Lysophosphatidylcholine Acyltransferase 3 Is the Key Enzyme for Incorporating Arachidonic Acid into Glycerophospholipids during Adipocyte Differentiation


 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Lysophospholipid Acyltransferase Activities Were Increased during Adipogenesis
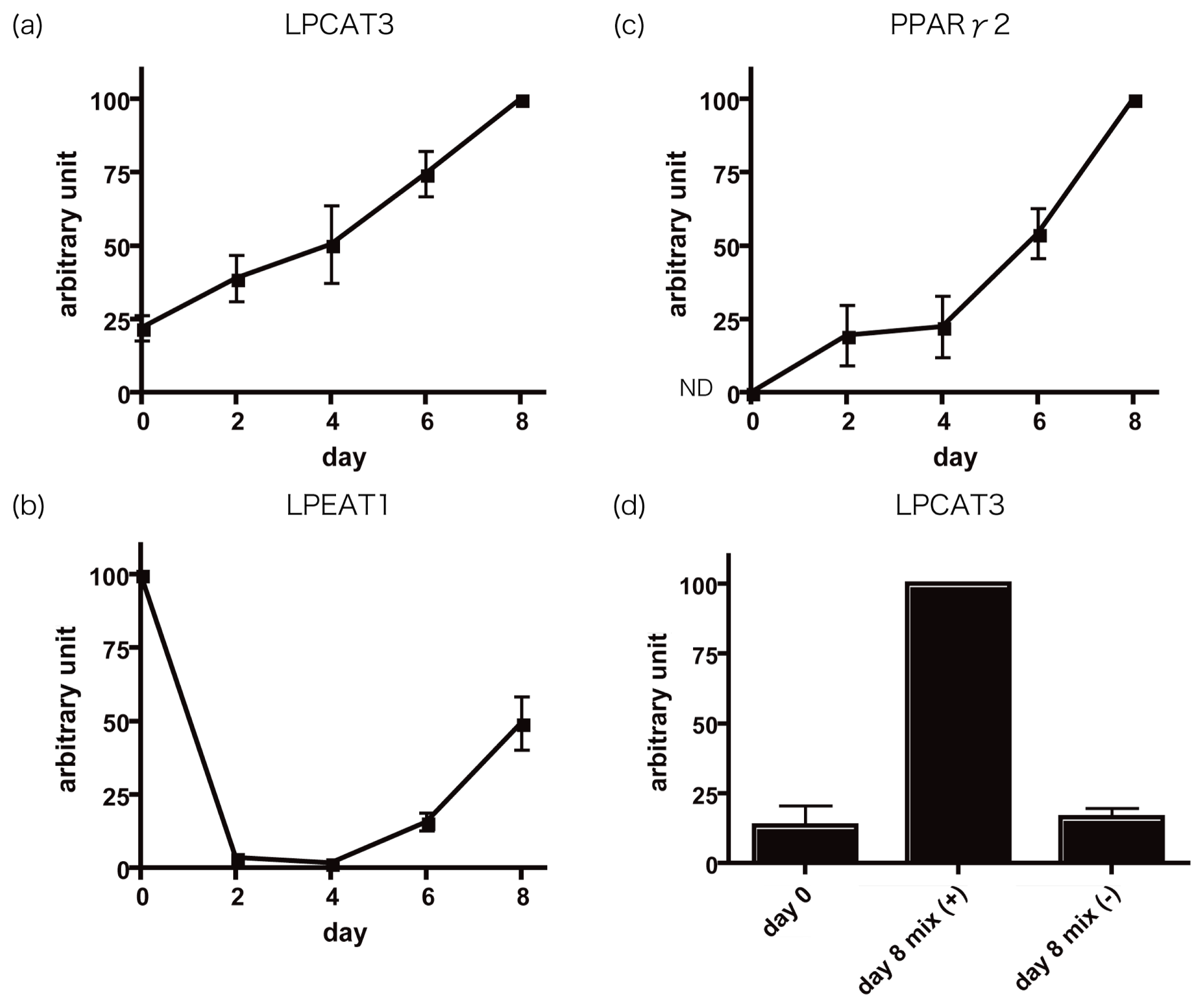
2.2. LPCAT3 mRNA Was Increased during Adipocyte Differentiation
2.3. Change in Phospholipid Composition during Adipocyte Differentiation
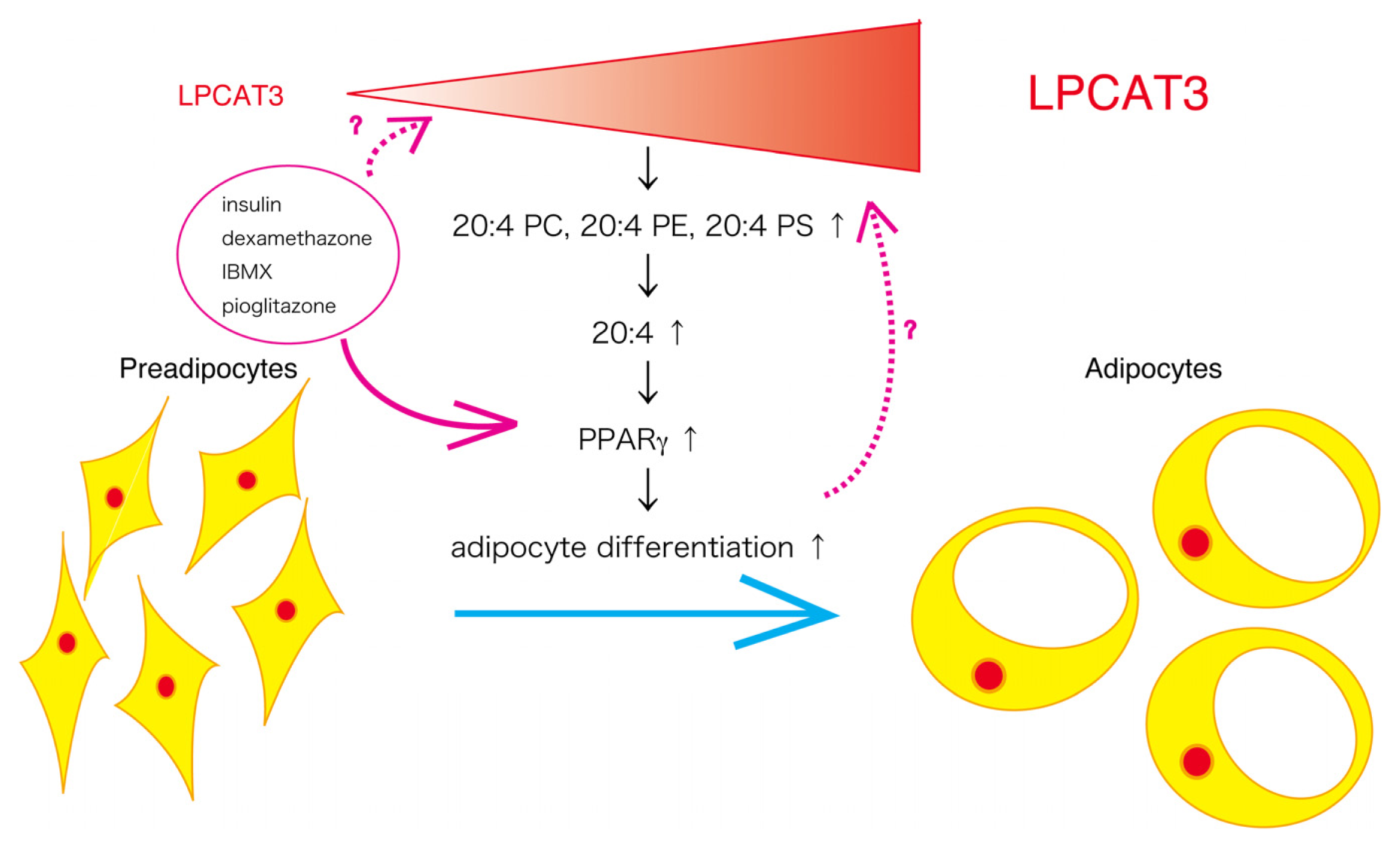
2.4. Possible Role of LPCAT3 in Adipocyte Differentiation
3. Experimental Section
3.1. Materials
3.2. Cell Culture
3.3. Quantitative RT-PCR Analysis
3.4. Microsomal Protein Preparation and Lipid Extraction
3.5. LPLAT Assays
3.6. Reversed Phase Liquid Chromatography
3.7. Mass Spectrometry
3.8. Statistics
4. Conclusions
Acknowledgments
References
- MacDonald, J.I.; Sprecher, H. Phospholipid fatty acid remodeling in mammalian cells. Biochim. Biophys. Acta 1991, 1084, 105–121. [Google Scholar]
- Wood, R.; Harlow, R.D. Structural analyses of rat liver phosphoglycerides. Arch. Biochem. Biophys 1969, 135, 272–281. [Google Scholar]
- Schlame, M.; Brody, S.; Hostetler, K.Y. Mitochondrial cardiolipin in diverse eukaryotes. Comparison of biosynthetic reactions and molecular acyl species. Eur. J. Biochem 1993, 212, 727–735. [Google Scholar]
- Yamashita, A.; Sugiura, T.; Waku, K. Acyltransferases and transacylases involved in fatty acid remodeling of phospholipids and metabolism of bioactive lipids in mammalian cells. J. Biochem 1997, 122, 1–16. [Google Scholar]
- Kennedy, E.P.; Weiss, S.B. The function of cytidine coenzymes in the biosynthesis of phospholipides. J. Biol. Chem 1956, 222, 193–214. [Google Scholar]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol 2008, 9, 112–124. [Google Scholar]
- Coleman, R.A.; Lee, D.P. Enzymes of triacylglycerol synthesis and their regulation. Prog. Lipid Res 2004, 43, 134–176. [Google Scholar]
