
Improving the Quality of Wheat Flour Bread by a Thermophilic Xylanase with Ultra Activity and Stability Reconstructed by Ancestral Sequence and Computational-Aided Analysis

 ,
,
Abstract
:1. Introduction
2. Results
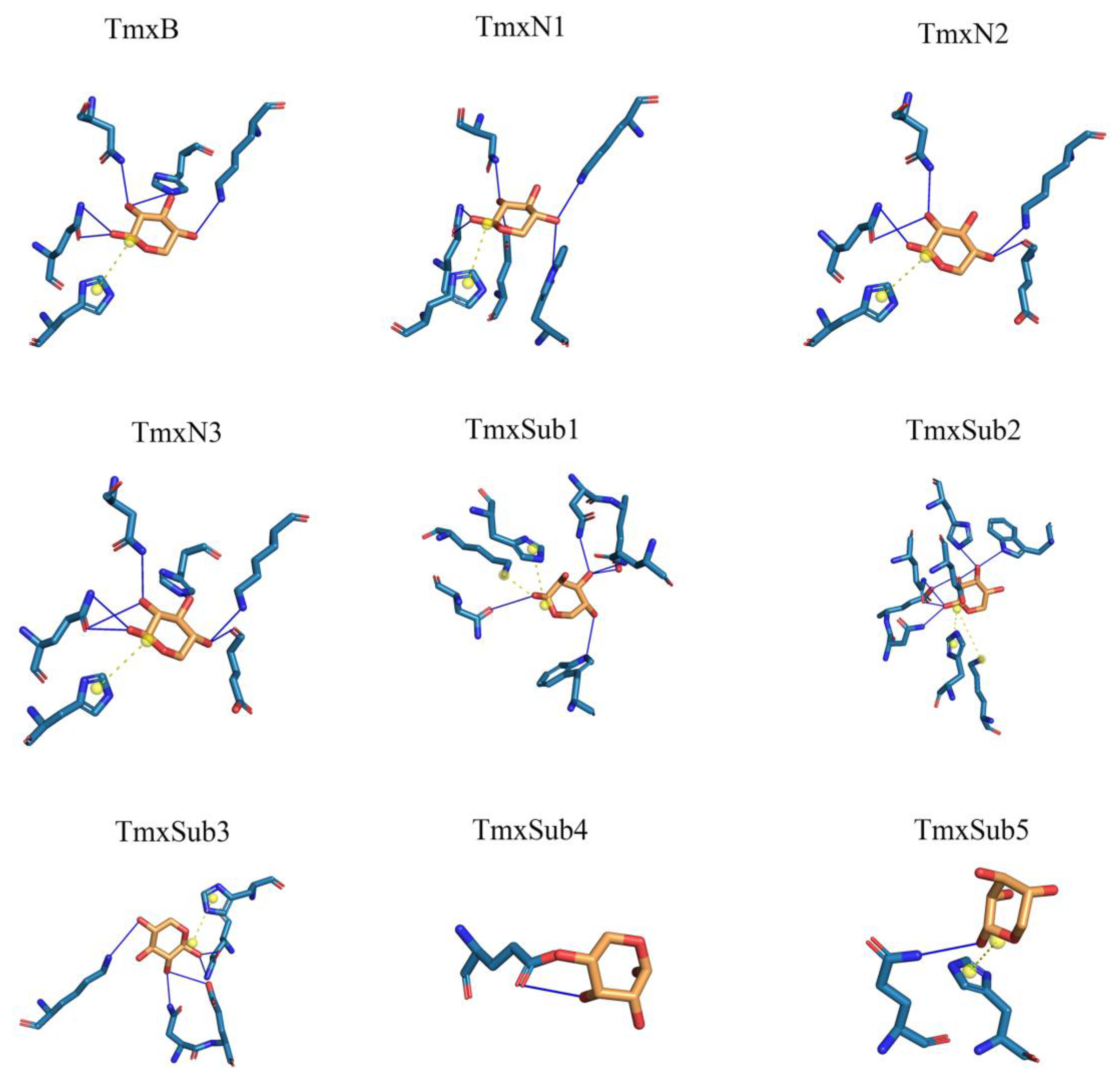
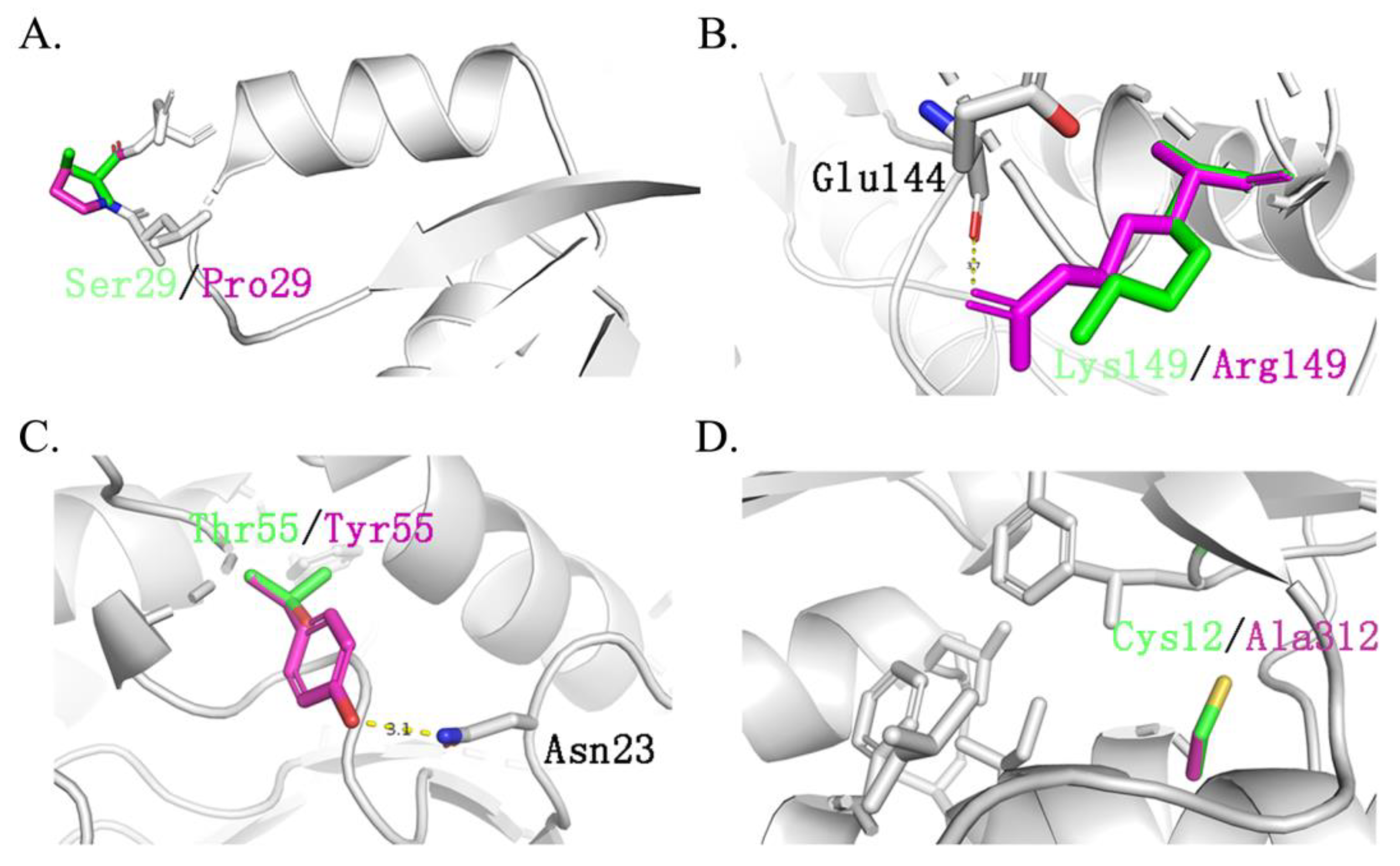
2.1. Inferring the Ancestral Sequences Based on the Evolutionary Branches and Molecular Dynamic Analysis
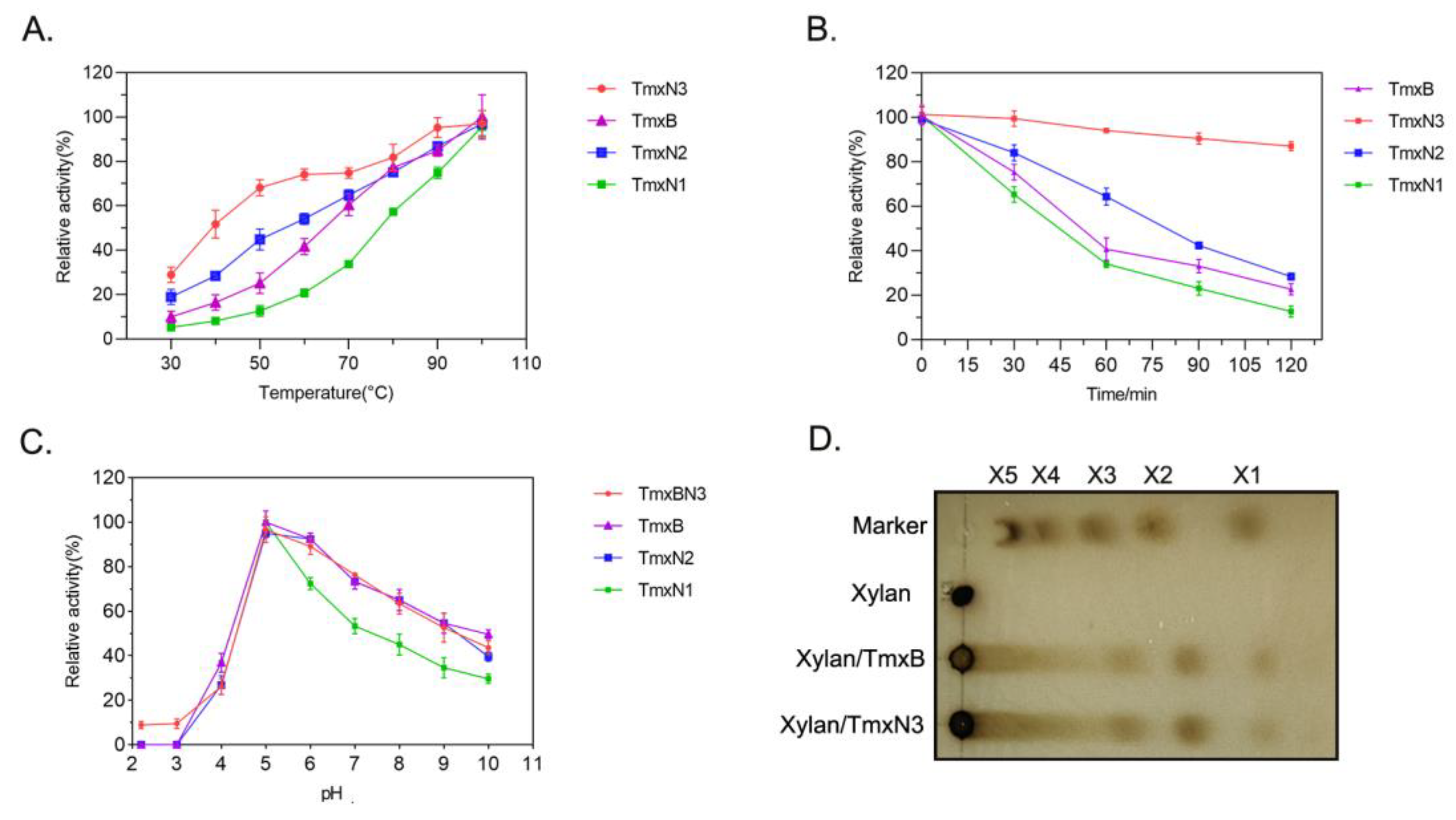
2.2. Ancestral Xylanases Exhibited Significant Improvement in Thermal Stability and Activity
2.3. The Expression Level of Xylanases Improved by Increasing the Gene Dosage in Host Genome
2.4. Bioreactor Production of Thermalphilic Xylanase by High-Density Cultivation
2.5. Thermophilic Xylanases Significantly Improved the Texture of Bread
3. Discussion
3.1. Ancestral Sequences Reconstruction and Molecular Dynamics Analysis Improved the Activity and Thermostability of Xylanase
3.2. Xylanase TmxN3 Exhibited Excellent Thermal Stability
3.3. The Thermophilic Xylanases Effectively Improve Wheat Bread Quality
4. Materials and Methods
4.1. Constructing a Phylogenetic Tree of Xylanases and Ancestral Sequences Mining
4.2. Molecular Dynamics Analysis of Xylanase and the Ligand
4.3. Recombinants Construction and Expression of the Xylanases
4.4. Kinetic Parameters Determination and Thermal Stable Characterization
4.5. High-Density Cultivation of Recombinant Strain and Xylanase Expression in a 5L Bioreactor
4.6. Wheat Bread Making and Quality Assessment
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santibáñez, L.; Henríquez, C.; Corro-Tejeda, R.; Bernal, S.; Armijo, B.; Salazar, O. Xylooligosaccharides from lignocellulosic biomass: A comprehensive review. Carbohydr. Polym. 2021, 251, 117118. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, M.; Barroca, M.; Collins, T. Endo-1, 4-β-xylanase-containing glycoside hydrolase families: Characteristics, singularities and similarities. Biotechnol. Adv. 2023, 65, 108148. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Han, Z. Understanding the positional binding and substrate interaction of a highly thermostable GH10 xylanase from Thermotoga maritima by molecular docking. Biomolecules 2018, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, P.; Mahajan, R. Cellulase and xylanase synergism in industrial biotechnology. Appl. Microbiol. Biotechnol. 2019, 103, 8711–8724. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ma, T.; Shang-Guan, F.; Han, Z. Improving the catalytic activity of thermostable xylanase from Thermotoga maritima via mutagenesis of non-catalytic residues at glycone subsites. Enzym. Microb. Technol. 2020, 139, 109579. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Dangi, A.K.; Shukla, P. Engineering thermostable microbial xylanases toward its industrial applications. Mol. Biotechnol. 2018, 60, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Golgeri, M.D.B.; Mulla, S.I.; Bagewadi, Z.K.; Tyagi, S.; Hu, A.; Sharma, S.; Bilal, M.; Bharagava, R.N.; Ferreira, L.F.R.; Gurumurthy, D.M.; et al. A systematic review on potential microbial carbohydrases: Current and future perspectives. Crit. Rev. Food Sci. Nutr. 2024, 64, 438–455. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Arif, S.; Khurshid, S.; Iqbal, H.M.; Akbar, Q.; Ali, T.M.; Mohiuddin, S. A combined use of different functional additives for improvement of wheat flour quality for bread making. J. Sci. Food Agric. 2023, 103, 3261–3271. [Google Scholar] [CrossRef]
- Zerva, A.; Pentari, C.; Ferousi, C.; Nikolaivits, E.; Karnaouri, A.; Topakas, E. Recent advances on key enzymatic activities for the utilisation of lignocellulosic biomass. Bioresour. Technol. 2021, 342, 126058. [Google Scholar] [CrossRef]
- Yadav, P.; Maharjan, J.; Korpole, S.; Prasad, G.S.; Sahni, G.; Bhattarai, T.; Sreerama, L. Production, purification, and characterization of thermostable alkaline xylanase from Anoxybacillus kamchatkensis NASTPD13. Front. Bioeng. Biotechnol. 2018, 6, 65. [Google Scholar] [CrossRef]
