Reactive Oxygen Species Released from Hypoxic Hepatocytes Regulates MMP-2 Expression in Hepatic Stellate Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Preparation of Conditioned Media
2.2. Real-Time RT-PCR
2.3. Western Blot
2.4. Gelatin Zymography for MMP-2
2.5. Quantitative Assay for Reactive Oxygen Species
2.6. Treatment of BRL-3A Cells with Reduced Glutathione
2.7. Statistical Analysis
3. Results
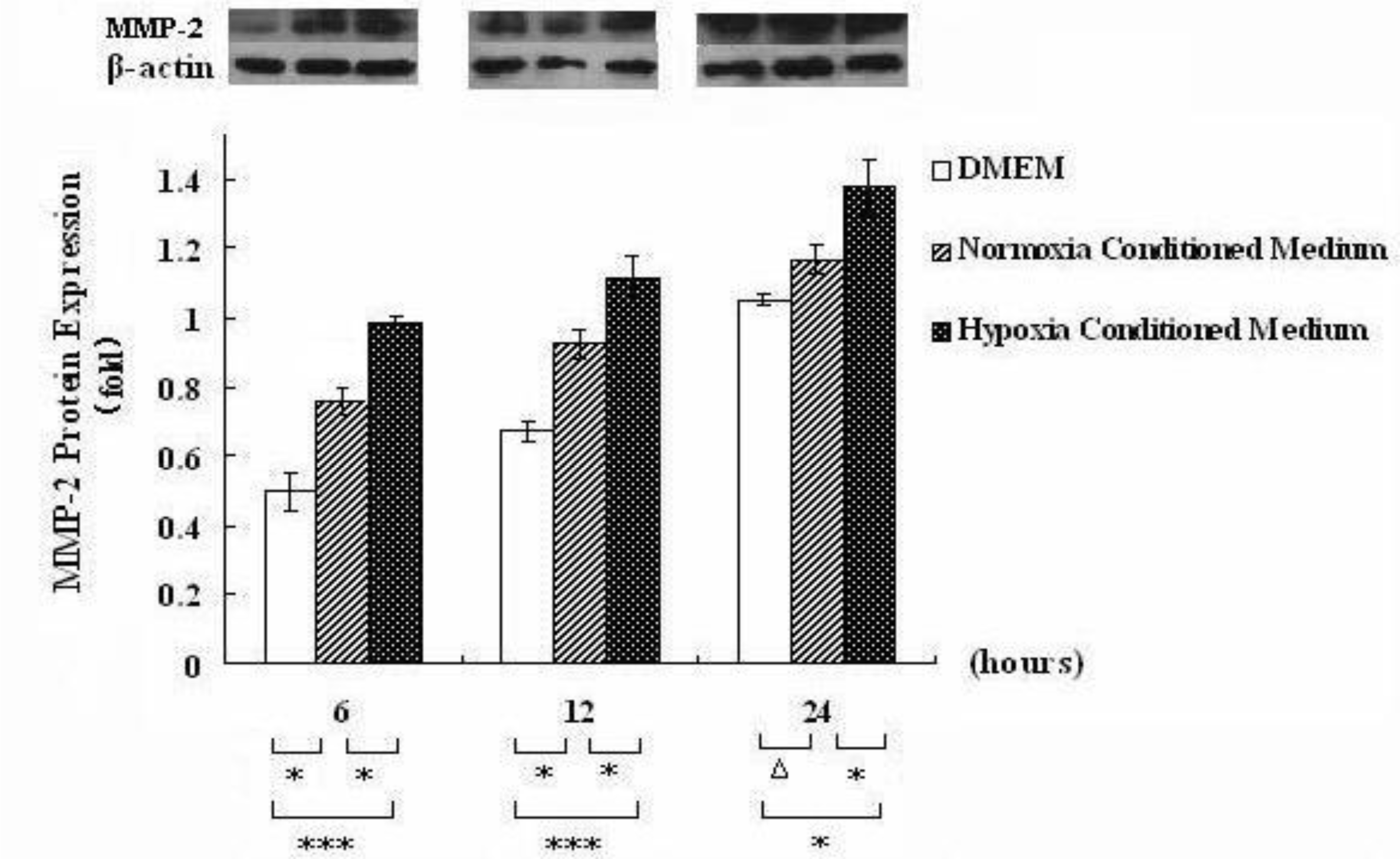
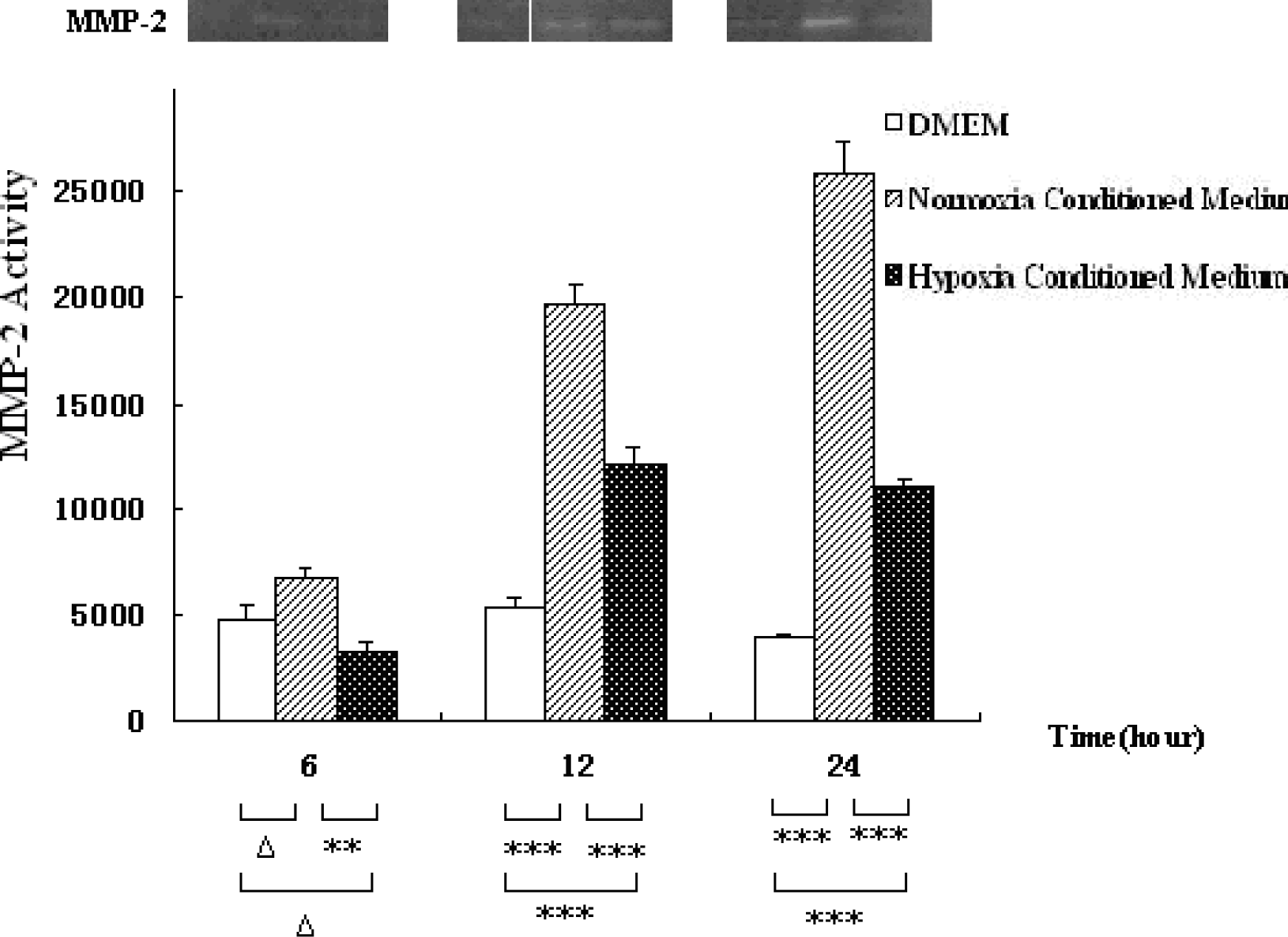
3.1. Hepatocyte Conditioned Medium Upregulates MMP-2 Expression in HSCs
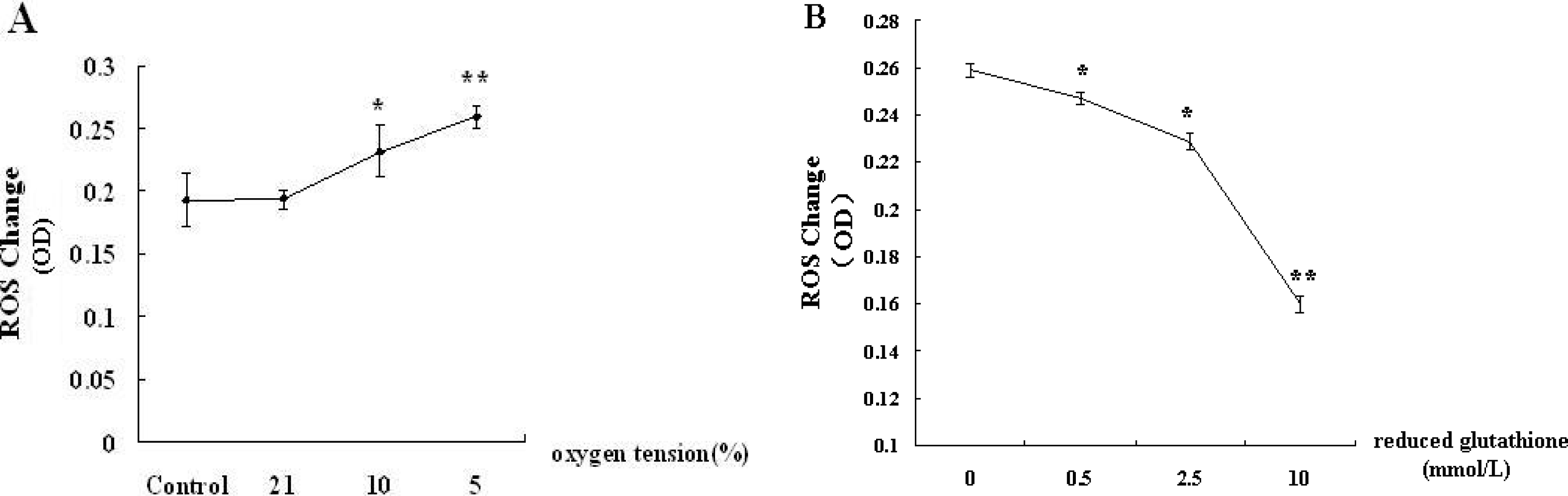
3.2. Reduced Glutathione Antagonizes the Generation of ROS in the Supernatants of Hepatocytes
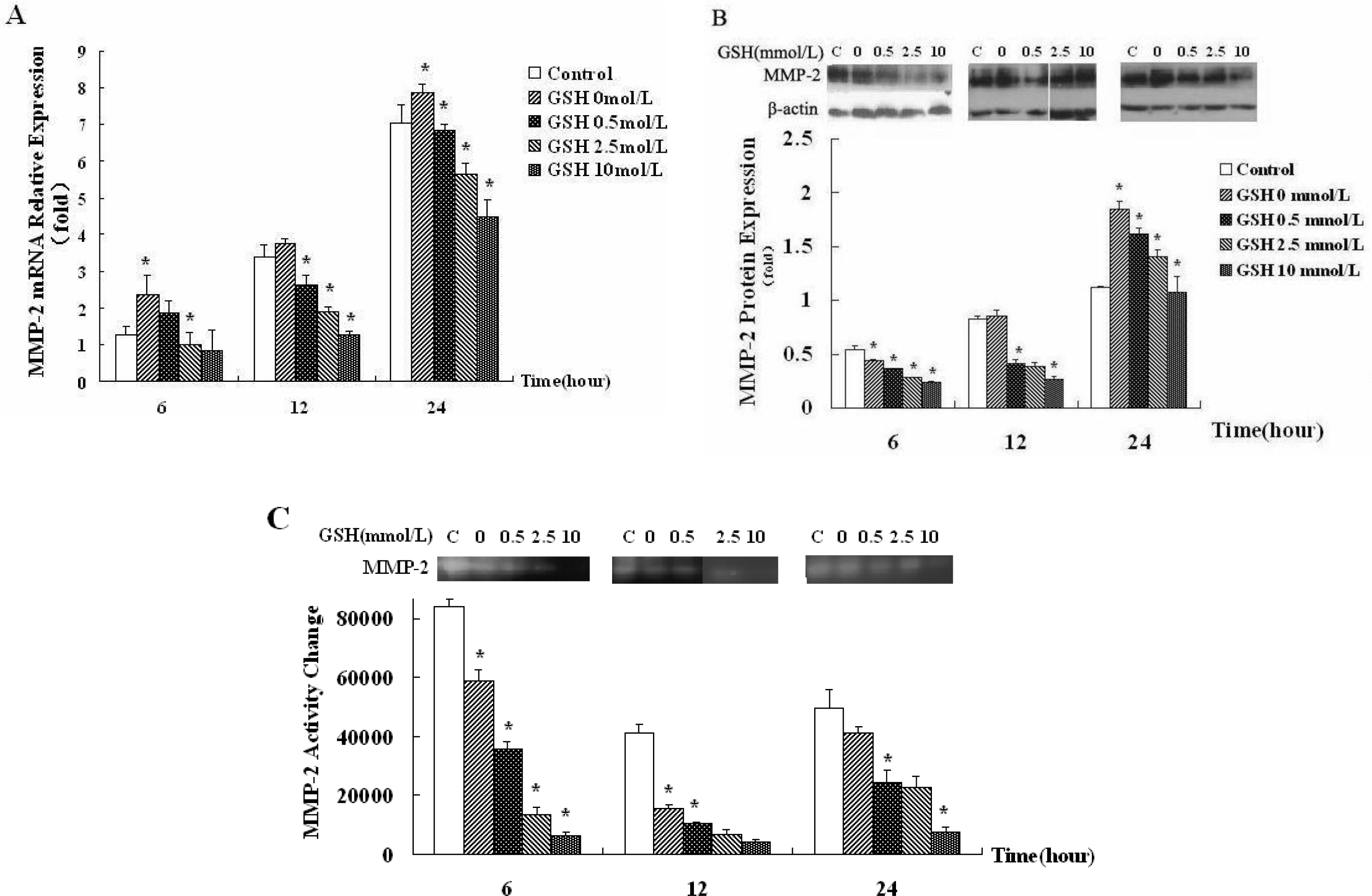
3.3. ROS Generated by Hypoxic Hepatocyte Contributes to the Downregulation of MMP-2 Expression and Activity in HSCs
3.4. Hepatocyte Conditioned Medium Upregulates MMP-2 Expression in HSCs via NF-κB Signaling
4. Discussion
5. Conclusions
Acknowledgments
References
- Bataller, R; Brenner, DA. Liver fibrosis. J. Clin. Invest 2005, 115, 209–218. [Google Scholar]
- Senoo, H. Structure and function of hepatic stellate cells. Med. Electron. Microsc 2004, 37, 3–15. [Google Scholar]
- Agnieszka, SC; Krzysztof, P; Jadwiga, D; Martyna, KS. Zinc supplementation attenuates ethanol-and acetaldehyde-induced liver stellate cell activation by inhibiting reactive oxygen species (ROS) production and by influencing intracellular signaling. Biochem. Pharmacol 2009, 78, 301–314. [Google Scholar]
- Yang, C; Zeisberg, M; Mosterman, B; Sudhakar, A; Yerramalla, U; Holthaus, K. Liver fibrosis: Insights into migration of hepatic stellate cells in response to extracellular matrix and growth factors. Gastroenterology 2003, 124, 147–159. [Google Scholar]
