1. Introduction
The endoplasmic reticulum (ER), a eukaryotic organelle, is the location of post-translational modification. The ER is a major signal transducing organelle that senses and responds to changes in cellular homeostasis. ER stress can be induced by an unfolded protein response (UPR) or by viral infection (ER overload response, EOR), which mediates multiple molecular biological processes via ER stress sensors (inositol-requiring enzyme 1, IRE1, protein kinase-like ER kinase, PERK, and activating transcription factor 6, ATF6), which participate directly or indirectly in the UPR of mammalian cells [
1–
3]. The ER stress response in mammalian cells is triggered by dissociation of BiP from stress transducers, such as PERK, ATF6, and IRE1. BiP binds to ER lumenal un/misfolded proteins and activates the ER stress response [
4]. The first response of ER stress involves up-regulation of genes encoding ER chaperones, which increase protein-folding/assembly activity and prevent protein aggregation in the ER. However, newly synthesized soluble and integral membrane secreted proteins are sometimes retained in the ER due to non-native conformations and are degraded by a process called ER-associated degradation (ERAD) [
5]. Retention of a large number of non-native proteins sometimes induces ER storage disease (ERSD) [
6]. Three functionally different groups of ER molecular chaperones reside in the ER: chaperones of the heat shock protein family, including BiP and its co-chaperone partners, chaperone lectins, such as calnexin and calreticulin, and the foldase family of PDI and ERp29. Radiation is known to induce a series of biochemical events in the cell, but whether radiation directly induces ER stress remains unclear. Recent reports have demonstrated that radiation induces expression of ERp29, a type of ER-resident chaperone, in several types of cultured cells [
7]. UPR is rapidly sensitive to environmental or physical changes. However, incessant ER stressors induce the proapoptotic potential of the UPR, and eventually initiate apoptosis through the ER signal pathway [
8].
Hemolymph is the circulating fluid of insects, similar to mammalian blood, which moves through the open circulatory system, directly bathing the organs and tissues. Compared with mammalian blood, insect hemolymph differs in the absence of erythrocytes and has a high concentration of several types of free amino acids. It serves important roles in the immune system and in transport of hormones, nutrients, and metabolites. Since the early 1970s, proteins of silkworm (
Bombyx mori) hemolymph have been studied as part of the effort to produce more silk; thus, silkworm hemolymph is the most well-understood insect hemolymph. Cultured insect cells have demonstrated greater resistance to radiation-irradiation than those of mammals, and the lepidopteran cell, by far, shows the greatest resistance [
9,
10]; however, the detailed processes underlying this resistance have remained largely unexplored. Recent results have demonstrated that silkworm hemolymph inhibits apoptosis, and associated proteins were isolated [
11–
14].
The PC12 cell line used in this study is widely used as an
in vitro model for the study of neuronal differentiation [
15]. Several studies of PC12 differentiation have demonstrated an association of ionizing radiation with increased expression of cytokines [
16], including IL-6 [
17], through NFxB activation [
18].
We are interested in the effect of silkworm hemolymph on gene expression of ER chaperones under radiation-irradiation in PC12 cells. In particular, we are interested in the expression of the “ischemia-responsive protein 94 kDa” (irp94) gene, which was first isolated from rat hippocampus. irp94, a novel member of the HSP110 family, was suggested to play an important role in transient forebrain ischemia [
19]. We also demonstrated up-regulation of irp94 by ER stressors in PC12 cells [
20,
21]. Eventually, we would like to determine whether radiation-irradiation induces expression of ER chaperones, and how silkworm hemolymph affects gene expression under such conditions. Additionally, we would like to determine whether the molecular behavior of irp94 to both radiation-irradiation and silkworm hemolymph is the same as those of ER chaperones in the PC12 cell. This may provide clues to understanding irp94 molecular features associated with extracellular stressors, including ER stress and radiation.
