Lateral Distribution of NBD-PC Fluorescent Lipid Analogs in Membranes Probed by Molecular Dynamics-Assisted Analysis of Förster Resonance Energy Transfer (FRET) and Fluorescence Quenching

Abstract
:1. Introduction
2. Results and Discussion
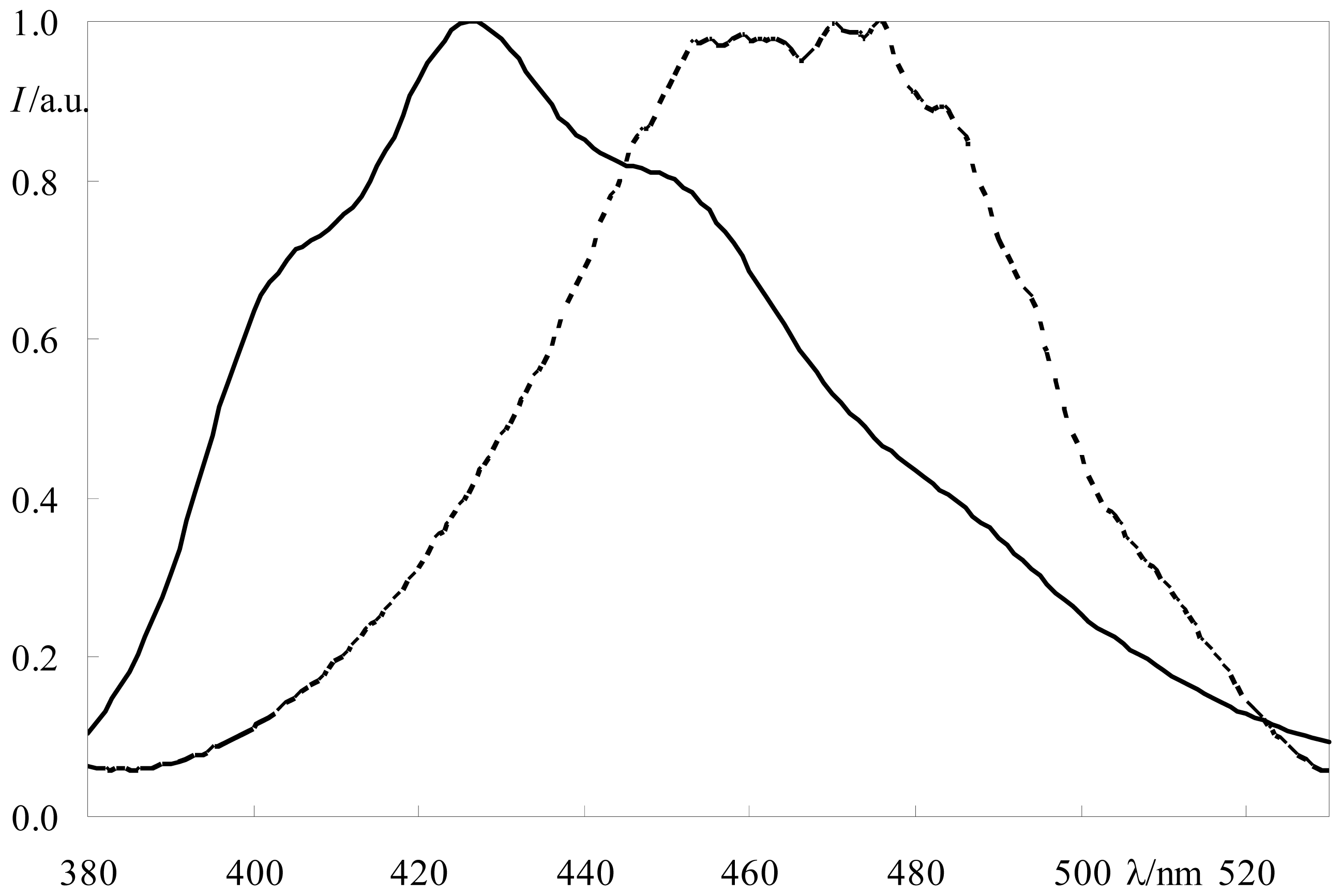
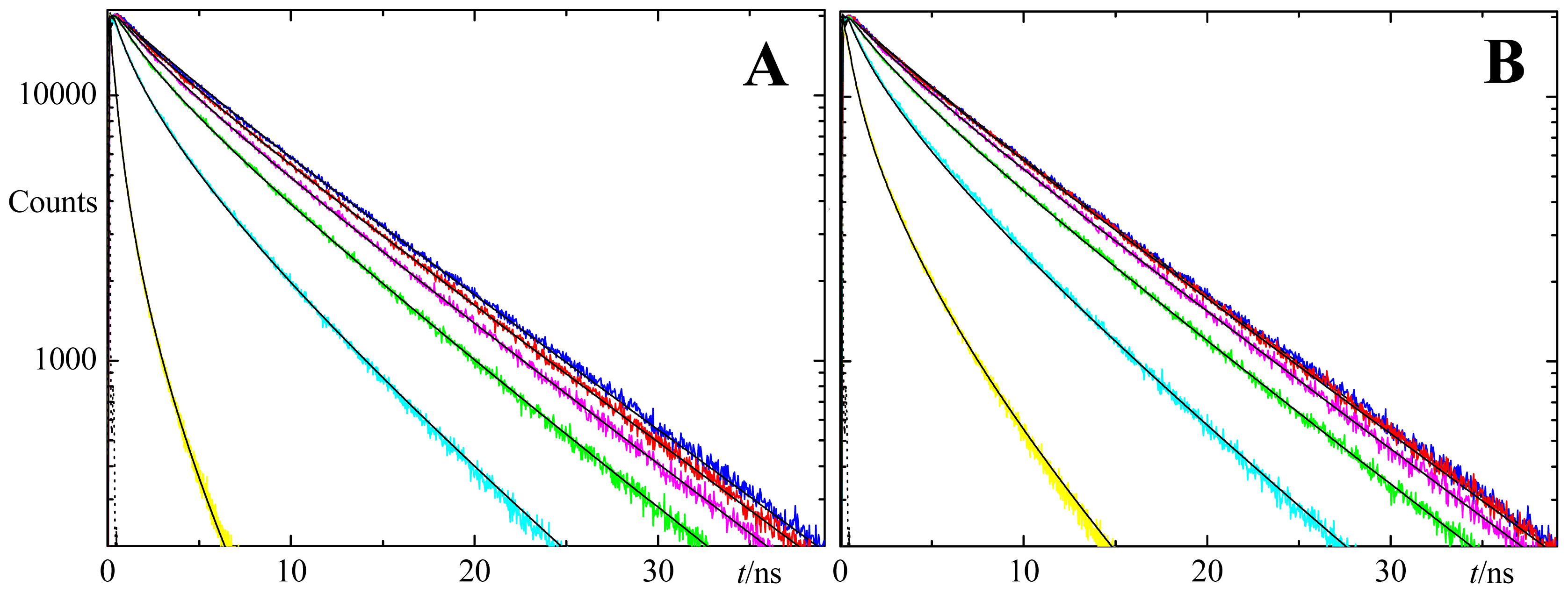
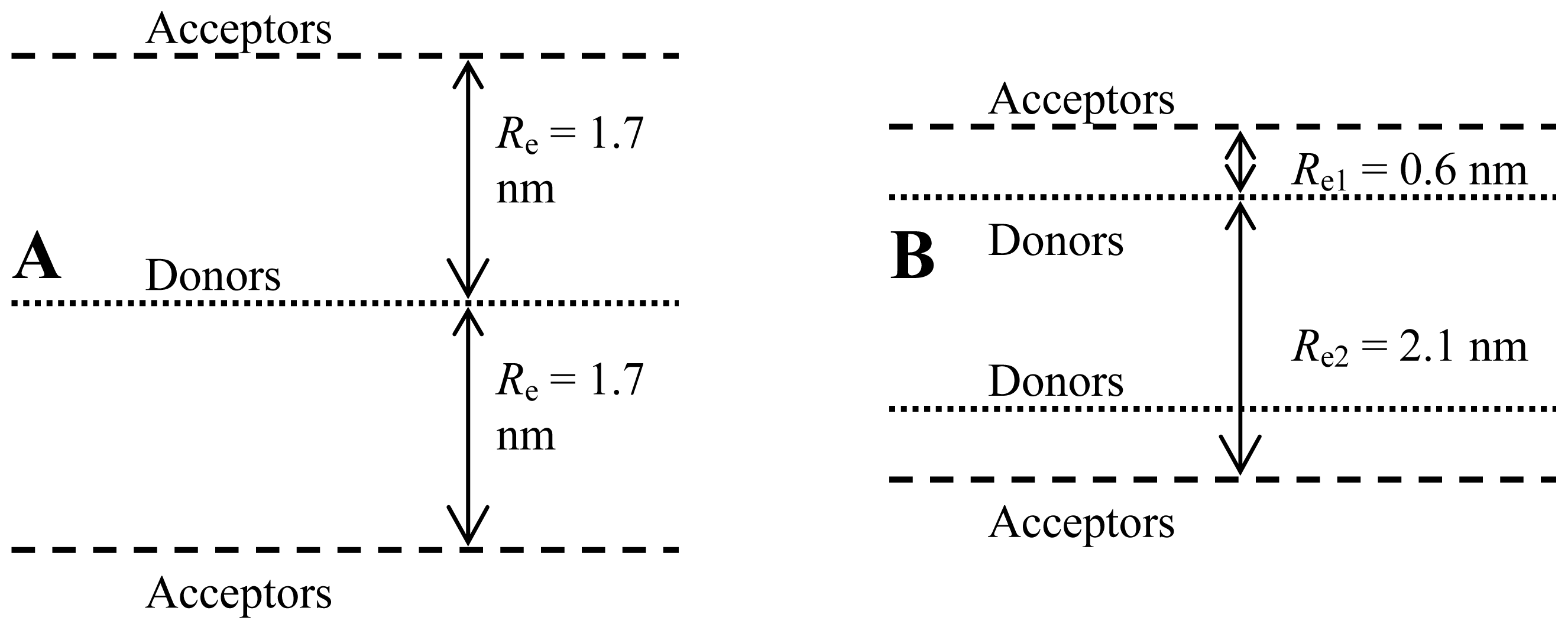
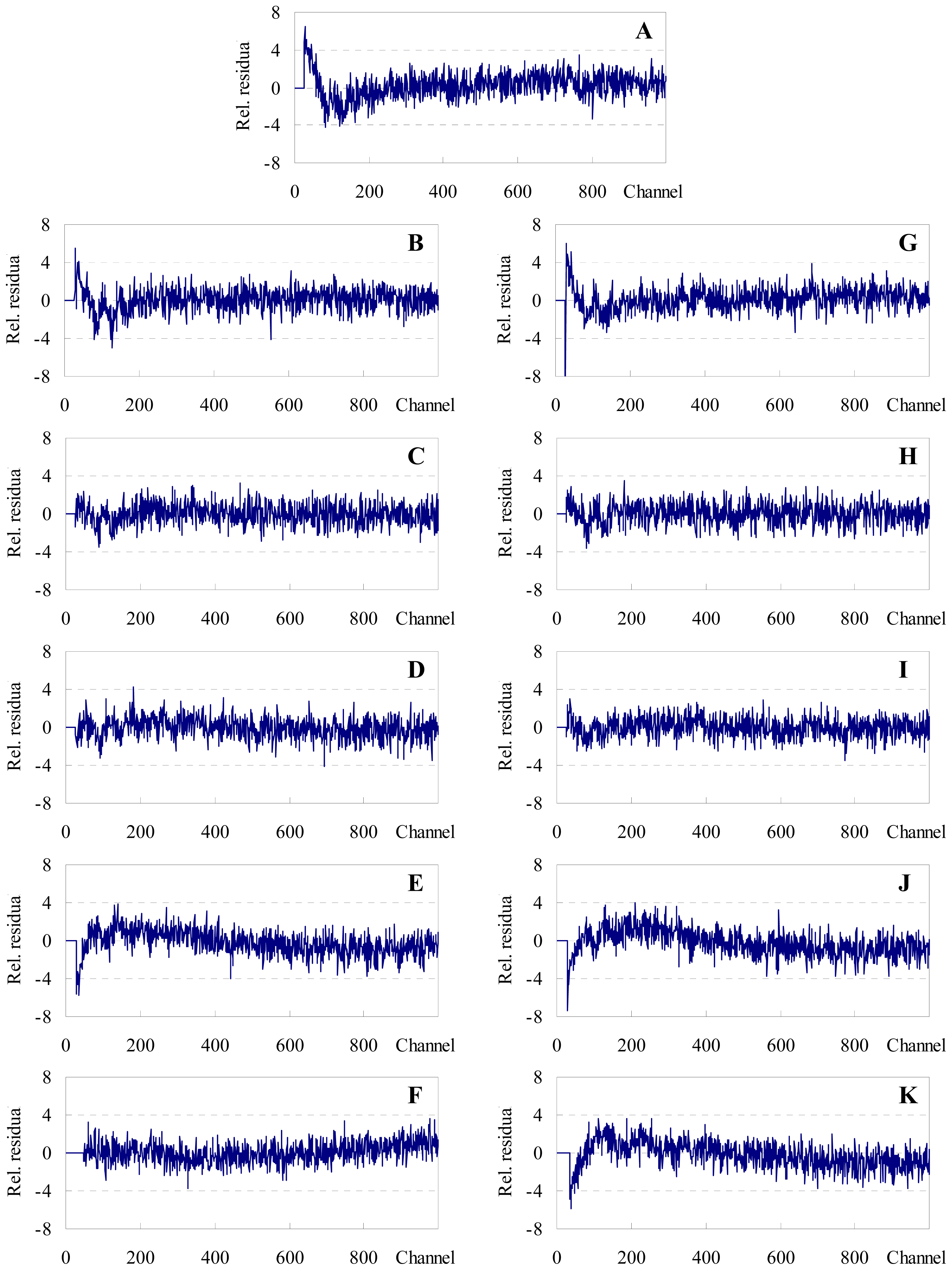
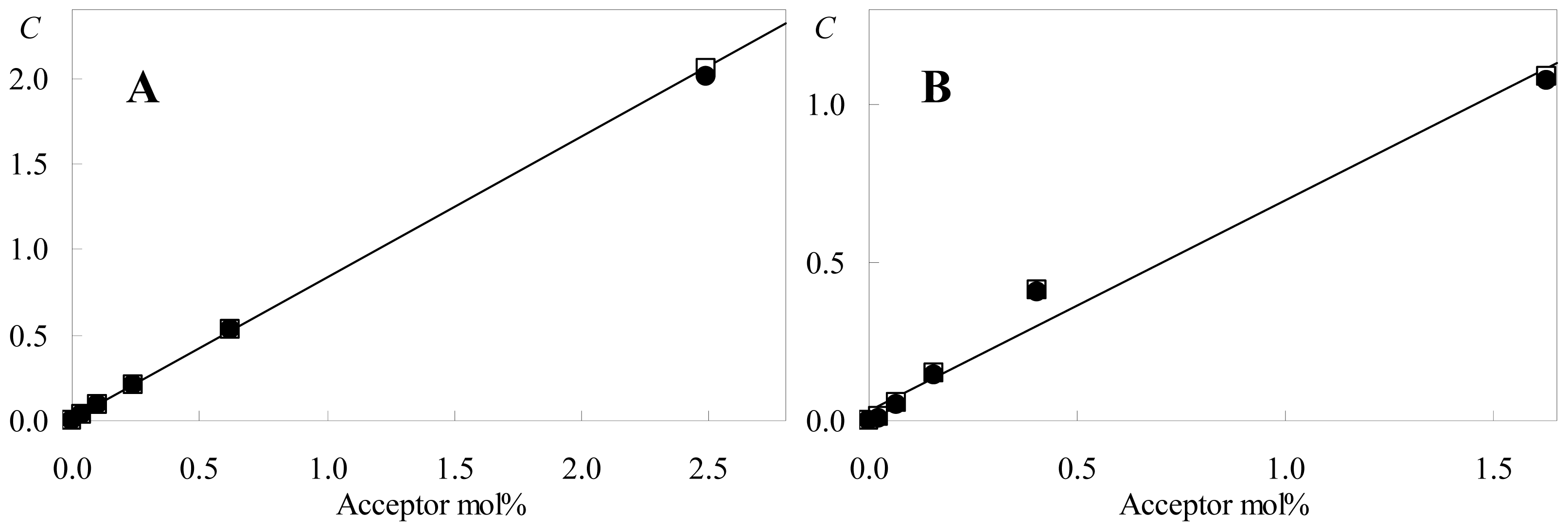
2.1. FRET
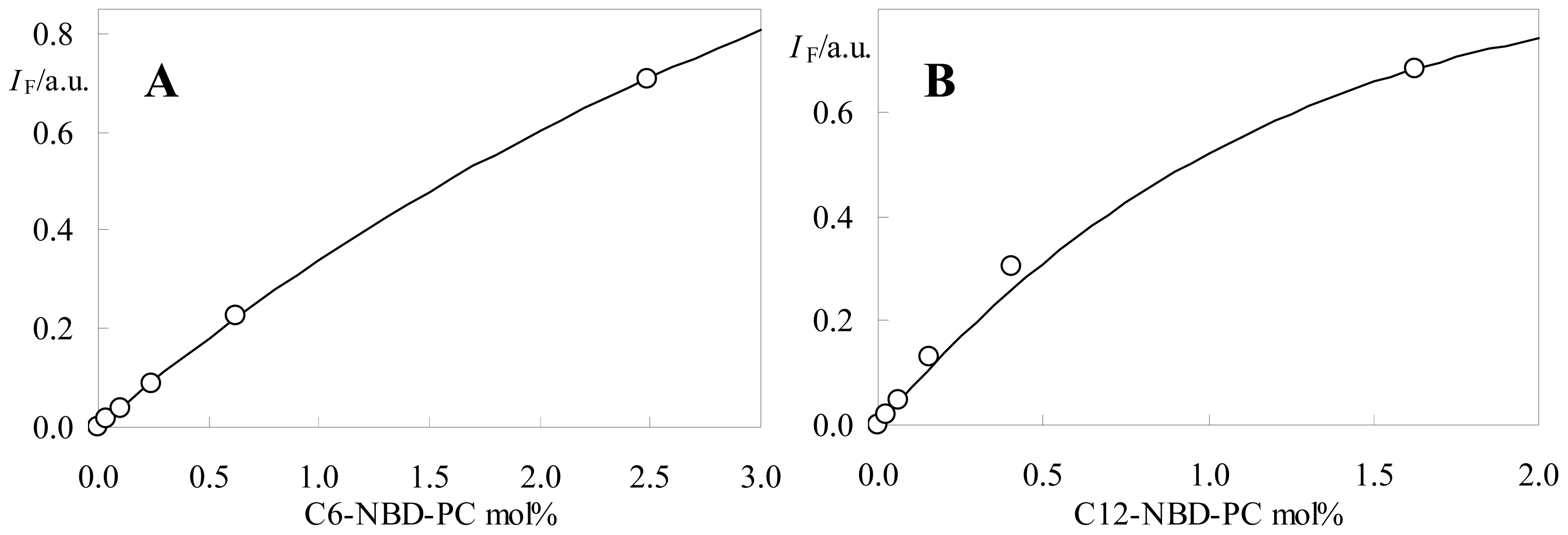
2.2. Fluorescence Self-Quenching
3. Experimental Section
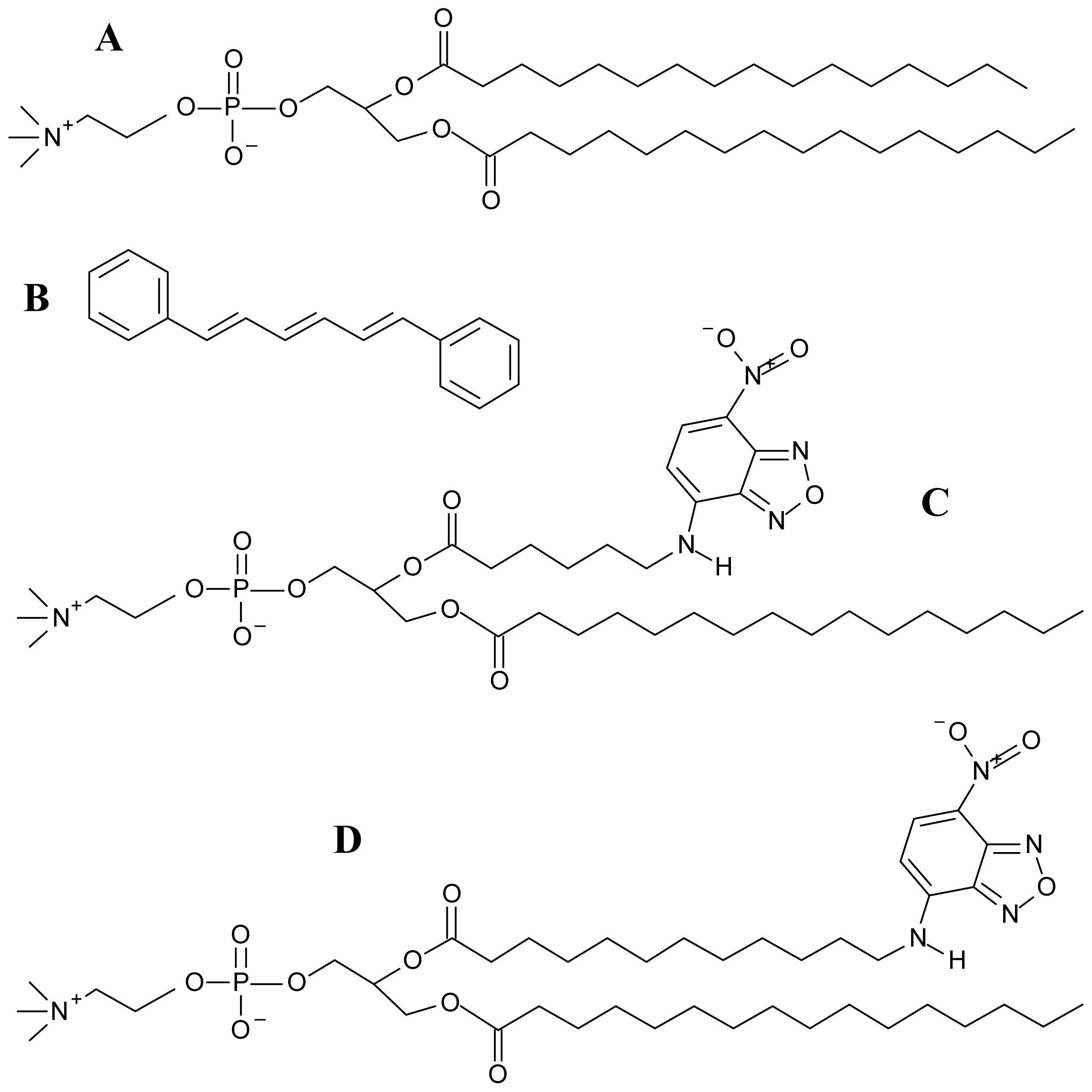
3.1. Materials
3.2. Preparation of Large Unilamellar Vesicles
3.3. Instrumentation
3.4. MD Simulations
4. Conclusions
Acknowledgments
References
- Loura, L.M.S.; Prieto, M. FRET in membrane biophysics: An overview. Front Physiol 2011, 2, 82. [Google Scholar]
- De Almeida, R.F.; Loura, L.M.S.; Prieto, M. Membrane lipid domains and rafts: Current applications of fluorescence lifetime spectroscopy and imaging. Chem. Phys. Lipids 2009, 157, 61–77. [Google Scholar]
- Loura, L.M.S.; de Almeida, R.F.M.; Silva, L.C.; Prieto, M. FRET analysis of domain formation and properties in complex membrane systems. Biochim. Biophys. Acta 2009, 1788, 209–224. [Google Scholar]
- Fung, B.K.; Stryer, L. Surface density determination in membranes by fluorescence energy transfer. Biochemistry 1978, 17, 5241–5248. [Google Scholar]
- Lantzsch, G.; Binder, H.; Heerklotz, H. Surface area per molecule in lipid/C12En membranes as seen by fluorescence resonance energy transfer. J. Fluoresc 1994, 4, 339–343. [Google Scholar]
- Loura, L.M.S.; Fedorov, A.; Prieto, M. Resonance energy transfer in a model system of membranes: Application to gel and liquid crystalline phases. Biophys. J. 1996, 71, 1823–1836. [Google Scholar]