- Parola, M; Marra, F; Pinzani, M. Myofibroblast-like cells and liver fibrogenesis: Emerging concepts in a rapidly moving scenario. Mol. Aspects Med 2008, 29, 58–66. [Google Scholar]
- Schuppan, D; Ruehl, M; Somasundaram, R; Hahn, EG. Matrix as a modulator of hepatic fibrogenesis. Semin. Liver Dis 2001, 21, 351–372. [Google Scholar]
- Arthur, MJP. Fibrogenesis II. Metalloproteinases and their inhibitors in liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol 2000, 279, 245–249. [Google Scholar]
- McCrudden, R; Iredale, JP. Liver fibrosis, the hepatic stellate cell and tissue inhibitors of metalloproteinases. Histol. Histopathol 2000, 15, 1159–1168. [Google Scholar]
- Li, JT; Liao, ZX; Ping, J; Xu, D; Wang, H. Molecular mechanism of hepatic stellate cell activation and antifibrotic therapeutic strategies. J. Gastroenterol 2008, 43, 419–428. [Google Scholar]
- Gardi, C; Arezzini, B; Fortino, V; Comporti, M. Effect of free iron on collagen synthesis, cell proliferation and MMP-2 expression in rat hepatic stellate cells. Biochem. Pharmacol 2002, 64, 1139–1145. [Google Scholar]
- Takahashi, C; Sheng, Z; Horan, TP; Kitayama, H; Maki, M; Hitomi, K. Regulation of matrix metalloproteinase-9 and inhibition of tumor invasion by the membrane-anchored-glycoprotein RECK. Proc. Natl. Acad. Sci. USA 1998, 95, 13221–13226. [Google Scholar]
- Semenza, GL. Life with Oxygen. Science 2007, 318, 62–64. [Google Scholar]
- Hao, XJ; Li, XJ; Du, L. Observation of shenqirougan decection on hepatic fibrosis. J Emerg Traditional Chin Med 2010, 19. [Google Scholar]