2. Results and Discussion
In preliminary experiments, the effect of various doses of radiation on BiP expression was assessed in PC12 cells by RT-PCR, and no evidence noting remarkable changes of ER chaperone BiP by
B. mori hemolymph was found, apart from in the cell culture condition (5%) (data not shown here). Increased gene expression of BiP was observed at tested doses (5, 10, 15 Gy) and maximal induction was observed at 10 Gy of irradiation (~2–fold more than controls). Time-course studies indicated that expression of BiP showed an increase at 6 h after radiation at a dose of 10 Gy, with a peak at 24 h, and remained elevated even at 48 h post-irradiation. We also tested whether
B. mori hemolymph directly regulated the expression of UPR molecules, and the dose-dependent effect on gene expression and induction of apoptosis according to previous data [
22,
23]. We also tested cell viability of
B. mori hemolymph without radiation condition by MTT assay that no obvious changes were detected (data not shown), and we then treated healthy cells, which showed no apoptosis over the previous 7 days, with 5% hemolymph.
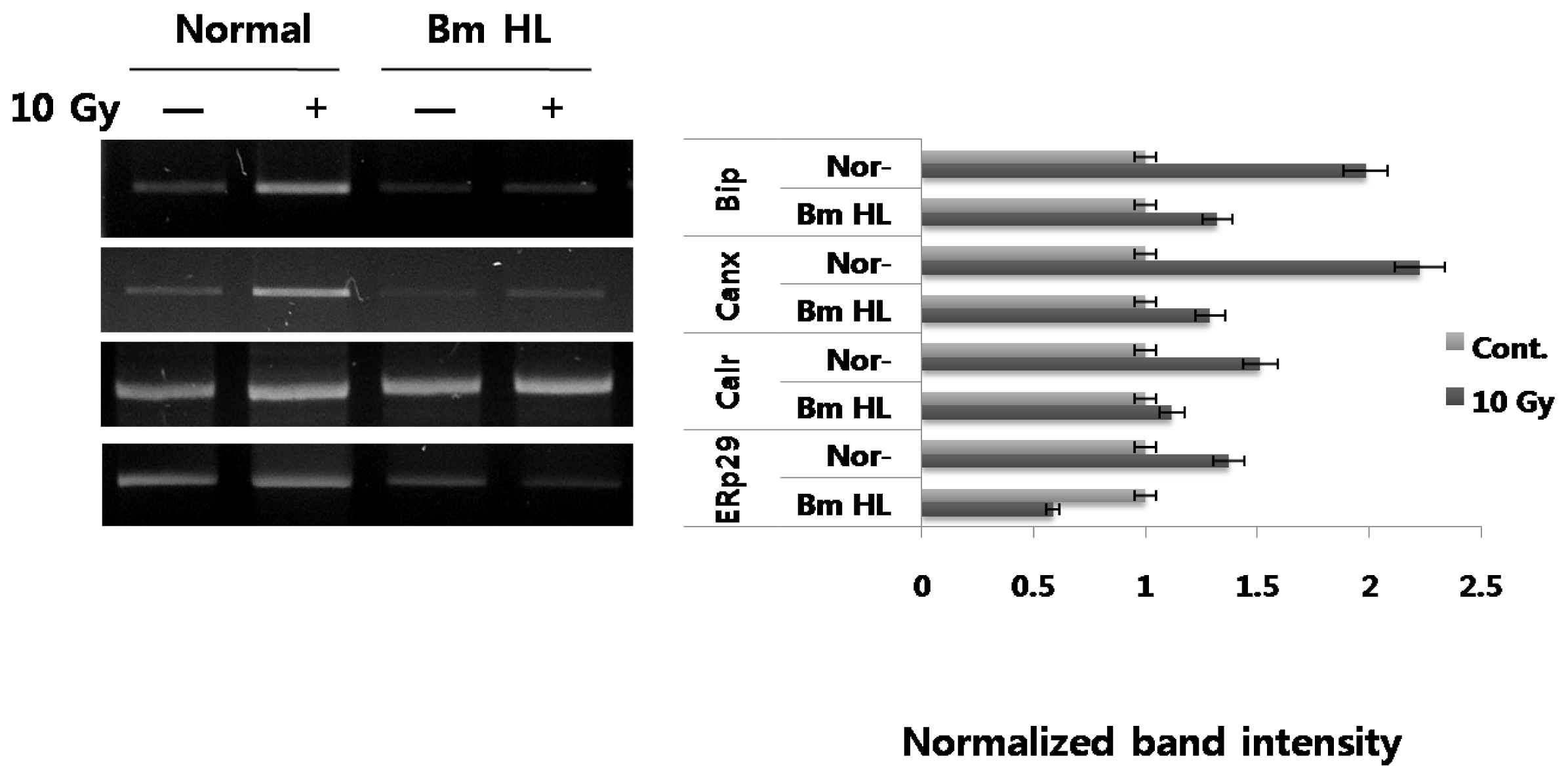
First, we evaluated the expression of ER chaperones induced by radiation in PC12 cells. As shown in
Figure 1, radiation induced up-regulation in all tested ER chaperones (BiP and calnexin (~2-fold), calreticulin and ERp29 (~1.5-fold)) at the mRNA level. Next, we tested the additional effect of silkworm hemolymph on ER chaperone expression under radiation-irradiation conditions. Interestingly, addition of silkworm hemolymph to irradiated cells resulted in an obvious decrease in ER chaperone expression, compared with radiation alone; in the extreme case of ERp29, expression was down-regulated, compared with the control (indicated by BmHL). Need more experiments to understand that pretreatment or posttreatment of silkworm hemolymph which directly changes the basal level of UPR state.