- Bhat, S.K.; Purushothaman, K.; Kini, K.R.; Gopala Rao Appu Rao, A.R. Design of mutants of GH11 xylanase from Bacillus pumilus for enhanced stability by amino acid substitutions in the N-terminal region: An in silico analysis. J. Biomol. Struct. Dyn. 2022, 40, 7666–7679. [Google Scholar] [CrossRef] [PubMed]
- Bello, A.; Giménez-Rico, R.D.; Gilani, S.; Hillen, B.C.; Venter, K.M.; Plumstead, P.; Dersjant-Li, Y. Application of enzyme matrix values for energy and nutrients to a wheat-corn-soybean meal-based broiler diet supplemented with a novel phytase, with or without a xylanase–β-glucanase, achieved a production benefit over a nutritionally adequate unsupplemented diet. Poult. Sci. 2023, 102, 103131. [Google Scholar] [PubMed]
- Kaushal, J.; Khatri, M.; Singh, G.; Arya, S.K. A multifaceted enzyme conspicuous in fruit juice clarification: An elaborate review on xylanase. Int. J. Biol. Macromol. 2021, 193, 1350–1361. [Google Scholar] [CrossRef] [PubMed]
- Spence, M.A.; Kaczmarski, J.A.; Saunders, J.W.; Jackson, C.J. Ancestral sequence reconstruction for protein engineers. Curr. Opin. Struct. Biol. 2021, 69, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Dou, Z.; Luo, T.; Sun, Z.; Ma, H.; Xu, G.; Ni, Y. Directed reconstruction of a novel ancestral alcohol dehydrogenase featuring shifted pH-profile, enhanced thermostability and expanded substrate spectrum. Bioresour. Technol. 2022, 363, 127886. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Zhao, L.; Hou, Y.; Yue, C.; Liu, P.; Zheng, Y.; Peng, W.; Yang, J. An Inferred Ancestral CotA Laccase with Improved Expression and Kinetic Efficiency. Int. J. Mol. Sci. 2023, 24, 10901. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Zhou, Y.; Yi, Z.; Zhou, R.; Jin, W.; Zhang, G. Highly thermostable and promiscuous β-1, 3-xylanasen designed by optimized ancestral sequence reconstruction. Bioresour. Technol. 2021, 340, 125732. [Google Scholar] [CrossRef]
- Siddiq, M.A.; Hochberg, G.K.; Thornton, J.W. Evolution of protein specificity: Insights from ancestral protein reconstruction. Curr. Opin. Struct. Biol. 2017, 47, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, G.K.; Liu, Y.; Marklund, E.G.; Metzger, B.P.; Laganowsky, A.; Thornton, J.W. A hydrophobic ratchet entrenches molecular complexes. Nature 2020, 588, 503–508. [Google Scholar] [CrossRef]