- Loura, L.M.S.; Fedorov, A.; Prieto, M. Membrane probe distribution heterogeneity: A resonance energy transfer study. J. Phys. Chem. B 2000, 104, 6920–6931. [Google Scholar]
- Lentz, B.R. Membrane “fluidity” as detected by diphenylhexatriene probes. Chem. Phys. Lipids 1989, 50, 171–190. [Google Scholar]
- Chattopadhyay, A. Chemistry and biology of N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)-labeled lipids: Fluorescent probes of biological and model membranes. Chem. Phys. Lipids 1990, 53, 1–15. [Google Scholar]
- Repáková, J.; Čapková, P.; Holopainen, J.M.; Vattulainen, I. Distribution, orientation, and dynamics of DPH probes in DPPC bilayer. J. Phys. Chem. B 2004, 108, 13438–13448. [Google Scholar]
- Repáková, J.; Holopainen, J.M.; Morrow, M.R.; McDonald, M.C.; Čapková, P.; Vattulainen, I. Influence of DPH on the structure and dynamics of a DPPC bilayer. Biophys. J 2005, 88, 3398–3410. [Google Scholar]
- Loura, L.M.S.; Prates Ramalho, J.P. Location and dynamics of acyl chain NBD-labeled phosphatidylcholine (NBD-PC) in DPPC bilayers. A molecular dynamics and time-resolved fluorescence anisotropy study. Biochim. Biophys. Acta 2007, 1768, 467–478. [Google Scholar]
- Loura, L.M.S.; Fernandes, F.; Fernandes, A.C.; Prates Ramalho, J.P. Effects of fluorescent probe NBD-PC on the structure, dynamics and phase transition of DPPC. A molecular dynamics and differential scanning calorimetry study. Biochim. Biophys. Acta 2008, 1778, 491–501. [Google Scholar]
- Loura, L.M.S.; Palace Carvalho, A.J.; Prates Ramalho, J.P. Direct calculation of Förster orientation factor of membrane probes by molecular simulation. J. Mol. Struct. THEOCHEM 2010, 946, 107–112. [Google Scholar]
- Loura, L.M.S.; Prates Ramalho, J.P. Fluorescent membrane probes’ behaviour in lipid bilayers: Insights from molecular dynamics simulations. Biophys. Rev 2009, 1, 141–148. [Google Scholar]