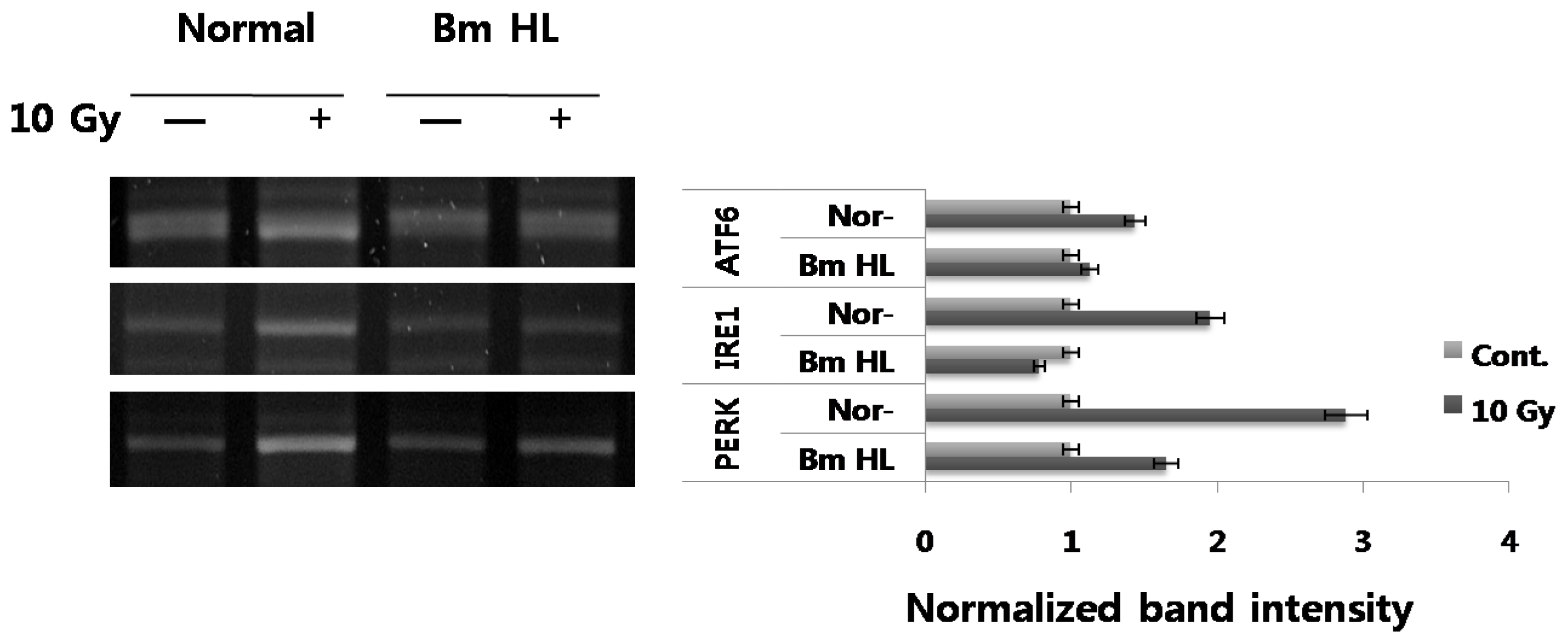
UPR in mammalian cells involves three distinct ER stress sensors (IRE1, PERK, ATF6) that are downstream components of ER chaperones, and which transmit stress signals from the ER to the nucleus in response to perturbations in protein folding in the ER. While activation (autophosphorylation and dimerization) of IRE1 activates the endonuclease domains, which cleave X-box DNA-binding protein (XBP) mRNA, generating an activated form of XBP1, activation of PERK results in phosphorylation of the α subunit of eukaryotic translation initiation factor 2 (eIF2α) and inhibits translation initiation. ATF6 is cleaved at the cytosolic face of the membrane in response to ER stress, causing nuclear translocation of the N-terminal cytoplasmic domain, which contains the DNA-binding, dimerization, and transactivation domains, and subsequent binding to both ER stress-response element (ERSE) and ATF6 sites to enhance expression of ER molecular chaperone genes. We also evaluated the expression of the three ER membrane kinases in response to radiation-irradiation and the addition of silkworm hemolymph (
Figure 2). As shown in
Figure 1, radiation induced up-regulation in the expression of IRE1 (~2-fold), PERK (~3-fold), and ATF6 (~1.5-fold), and with the addition of silkworm hemolymph, expression of these genes showed marked decrease.
B. mori hemolymph, specifically induced the ER stress sensors in the radiation condition as seen by transcriptional levels. However, its mechanism of action is not fully understood and needs more experiments for the XBP1 mRNA splicing, eIF2 alpha expression and ARF6 fragmentation.
Our results (
Figures 1 and
