashidi, M., Tardugno, R., Sergio, S., Trapani, A., & Maffia, M. (2024). Dopamine- and Grape-Seed-Extract-Loaded Solid Lipid Nanoparticles: Interaction Studies between Particles and Differentiated SH-SY5Y Neuronal Cell Model of Parkinson’s Disease. Molecules, 29(8), 1774. https://doi.org/10.3390/molecules29081774