- Guo, L; Wang, Z. Effects of ganxinning capsule on super micro-structure of fibrotic liver rats. J Emerg Traditional Chin Med 2010, 19. [Google Scholar]
- Shi, YF; Fong, CC; Zhang, Q; Cheung, PY; Tzang, CH; Wu, RS. Hypoxia induces the activation of human hepatic stellate cells LX-2 through TGF-beta signaling pathway. Febs. Lett 2007, 581, 203–210. [Google Scholar]
- Fan, RH; Chen, PS; Zhao, D; Zhang, WD. Hypoxia induced by CoCl2 influencing the expression and the activity of matrix metalloproteinase-2 in rat hepatic stellate cells. Chin. J. Hepatol 2007, 15, 654–657. [Google Scholar]
- Chen, PS; Zhai, WR; Zhou, XM; Zhang, JS; Zhang, YE; Ling, YQ. Effects of hypoxia on the regulation of expression and activity of matrix metalloproteinase-2 in hepatic stellate cell. World J. Gastroenterol 2001, 7, 647–651. [Google Scholar]
- Zhao, D; Fan, RH; Chen, PS. Effects of hypoxia on expression and activity of matrix metalloproteinase-2 and the mechanism of regulation by hypoxia-inducible factor1α in hepatic stellate cells. Pathol. Int 2007, 57, A24. [Google Scholar]
- Nissen, NN; Polverini, PJ; Koch, AE; Volin, MV; Gamelli, RL; DiPietro, LA. Vascular endothelial growth factor mediates angiogenic activity during the proliferative phase of wound healing. Am. J. Pathol 1998, 152, 1445–1452. [Google Scholar]
- Scheid, A; Wenger, RH; Christina, H; Camenisch, I; Ferenc, A; Stauffer, UG. Hypoxia-regulated gene expression in fetal wound regeneration and adult wound repair. Pediatr. Surg. Int 2000, 16, 232–236. [Google Scholar]
- Zhang, AF; Chen, PS. Effects of hyperbaric oxygen with free-radical antagonists on the expression and activity of matrix metalloproteinase-2 in rat livers. Chin. J. Hepatol 2005, 6, 545–546. [Google Scholar]