2) indicated that radiation-irradiation induces up-regulation of ER chaperones, including BiP, calnexin, calreticulin, and ERp29, and ER stress sensors, including IRE1, PERK, and ATF6 at the mRNA level in PC12 cells. In particular, increased expression of ER membrane kinases suggested a strong association of stimulation by radiation with expression of ER chaperone genes and the direct ER signal transduction pathway. However, the detailed molecular mechanism underlying UPR induced by radiation will require significant study. Here, suppression of the expression of both ER chaperones and ER stress sensors by the addition of silkworm hemolymph under radiation-irradiation conditions is an interesting finding. Factors in silkworm hemolymph point to its potential for protection against extracellular-disturbance induced by radiation through down-regulation of ER stress associated proteins.
A gene encoding irp94 was first identified following characterization of a novel protein of the HSP110 family that may play an important role against ischemia. We previously demonstrated that expression of irp94 is regulated by ER stressors in neurons and thyrocytes. Although increased expression of ER chaperones (BiP, calnexin, calreticulin, ERp29) by radiation-irradiation was demonstrated in this study (
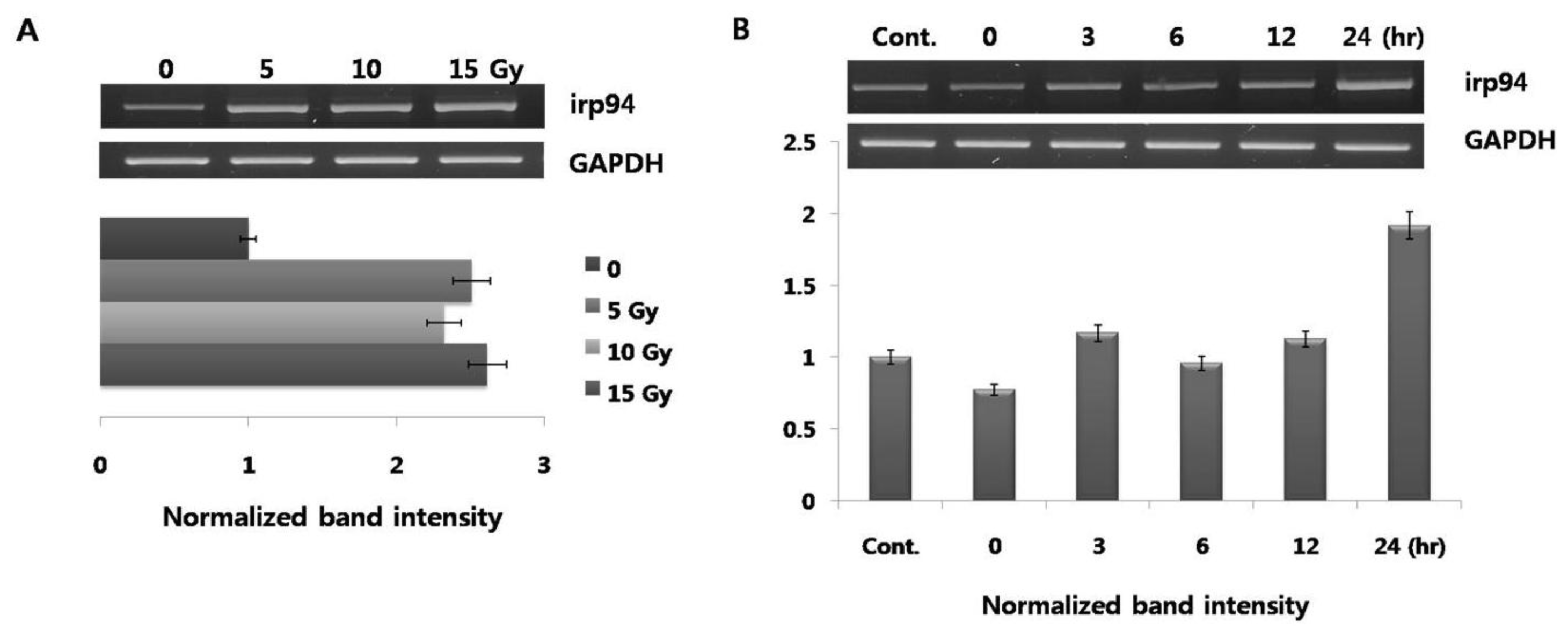
Figure 1), expression of irp94 is unclear. We then performed testing to determine whether expression of irp94 was activated in PC12 cells stimulated by radiation-irradiation. Its expression showed a rapid increase in response to a radiation dose of 5 Gy (~2.5-fold;
Figure 3A), and its expression was almost constant at 15 Gy. The level of increased irp94 expression by radiation is similar to those of ER chaperones (BiP and calnexin in
Figure 1). When cells were irradiated with 10 Gy, the highest expression was detected after a 24 h chase (