- Singh, A.; Vanga, S.K.; Orsat, V.; Raghavan, V. Application of molecular dynamic simulation to study food proteins: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 2779–2789. [Google Scholar] [CrossRef]
- Scossa, F.; Fernie, A.R. Ancestral sequence reconstruction-An underused approach to understand the evolution of gene function in plants? Comput. Struct. Biotechnol. J. 2021, 19, 1579–1594. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Mu, T.; Sun, H.; Zhou, L. Evaluation of texture, retrogradation enthalpy, water mobility, and anti-staling effects of enzymes and hydrocolloids in potato steamed bread. Food Chem. 2022, 368, 130686. [Google Scholar] [CrossRef] [PubMed]
- Kamilari, E.; Stanton, C.; Reen, F.J.; Ross, R.P. Uncovering the biotechnological importance of Geotrichum candidum. Foods 2023, 12, 1124. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.L.; Evans, P.N.; Li, Y.X.; Rao, Y.Z.; Qu, Y.N.; Tan, S.; Jiao, J.Y.; Chen, Y.T.; Hedlund, B.P.; Shu, W.S.; et al. Comparative genomics reveals thermal adaptation and a high metabolic diversity in “Candidatus Bathyarchaeia”. Msystems 2021, 6, e0025221. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Fang, C.; Wang, Q.; Zhou, Y.; Zhang, G.; Ma, Y. High-level expression of improved thermo-stable alkaline xylanase variant in Pichia pastoris through codon optimization, multiple gene insertion and high-density fermentation. Sci. Rep. 2016, 6, 37869. [Google Scholar] [CrossRef] [PubMed]
- Wongwisansri, S.; Promdonkoy, P.; Matetaviparee, P.; Roongsawang, N.; Eurwilaichitr, L.; Tanapongpipat, S. High-level production of thermotolerant β-xylosidase of Aspergillus sp. BCC125 in Pichia pastoris: Characterization and its application in ethanol production. Bioresour. Technol. 2013, 132, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, J.; Bao, C.; Dong, B.; Cao, Y. Characterization of a novel GH10 xylanase with a carbohydrate binding module from Aspergillus sulphureus and its synergistic hydrolysis activity with cellulase. Int. J. Biol. Macromol. 