- Friedman, SL. Cytokines and fibrogenesis. Semin. Liver Dis 1999, 19, 129–140. [Google Scholar]
- Friedman, SL. Molecular regulation of hepatic fibrosis, an integrated cellular response to tissue injury. J. Biol. Chem 2000, 275, 2247–2250. [Google Scholar]
- Lee, JS; Basalyga, DM; Simionescu, A; Isenburg, JC; Simionescu, DT; Vyavahare, NR. Elastin calcification in the rat subdermal model is accompanied by up-regulation of degradative and osteogenic cellular responses. Am. J. Pathol 2006, 168, 490–498. [Google Scholar]
- Mason, HR; Grove, SD; Rubin, BS; Nowak, RA; Castellot, JJ, Jr. Estrogen induces CCN5 expression in the rat uterus in vivo. Endocrinology 2004, 145, 976–982. [Google Scholar]
- Friedman, SL. Mechanisms of hepatic fibrogenesis. Gastroenterology 2008, 134, 1655–1669. [Google Scholar]
- Yang, C; Zeisberg, M; Mosterman, B; Sudhakar, A; Yerramalla, U; Holthaus, K. Liver fibrosis: Insights into migration of hepatic stellate cells in response to extracellular matrix and growth factors. Gastroenterology 2003, 124, 147–159. [Google Scholar]
- Nelson, KK; Melendez, JA. The powerhouse takes control of the cell: The role of mitochondria in signal transduction. Free Radic. Biol. Med 2004, 37, 768–784. [Google Scholar]
- Visse, R; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res 2003, 92, 827–839. [Google Scholar]
- Oh, J; Takahashi, R; Kondo, S; Mizoguchi, A; Adachi, E; Sasahara, RM; Nishimura, S; Imamura, Y; Kitayama, H; Alexander, DB; et al. The membrane-anchored MMP inhibitor RECK is a key regulator of extracellular matrix integrity and angiogenesis. Cell 2001, 107, 789–800. [Google Scholar]