- Loura, L.M.S.; Prates Ramalho, J.P. Recent developments in molecular dynamics simulations of fluorescent membrane probes. Molecules 2011, 16, 5437–5452. [Google Scholar]
- Filipe, H.A.; Moreno, M.J.; Loura, L.M.S. Interaction of 7-nitrobenz-2-oxa-1,3-diazol-4-yl-labeled fatty amines with 1-palmitoyl, 2-oleoyl-sn-glycero-3-phosphocholine bilayers: A molecular dynamics study. J. Phys. Chem. B 2011, 115, 10109–10119. [Google Scholar]
- Van Der Meer, B.W.; Coker, G., III; Chen, S.-Y.S. Resonance Energy Transfer: Theory and Data; VCH: New York, NY, USA; p. 1994.
- Loura, L.M.S.; Coutinho, A.; Silva, A.; Fedorov, A.; Prieto, M. Structural effects of a basic peptide on the organization of dipalmitoylphosphatidylcholine/dipalmitoylphosphatidylserine membranes: A fluorescent resonance energy transfer study. J. Phys. Chem. B 2006, 110, 8130–8141. [Google Scholar]
- Davenport, L.; Dale, R.E.; Bisby, R.H.; Cundall, R.B. Transverse location of the fluorescent probe 1,6-diphenyl-1,3,5-hexatriene in model lipid bilayer membrane systems by resonance excitation energy transfer. Biochemistry 1985, 24, 4097–4108. [Google Scholar]
- Kaiser, R.D.; London, E. Location of diphenylhexatriene (DPH) and its derivatives within membranes: Comparison of different fluorescence quenching analyses of membrane depth. Biochemistry 1998, 37, 8180–8190. [Google Scholar]
- Madeira, C.; Loura, L.M.S.; Aires-Barros, M.R.; Fedorov, A.; Prieto, M. Characterization of DNA/lipid complexes by fluorescence resonance energy transfer. Biophys. J 2003, 85, 3106–3119. [Google Scholar]
- Fernandes, F.; Loura, L.M.S.; Fedorov, A.; Prieto, M. Absence of clustering of phosphatidylinositol-(4,5)-bisphosphate in fluid phosphatidylcholine. J. Lipid Res. 2006, 47, 1521–1525. [Google Scholar]