- Lands, W.E. Stories about acyl chains. Biochim. Biophys. Acta 2000, 1483, 1–14. [Google Scholar]
- Waku, K.; Nakazawa, Y. Acyltransferase activity to 1-acyl-, 1-O-alkenyl-, and 1-O-alkyl-glycero-3-phosphorylcholine in ehrlich ascites tumor cells. J. Biochem 1972, 72, 495–497. [Google Scholar]
- Shimizu, T. Lipid mediators in health and disease: Enzymes and receptors as therapeutic targets for the regulation of immunity and inflammation. Annu. Rev. Pharmacol. Toxicol 2009, 49, 123–150. [Google Scholar]
- Lands, W.E. Metabolism of glycerolipides: A comparison of lecithin and triglyceride synthesis. J. Biol. Chem 1958, 231, 883–888. [Google Scholar]
- Shindou, H.; Hishikawa, D.; Harayama, T.; Yuki, K.; Shimizu, T. Recent progress on acyl CoA: Lysophospholipid acyltransferase research. J. Lipid Res 2009, 50, S46–S51. [Google Scholar]
- Shindou, H.; Shimizu, T. Acyl-CoA:Lysophospholipid acyltransferases. J. Biol. Chem 2009, 284, 1–5. [Google Scholar]
- Lewin, T.M.; Wang, P.; Coleman, R.A. Analysis of amino acid motifs diagnostic for the sn-glycerol-3-phosphate acyltransferase reaction. Biochemistry 1999, 38, 5764–5771. [Google Scholar]
- Yamashita, A.; Nakanishi, H.; Suzuki, H.; Kamata, R.; Tanaka, K.; Waku, K.; Sugiura, T. Topology of acyltransferase motifs and substrate specificity and accessibility in 1-acyl-sn-glycero-3-phosphate acyltransferase 1. Biochim. Biophys. Acta 2007, 1771, 1202–1215. [Google Scholar]
- Harayama, T.; Shindou, H.; Ogasawara, R.; Suwabe, A.; Shimizu, T. Identification of a novel noninflammatory biosynthetic pathway of platelet-activating factor. J. Biol. Chem 2008, 283, 11097–11106. [Google Scholar]
- Shindou, H.; Eto, M.; Morimoto, R.; Shimizu, T. Identification of membrane O-acyltransferase family motifs. Biochem. Biophys. Res. Commun 2009, 383, 320–325. [Google Scholar]