2021, 182, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Morgan, N.; Bhuiyan, M.; Hopcroft, R. Non-starch polysaccharide degradation in the gastrointestinal tract of broiler chickens fed commercial-type diets supplemented with either a single dose of xylanase, a double dose of xylanase, or a cocktail of non-starch polysaccharide-degrading enzymes. Poult. Sci. 2022, 101, 101846. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, L.; Jiang, Z.; Liu, L.; Wang, J.; Zhong, L.; Yang, T.; Zhou, Q.; Dong, W.; Zhou, J.; et al. A novel cold-active GH8 xylanase from cellulolytic myxobacterium and its application in food industry. Food Chem. 2022, 393, 133463. [Google Scholar] [CrossRef]
- de Queiroz Brito Cunha, C.C.; Gama, A.R.; Cintra, L.C.; Bataus, L.A.M.; Ulhoa, C.J. Improvement of bread making quality by supplementation with a recombinant xylanase produced by Pichia pastoris. PLoS ONE 2018, 13, e0192996. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, C.; Yang, M.; Ou, Z.; Lin, Y.; Zhao, F.; Han, S. Characterization and application of a novel xylanase from Halolactibacillus miurensis in wholewheat bread making. Front. Bioeng. Biotechnol. 2022, 10, 1018476. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Wu, G.; Wu, H. Biochemical characterization of a GH10 xylanase from the anaerobic rumen fungus Anaeromyces robustus and application in bread making. 3 Biotech 2021, 11, 406. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKO ALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.; Bernstein, Y.; Williams, H.E. Clustered sequence representation for fast homology search. J. Comput. Biol. 2007, 14, 594–614. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Moshe, A.; Pupko, T. Ancestral sequence reconstruction: Accounting for structural information by averaging over replacement matrices. Bioinformatics 2019, 35, 2562–2568. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Su, H.; Wang, W.; Ye, L.; Wei, H.; Peng, Z.; Anishchenko, I.; Baker, D.; Yang, J. The trRosetta server for fast and accurate protein structure prediction. Nat. Protoc. 2021, 16, 5634–5651. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4, Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein-ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef]
- Gupteshwar Gupta, G.G.; Vikram Sahai, V.S.; Gupta, R.K. Thermal stability and thermodynamics of xylanase from Melanocarpus albomyces in presence of polyols and salts. BioResources 2014, 9, 5801–5816. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H-Bond Number | H-Bond Min-Distance | H-Bond Mean-Distance | Salt Bridge Number | Salt Bridge Min-Distance | |
|---|---|---|---|---|---|
| TmxB | 6 | 2.79 | 3.28 | 1 | 4.47 |
| TmxN1 | 6 | 2.86 | 3.26 | 1 | 4.46 |
| TmxN2 | 5 | 3.07 | 3.55 | 1 | 4.47 |
| TmxN3 | 7 | 2.79 | 3.35 | 1 | 4.46 |
| TmxSub3 | 5 | 2.48 | 3.17 | 1 | 4.59 |
| TmxSub2 | 7 | 2.81 | 2.97 | 2 | 4.8 |
| TmxSub1 | 5 | 2.58 | 3.23 | 2 | 3.85 |
| TmxSub4 | 1 | 3.19 | 3.19 | 0 | - |
| TmxSub5 | 1 | 3.70 | 3.70 | 1 | 4.93 |
| Enzyme | Vmax (U/mg) | Km (mg/mL) | Kcat (s−1) | Kcat/Km (mL/s/mg) |
|---|---|---|---|---|
| TmxB | 17,019 | 0.82 | 4758 | 5468 |
| TmxN3 | 45,827 | 0.80 | 7986 | 9367 |
| TmxN2 | 2025 | 0.69 | 3470 | 5028 |
| TmxN1 | 1959 | 1.00 | 2163 | 2160 |
| Xylanase | Hardness (g) | Chewiness (g × mm) | Gumminess | Cohesiveness | Springiness (mm) | Resilience |
|---|---|---|---|---|---|---|
| Control (No) | 508.28 ± 39.56 c | 370.07 ± 70.35 c | 418.69 ± 71.82 c | 0.81 ± 0.08 a | 0.88 ± 0.02 a | 0.07 ± 0.00 a |
| 1 d | 563.25 ± 42.22 b | 498.05 ± 24.78 d | 552.95 ± 51.82 d | 0.85 ± 0.032 d | 0.75 ± 0.042 a | 0.059 ± 0.012 a |
| 7 d | 3701.48 ± 20.33 e | 3184.67 ± 19.80 f | 4421.41 ± 43.45 f | 0.66 ± 0.020 d | 0.65 ± 0.017 c | 0.02 a |
| 10 d | 3905.47 ± 47.36 e | 3579.77 ± 11.75 e | 4729.76 ± 18.21 e | 0.61 ± 0.015 c | 0.65 ± 0.029 d | 0.01 a |
| TmxB (1.5 mg/kg) | 454.82 ± 38.43 bc | 335.82 ± 68.16 c | 380.04 ± 69.18 c | 0.83 ± 0.09 a | 0.88 ± 0.02 a | 0.08 a |