- Franquelim, H.G.; Loura, L.M.S.; Santos, N.C.; Castanho, M.A.R.B. Sifuvirtide screens rigid membrane surfaces. Establishment of a correlation between efficacy and membrane domain selectivity among HIV fusion inhibitor peptides. J. Am. Chem. Soc 2008, 130, 6215–6523. [Google Scholar]
- Abrams, F.S.; London, E. Extension of the parallax analysis of membrane penetration depth to the polar region of model membranes: Use of fluorescence quenching by a spin-label attached to the phospholipid polar headgroup. Biochemistry 1993, 32, 10826–10831. [Google Scholar]
- Huster, D.; Müller, P.; Arnold, K.; Herrmann, A. Dynamics of membrane penetration of the fluorescent 7-nitrobenz-2-oxa-1,3-diazol-4-yl (NBD) group attached to an acyl chain of phosphatidylcholine. Biophys. J 2001, 80, 822–831. [Google Scholar]
- Loura, L.M.S. Simple estimation of FRET orientation factor distribution in membranes. Int. J. Mol. Sci 2012. submitted for publication. [Google Scholar]
- Loura, L.M.S.; Prieto, M. Resonance energy transfer in heterogeneous planar and bilayer systems: Theory and simulation. J. Phys. Chem. B 2000, 104, 6911–6919. [Google Scholar]
- Loura, L.M.S.; Castanho, M.A.R.B.; Fedorov, A.; Prieto, M. A photophysical study of the polyene antibiotic filipin. Self-aggregation and filipin-ergosterol interaction. Biochim. Biophys. Acta 2001, 1510, 125–135. [Google Scholar]
- Lancet, D.; Pecht, I. Spectroscopic and immunochemical studies with nitrobenzoxadiazolealanine, a fluorescent dinitrophenyl analog. Biochemistry 1977, 16, 5150–5157. [Google Scholar]
- Fery-Forgues, S.; Fayet, J.-P.; Lopez, A. Drastic changes in the fluorescence properties of NBD probes with the polarity of the medium—Involvement of a TICT state? J. Photochem. Photobiol. A Chem 1993, 70, 229–243. [Google Scholar]