- Hishikawa, D.; Shindou, H.; Kobayashi, S.; Nakanishi, H.; Taguchi, R.; Shimizu, T. Discovery of a lysophospholipid acyltransferase family essential for membrane asymmetry and diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 2830–2835. [Google Scholar]
- Zhao, Y.; Chen, Y.Q.; Bonacci, T.M.; Bredt, D.S.; Li, S.; Bensch, W.R.; Moller, D.E.; Kowala, M.; Konrad, R.J.; Cao, G. Identification and characterization of a major liver lysophosphatidylcholine acyltransferase. J. Biol. Chem 2008, 283, 8258–8265. [Google Scholar]
- Gijon, M.A.; Riekhof, W.R.; Zarini, S.; Murphy, R.C.; Voelker, D.R. Lysophospholipid acyltransferases and arachidonate recycling in human neutrophils. J. Biol. Chem 2008, 283, 30235–30245. [Google Scholar]
- Matsuda, S.; Inoue, T.; Lee, H.C.; Kono, N.; Tanaka, F.; Gengyo-Ando, K.; Mitani, S.; Arai, H. Member of the membrane-bound O-acyltransferase (MBOAT) family encodes a lysophospholipid acyltransferase with broad substrate specificity. Genes Cells 2008, 13, 879–888. [Google Scholar]
- Kadowaki, T.; Yamauchi, T.; Kubota, N.; Hara, K.; Ueki, K.; Tobe, K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J. Clin. Invest 2006, 116, 1784–1792. [Google Scholar]
- Gale, S.E.; Frolov, A.; Han, X.; Bickel, P.E.; Cao, L.; Bowcock, A.; Schaffer, J.E.; Ory, D.S. A regulatory role for 1-acylglycerol-3-phosphate-O-acyltransferase 2 in adipocyte differentiation. J. Biol. Chem 2006, 281, 11082–11089. [Google Scholar]
- Cortes, V.A.; Curtis, D.E.; Sukumaran, S.; Shao, X.; Parameswara, V.; Rashid, S.; Smith, A.R.; Ren, J.; Esser, V.; Hammer, R.E.; et al. Molecular mechanisms of hepatic steatosis and insulin resistance in the AGPAT2-deficient mouse model of congenital generalized lipodystrophy. Cell Metab 2009, 9, 165–176. [Google Scholar]
- Agarwal, A.K.; Arioglu, E.; De Almeida, S.; Akkoc, N.; Taylor, S.I.; Bowcock, A.M.; Barnes, R.I.; Garg, A. AGPAT2 is mutated in congenital generalized lipodystrophy linked to chromosome 9q34. Nat. Genet 2002, 31, 21–23. [Google Scholar]