Figure 3B). This study is the first to show that radiation induced an increase in the mRNA level of irp94 in PC12 cells.
From the data above (
Figure 1), while ER chaperones were up-regulated by radiation-irradiation, expression of these genes was down-regulated by the addition of silkworm hemolymph. We also performed testing to determine whether irp94 expression was induced by the addition of silkworm hemolymph after irradiation. Under these expression conditions, when cells are exposed to silkworm hemolymph, expression of ER chaperones was significantly down-regulated, as demonstrated by the results above. However, as shown in
Figure 4, in contrast to expectations, irp94 expression was not obviously changed, and was not down-regulated by the addition of silkworm hemolymph after irradiation, unlike the other ER chaperones.
PC12 cells were irradiated at a dose of 10 Gy and treated with or without 5% silkworm hemolymph. Total RNA was harvested at 24 h post irradiation. All mRNA levels of immunoglobulin heavy chain binding protein (BiP), calnexin (Canx), calreticulin (Calr), and endoplasmic reticulum protein 29 (ERp29) were measured by semiquantitative RT-PCR. All experiments were performed at least three times and results represent the average. The uptake ratio (fold) is shown relative to control (1-fold). RT-PCR primers used are shown in the Experimental Section.
All mRNA levels of activating transcription factor 6 (ATF6), inositol-requiring enzyme 1 (IRE1), and protein kinase-like ER kinase (PERK) were measured by semiquantitative RT-PCR. All experimental conditions were the same as those in
Figure 1.
PC12 cells were irradiated at a dose of 10 Gy and total RNA was harvested. All mRNA levels of irp94 were measured by semiquantitative RT-PCR. All experimental conditions were the same as those in
Figure 1.