- Hoekstra, D. Fluorescence method for measuring the kinetics of Ca2+-induced phase separations in phosphatidylserine-containing lipid vesicles. Biochemistry 1982, 21, 1055–1061. [Google Scholar]
- Loura, L.M.S.; Fedorov, A.; Prieto, M. Exclusion of a cholesterol analog from the cholesterol-rich phase in model membranes. Biochim. Biophys. Acta 2001, 1511, 236–243. [Google Scholar]
- Tamai, N.; Nambu, Y.; Tanaka, S.; Goto, M.; Matsuki, H.; Kaneshina, S. Volumetric characterization of ester- and ether-linked lipid bilayers by pressure perturbation calorimetry and densitometry. Colloids Surf. B Biointerfaces 2012, 92, 232–239. [Google Scholar]
- Mazères, S.; Schram, V.; Tocanne, J.F.; Lopez, A. 7-nitrobenz-2-oxa-1,3-diazole-4-yl-labeled phospholipids in lipid membranes: Differences in fluorescence behavior. Biophys. J 1996, 71, 327–335. [Google Scholar]
- Stöckl, M.; Plazzo, A.P.; Korte, T.; Herrmann, A. Detection of lipid domains in model and cell membranes by fluorescence lifetime imaging microscopy of fluorescent lipid analogues. J. Biol. Chem 2008, 283, 30828–30837. [Google Scholar]
- Lee, J.; Lentz, B.R. Outer leaflet-packing defects promote poly(ethylene glycol)-mediated fusion of large unilamellar vesicles. Biochemistry 1997, 36, 421–431. [Google Scholar]
- Umberger, J.Q.; LaMer, V.K. The kinetics of diffusion controlled molecular and ionic reactions in solution as determined by measurements of the quenching of fluorescence. J. Am. Chem. Soc 1945, 67, 1099–1109. [Google Scholar]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed; Springer: New York, NY, USA, 2006. [Google Scholar]
- Haugland, R.P. Handbook of Fluorescent Probes and Research Chemicals, 6th ed; Molecular Probes: Eugene, OR, USA, 1996. [Google Scholar]