| 1 d | 504.66 ± 37.58 b | 352.66 ± 15.71 c | 478.46 ± 28.43 c | 0.66 ± 0.0226 a | 0.76 ± 0.043 a | 0.056 ± 0.012 a |
| 7 d | 2010.62 ± 37.42 d | 1479.08 ± 38.14 e | 1828.88 ± 37.24 e | 0.44 ± 0.025 | 0.45 ± 0.04 a | 0.02 a |
| 10 d | 2682.96 ± 41.48 d | 3412.97 ± 23.26 d | 4541.45 ± 73.01 d | 0.43 ± 0.042 ab | 0.44 ± 0.01 b | 0.01 a |
| TmxB (4.5 mg/kg) | 401.07 ± 44.13 b | 296.81 ± 66.45 bc | 335.07 ± 66.78 bc | 0.82 ± 0.08 a | 0.88 ± 0.02 a | 0.08 ± 0.01 a |
| 1 d | 505.46 ± 50.062 b | 325.06 ± 19.58 c | 408.76 ± 41.7 b | 0.76 ± 0.022 b | 0.76 ± 0.042 a | 0.06 ± 0.012 a |
| 7 d | 1680.29 ± 49.59 c | 1316.77 ± 33.44 d | 1558.10 ± 43.20 d | 0.45 ± 0.012 ab | 0.47 ± 0.012 a | 0.02 a |
| 10 d | 2444.99 ± 29.08 c | 3274.85 ± 27.24 c | 3983.09 ± 61.62 c | 0.44 ± 0.01 ab | 0.49 ± 0.0058 bc | 0.01 a |
| TmxN3 (1.5 mg/kg) | 290.02 ± 40.09 a | 214.21 ± 53.81 ab | 243.64 ± 51.45 ab | 0.83 ± 0.07 a | 0.87 ± 0.04 a | 0.078 a |
| 1 d | 329.26 ± 3.23 a | 243.76 ± 33.202 b | 266.36 ± 20.42 a | 0.76 ± 0.022 ab | 0.76 ± 0.032 a | 329.26 ± 3.23 a |
| 7 d | 1328.42 ± 47.99 b | 1160.25 ± 30.54 c | 1152.6 ± 27.24 c | 0.45 ± 0.023 ab | 0.48 ± 0.011 a | 1328.42 ± 47.99 b |
| 10 d | 1609.21 ± 50.88 b | 2159.16 ± 30.13 b | 2391.89 ± 33.4 b | 0.48 ± 0.017 b | 0.38 ± 0.042 a | 1609.21 ± 50.88 b |
| TmxN3 (3.0 mg/kg) | 230.95 ± 20.01 a | 165.99 ± 38.75 a | 189.08 ± 39.51 a | 0.81 ± 0.11 a | 0.87 ± 0.03 a | 0.08 ± 0.01 a |
| 1 d | 302.26 ± 40.21 a | 191.46 ± 30.48 a | 196.75 ± 38.41 a | 0.76 ± 0.042 ab | 0.86 ± 0.032 ab | 0.06 ± 0.012 a |
| 7 d | 916.4845 ± 13.58 a | 1014.6991 ± 22.72 b | 1048.1843 ± 40.67 b | 0.51 ± 0.055 bc | 0.53 ± 0.025 b | 0.0167 ± 0.0058 a |
| 10 d | 1192.6785 ± 52.21 a | 1927.5211 ± 25.39 a | 2205.6673 ± 41.09 a | 0.41 ± 0.017 a | 0.4767 ± 0.015 bc | 0.01 a |
| TmxN3 (4.5 mg/kg) | 228.64 ± 15.67 a | 177.75 ± 32.36 a | 194.61 ± 32.24 a | 0.84 ± 0.09 a | 0.91 ± 0.02 a | 0.07 ± 0.03 a |
| 1 d | 276.0596 ± 16.88 a | 197.6596 ± 27.86 ab | 232.0596 ± 42.12 a | 0.7596 ± 0.042 b | 0.8596 ± 0.032 b | 0.057 ± 0.0025 a |
| 7 d | 962.9357 ± 27.14 a | 911.1942 ± 35.38 a | 955.4933 ± 58.99 a | 0.5133 ± 0.031 c | 0.68 ± 0.036 c | 0.017 ± 0.0057 a |