- Newsome, AL; Johnson, JP; Seipelt, RL; Thompson, MW. Apolactoferrin inhibits the catalytic domain of matrix metalloproteinase-2 by zinc chelation. Biochem. Cell Biol 2007, 85, 563–572. [Google Scholar]
- Han, YH; Park, WH. The changes of reactive oxygen species and glutathione by MG132, a proteasome inhibitor affect As4.1 juxtaglomerular cell growth and death. Chem. Biol. Interact 2010, 184, 319–332. [Google Scholar]
- Tsukada, S; Parsons, CJ; Rippe, RA. Mechanisms of liver fibrosis. Clin. Chim. Acta 2006, 364, 33–60. [Google Scholar]
- Galli, A; Svegliati-Baroni, G; Ceni, E; Milani, S; Ridolfi, F; Salzano, R; Tarocchi, M; Grappone, C; Pellegrini, G; Benedetti, A; et al. Oxidative stress stimulates proliferation and invasiveness of hepatic stellate cells via a MMP-2 mediated mechanism. Hepatology 2005, 41, 1074–1084. [Google Scholar]
- Sariahmetoglu, M; Crawford, BD; Leon, H; Sawicka, J; Li, L; Ballermann, BJ; Holmes, C; Berthiaume, LG; Holt, A; Sawicki, G; Schulzet, R. Regulation of matrix metalloproteinase-2 activity by phosphorylation. FASEB J 2007, 21, 2486–2495. [Google Scholar]
- Belotti, D; Paganoni, P; Manenti, L; Garofalo, A; Marchini, S; Taraboletti, G; Giavazzi, R. Matrix metalloproteinases (MMP9 and MMP2) induce the release of vascular endothelial growth factor (VEGF) by ovarian carcinoma cells: Implications for ascites formation. Cancer Res 2003, 63, 5224–5229. [Google Scholar]
- Kumar, M; Sarin, SK. Is cirrhosis of the liver reversible? Indian J. Pediatr 2007, 74, 393–399. [Google Scholar]