- Hope-Roberts, M.; Wainwright, M.; Horobin, R.W. A nitrobenzofuran-conjugated phosphatidylcholine (C12-NBD-PC) as a stain for membrane lamellae for both microscopic imaging and spectrofluorimetry. Biotech. Histochem 2008, 83, 25–28. [Google Scholar]
- McClare, C.W. An accurate and convenient organic phosphorus assay. Anal. Biochem 1971, 39, 527–530. [Google Scholar]
- Coutinho, A.; Prieto, M. Cooperative partition model of nystatin interaction with phospholipid vesicles. Biophys. J 2003, 84, 3061–3078. [Google Scholar]
- Castanho, M.A.R.B.; Santos, N.C.; Loura, L.M.S. Separating the turbidity spectra of vesicles from the absorption spectra of membrane probes and other chromophores. Eur. Biophys. J 1997, 26, 253–259. [Google Scholar]
- Loura, L.M.S.; Fedorov, A.; Prieto, M. Partition of membrane probes in a gel/fluid two-component lipid system: A fluorescence resonance energy transfer study. Biochim. Biophys. Acta 2000, 1467, 101–112. [Google Scholar]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; Hermans, J. Interaction models for water in relation to protein hydration. In Intermolecular Forces; Pullman, B., Ed.; Reidel: Dordrecht, The Netherlands, 1981; pp. 331–342. [Google Scholar]
- Berendsen, H.J.C.; van der Spoel, D.; van Drunen, R. GROMACS: A message-passing parallel molecular dynamics implementation. Comp. Phys. Comm 1995, 91, 43–56. [Google Scholar]
- Lindahl, E.; Hess, B.; van der Spoel, D. GROMACS 3.0: A package for molecular simulation and trajectory analysis. J. Mol. Mod 2001, 7, 306–317. [Google Scholar]
- Berendsen, H.J.C.; Postma, J.P.M.; di Nola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys 1984, 81, 3684–3690. [Google Scholar]
- Miyamoto, S.; Kollman, P.A. SETTLE: An analytical version of the SHAKE and RATTLE algorithms for rigid water models. J. Comp. Chem 1992, 13, 952–962. [Google Scholar]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A linear constraint solver for molecular simulations. J. Comp. Chem 1997, 18, 1463–1472. [Google Scholar]