| 10 d | 1053.167 ± 14.13 a | 965.3332 ± 33.82 a | 1127.2548 ± 7.44 a | 0.4467 ± 0.031 ab | 0.5267 ± 0.042 bc | 0.01 a |
| Xylanase Family | Taxon | Optimal Temperature (°C) | Optimal pH | Enzyme Amount | Hardness Reduced | Chewiness Reduced | Reference |
|---|---|---|---|---|---|---|---|
| GH10 | Thermotoga maritima | 100 | 5 | 4.5 mg/kg | 55.2% | 40.11% | This study |
| GH11 | Streptomyces sp. S27 | 75 | 6 | 300 U/kg | ND | ND | [30] |
| GH11 | Halolactibacillus miurensis | 45 | 6.5 | 6.0 mg/kg | 34.73% | 31.74 | [31] |
| GH 10 | Anaeromyces robustus | 40 | 5.5 | 75 mg/kg | 31.03% | 34.76% | [32] |
| GH8 | Sorangium cellulosum | 50 | 6 | 0.2 mg/kg | 50.6% | 41.3% | [29] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, G.; Hong, X.; Zhu, M.; Lei, L.; Han, Z.; Meng, Y.; Yang, J. Improving the Quality of Wheat Flour Bread by a Thermophilic Xylanase with Ultra Activity and Stability Reconstructed by Ancestral Sequence and Computational-Aided Analysis. Molecules 2024, 29, 1895. https://doi.org/10.3390/molecules29081895
Hu G, Hong X, Zhu M, Lei L, Han Z, Meng Y, Yang J. Improving the Quality of Wheat Flour Bread by a Thermophilic Xylanase with Ultra Activity and Stability Reconstructed by Ancestral Sequence and Computational-Aided Analysis. Molecules. 2024; 29(8):1895. https://doi.org/10.3390/molecules29081895
Chicago/Turabian StyleHu, Guoshuai, Xizhi Hong, Meixin Zhu, Lei Lei, Zhenggang Han, Yong Meng, and Jiangke Yang. 2024. "Improving the Quality of Wheat Flour Bread by a Thermophilic Xylanase with Ultra Activity and Stability Reconstructed by Ancestral Sequence and Computational-Aided Analysis" Molecules 29, no. 8: 1895. https://doi.org/10.3390/molecules29081895
APA StyleHu, G., Hong, X., Zhu, M., Lei, L., Han, Z., Meng, Y., & Yang, J. (2024). Improving the Quality of Wheat Flour Bread by a Thermophilic Xylanase with Ultra Activity and Stability Reconstructed by Ancestral Sequence and Computational-Aided Analysis. Molecules, 29(8), 1895. https://doi.org/10.3390/molecules29081895