- Park, MJ; Kwak, HJ; Lee, HC; Yoo, DH; Park, IC; Kim, MS; Lee, SH; Rhee, CH; Hong, SI. Nerve growth factor induces endothelial cell invasion and cord formation by promoting matrix metalloproteinase-2 expression through the phosphatidylinositol 3-kinase/Akt signaling pathway and AP-2 transcription factor. J. Biol. Chem 2007, 282, 30485–30496. [Google Scholar]
- Chen, PS; Feng, Z; Zhang, XM; Liu, DF. Expression of HIF-1α and its downstream molecules in rat liver fibrosis tissue. J. Gastroenterol. Hepatol 2006, 21, A144. [Google Scholar]







© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, J.; Fan, R.; Zhao, S.; Liu, L.; Guo, S.; Wu, N.; Zhang, W.; Chen, P. Reactive Oxygen Species Released from Hypoxic Hepatocytes Regulates MMP-2 Expression in Hepatic Stellate Cells. Int. J. Mol. Sci. 2011, 12, 2434-2447. https://doi.org/10.3390/ijms12042434
Li J, Fan R, Zhao S, Liu L, Guo S, Wu N, Zhang W, Chen P. Reactive Oxygen Species Released from Hypoxic Hepatocytes Regulates MMP-2 Expression in Hepatic Stellate Cells. International Journal of Molecular Sciences. 2011; 12(4):2434-2447. https://doi.org/10.3390/ijms12042434
Chicago/Turabian StyleLi, Jing, Renhua Fan, Susu Zhao, Leilei Liu, Shanshan Guo, Nan Wu, Wandong Zhang, and Pingsheng Chen. 2011. "Reactive Oxygen Species Released from Hypoxic Hepatocytes Regulates MMP-2 Expression in Hepatic Stellate Cells" International Journal of Molecular Sciences 12, no. 4: 2434-2447. https://doi.org/10.3390/ijms12042434
APA StyleLi, J., Fan, R., Zhao, S., Liu, L., Guo, S., Wu, N., Zhang, W., & Chen, P. (2011). Reactive Oxygen Species Released from Hypoxic Hepatocytes Regulates MMP-2 Expression in Hepatic Stellate Cells. International Journal of Molecular Sciences, 12(4), 2434-2447. https://doi.org/10.3390/ijms12042434