- Feenstra, K.A.; Hess, B.; Berendsen, H.J.C. Improving efficiency of large time-scale molecular dynamics simulations of hydrogen-rich systems. J. Comput. Chem 1999, 20, 786–798. [Google Scholar]
- Anézo, C.; de Vries, A.H.; Höltje, H.D.; Tieleman, D.P.; Marrink, S.J. Methodological issues in lipid bilayer simulations. J. Phys. Chem. B 2003, 107, 9424–9433. [Google Scholar]
- Essman, U.; Perela, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys 1995, 103, 8577–8592. [Google Scholar]
- Berger, O.; Edholm, O.; Jahnig, F. Molecular dynamics simulations of a fluid bilayer of dipalmitoylphosphatidylcholine at full hydration, constant pressure, and constant temperature. Biophys. J 1997, 72, 2002–2013. [Google Scholar]
- Demchenko, A.P.; Mély, Y.; Duportail, G.; Klymchenko, A.S. Monitoring biophysical properties of lipid membranes by environment-sensitive fluorescent probes. Biophys. J 2009, 96, 3461–3470. [Google Scholar]
- London, E.; Ladokhin, A.S. Measuring the depth of amino-acid residues in membrane-inserted peptides by fluorescence quenching. In Peptide Lipid Interactions: Current Topics in Membranes; Simon, S.A., McIntosh, T.J., Eds.; Academic: San Diego, CA, USA, 2002; Volume 52, Chapter 4, pp. 89–115. [Google Scholar]
- Fernandes, M.X.; de la Torre, J.G.; Castanho, M.A.R.B. Joint determination by Brownian dynamics and fluorescence quenching of the in-depth location profile of biomolecules in membranes. Anal. Biochem 2002, 307, 1–12. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model I | Model II | ||||
|---|---|---|---|---|---|
| mol% acceptor | C | χ2 | C | χ2 | |
| No acceptor | 0 | 0 (fixed) | 1.97 | 0 (fixed) | 1.92 |