3. Experimental Section
3.1. Cell Culture and Radiation Exposure
PC12 is a cell line derived from a pheochromocytoma of the rat adrenal medulla, which is useful as a model system for neuronal experiments. PC12 cells were cultured on collagen coated flasks in 85% RPMI 1640 supplemented with 25 mM Hepes buffer, 10% heat-inactivated horse serum, and 5% heat-inactivated fetal bovine serum, 2 mM L-glutamine, 1 mM sodium pyruvate, 1 g/L d-(+)-glucose, and antibiotics: 25 μg/mL streptomycin and 25 U/mL penicillin. Cells were maintained in a humidified incubator at 37 °C in a 5% CO2 atmosphere. The medium was exchanged every 48 h. Cells were rinsed with 1 × PBS, pH 7.0, and detached with 0.25% trypsin/EDTA. Following centrifugation (1000 × g, 5 min), cells were sub-cultured in 25-cm2 flasks using a sub-cultivation ratio of 1:2 to 1:4 and were photographed every 24 h with an inverted microscope. Cells were passaged twice per week. The 80% confluent monolayer of PC12 cells was pretreated with or without 5% silkworm hemolymph for 24 h. The resulting cells were irradiated (X-ray) with a linear accelerator (Clinac 2100 C, Varian, USA) at indicated doses and times. After irradiation, the medium was immediately replaced with fresh PBS. The dishes were then returned to the incubator for another 24 h culture. Total RNA from cultured cells was extracted using an RNA isolation reagent (TRI-Reagent Ambion, Austin, USA) and used for the following RT-PCR experiments.
3.2. Silkworm Hemolymph Collection
Silkworm hemolymph was collected from the fifth-instar larvae of Bombyx mori by clipping the side of an abdominal leg. On average, 0.5 mL of hemolymph was obtained per larva. Collected hemolymph was heat-treated at 60 °C for 30 min, and then chilled and centrifuged (10,000 × g, 30 min). The supernatant was filtered through a 0.2-μ membrane filter and kept at 4 °C; 5% SH was added directly to the medium.
3.3. Semiquantitative RT-PCR
RT-PCR was performed using the forward primer (F) (5′–ACCACCAGTCCATCGCCATT-3′) and reverse primer (R) (5′–CCACCCTGGACGGAAGTTTG-3′) for IRE1, F (5′–AGTGGTGGCCAC TAATGGAG-3′) and R (5′–TCTTTTGTCAGGGGTCGTTC-3′) for BiP, F (5′–CTAGGCCTGGAGG CCAGGTT-3′) and R (5′–ACCCTGGAGTATGCGGGTTT-3′) for ATF6, F (5′–GGTCTGGTTCCT TGGTTTCA-3′) and R (5′–TTCGCTGGCTGTGTAACTTG-3′) for PERK, F (5′–GGGAGTCTTG TCGTGGAATTG-3′) and R (5′–TGCTTTCCAAGACGGCAGA-3′) for calnexin, F (5′–ACATCAG GAGCTAAAAGCAGCC-3′) and R (5′–TGAAACATACGTCACCCGCA-3′) for calreticulin, F (5′–CAGGATTTGCCCTATCCAGA-3′) and R (5′–GTCATTCCGTTCCTTCTCCA-3′) for irp94, F (5′–TAC AAGGTCATTCCCAAAAGCAAGT-3′) and R (5′–CGGAAGAGGTAGAAGACTGGG TAGC-3′) for ERp29, and F (5′–ACATCAAATGGGGTGATGCT-3′) and R (5′-AGGAGACAAC CTGGT CCTCA-3′) for GAPDH. RT-PCR primers were supplied from Bioneer Co. (Taejon, Korea). Unless otherwise noted, all other chemicals were purchased from Sigma (St. Louis, USA). RT-PCR conditions were as follows: 30 cycles (94 °C for 30 s, 58 °C for 30 s, and 72 °C for 1 min (with 10 min for the final cycle)) using the above primers with Taq DNA polymerase.


{kind=link}
{kind=link}
{kind=link}
{kind=link}