| C6-NBD-PC acceptor | 0.040 | 0.032 | 1.45 | 0.034 | 1.43 |
| 0.099 | 0.091 | 1.18 | 0.093 | 1.17 | |
| 0.24 | 0.207 | 1.23 | 0.210 | 1.21 | |
| 0.62 | 0.525 | 1.93 | 0.530 | 1.75 | |
| 2.5 | 2.011 | 1.89 | 2.054 | 1.19 | |
| C12-NBD-PC acceptor | 0.026 | 0.0093 | 1.67 | 0.011 | 1.65 |
| 0.065 | 0.053 | 1.24 | 0.055 | 1.23 | |
| 0.16 | 0.146 | 1.09 | 0.149 | 1.08 | |
| 0.41 | 0.407 | 2.06 | 0.411 | 1.95 | |
| 1.6 | 1.076 | 2.47 | 1.085 | 1.94 | |
| Global χ2 | 1.64 | 1.49 | |||
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Loura, L.M.S. Lateral Distribution of NBD-PC Fluorescent Lipid Analogs in Membranes Probed by Molecular Dynamics-Assisted Analysis of Förster Resonance Energy Transfer (FRET) and Fluorescence Quenching. Int. J. Mol. Sci. 2012, 13, 14545-14564. https://doi.org/10.3390/ijms131114545
Loura LMS. Lateral Distribution of NBD-PC Fluorescent Lipid Analogs in Membranes Probed by Molecular Dynamics-Assisted Analysis of Förster Resonance Energy Transfer (FRET) and Fluorescence Quenching. International Journal of Molecular Sciences. 2012; 13(11):14545-14564. https://doi.org/10.3390/ijms131114545
Chicago/Turabian StyleLoura, Luís M. S. 2012. "Lateral Distribution of NBD-PC Fluorescent Lipid Analogs in Membranes Probed by Molecular Dynamics-Assisted Analysis of Förster Resonance Energy Transfer (FRET) and Fluorescence Quenching" International Journal of Molecular Sciences 13, no. 11: 14545-14564. https://doi.org/10.3390/ijms131114545
APA StyleLoura, L. M. S. (2012). Lateral Distribution of NBD-PC Fluorescent Lipid Analogs in Membranes Probed by Molecular Dynamics-Assisted Analysis of Förster Resonance Energy Transfer (FRET) and Fluorescence Quenching. International Journal of Molecular Sciences, 13(11), 14545-14564. https://doi.org/10.3390/ijms131114545