- Krahmer, N.; Guo, Y.; Wilfling, F.; Hilger, M.; Lingrell, S.; Heger, K.; Newman, H.W.; Schmidt-Supprian, M.; Vance, D.E.; Mann, M.; et al. Phosphatidylcholine synthesis for lipid droplet expansion is mediated by localized activation of CTP:Phosphocholine cytidylyltransferase. Cell Metab 2011, 14, 504–515. [Google Scholar]
- Horl, G.; Wagner, A.; Cole, L.K.; Malli, R.; Reicher, H.; Kotzbeck, P.; Kofeler, H.; Hofler, G.; Frank, S.; Bogner-Strauss, J.G.; et al. Sequential synthesis and methylation of phosphatidylethanolamine promote lipid droplet biosynthesis and stability in tissue culture and in vivo. J. Biol. Chem 2011, 286, 17338–17350. [Google Scholar]
- Reznikoff, C.A.; Brankow, D.W.; Heidelberger, C. Establishment and characterization of a cloned line of C3H mouse embryo cells sensitive to postconfluence inhibition of division. Cancer Res 1973, 33, 3231–3238. [Google Scholar]
- Feldman, B.J.; Streeper, R.S.; Farese, R.V., Jr; Yamamoto, K.R. Myostatin modulates adipogenesis to generate adipocytes with favorable metabolic effects. Proc. Natl. Acad. Sci. USA 2006, 103, 15675–15680. [Google Scholar]
- Ntambi, J.M.; Buhrow, S.A.; Kaestner, K.H.; Christy, R.J.; Sibley, E.; Kelly, T.J., Jr; Lane, M.D. Differentiation-induced gene expression in 3T3-L1 preadipocytes. Characterization of a differentially expressed gene encoding stearoyl-CoA desaturase. J. Biol. Chem. 1988, 263, 17291–17300. [Google Scholar]
- Harmon, G.S.; Lam, M.T.; Glass, C.K. PPARs and lipid ligands in inflammation and metabolism. Chem. Rev 2011, 111, 6321–6340. [Google Scholar]
- Massiera, F.; Saint-Marc, P.; Seydoux, J.; Murata, T.; Kobayashi, T.; Narumiya, S.; Guesnet, P.; Amri, E.Z.; Negrel, R.; Ailhaud, G. Arachidonic acid and prostacyclin signaling promote adipose tissue development: A human health concern? J. Lipid Res 2003, 44, 271–279. [Google Scholar]
- Savva, S.C.; Chadjigeorgiou, C.; Hatzis, C.; Kyriakakis, M.; Tsimbinos, G.; Tornaritis, M.; Kafatos, A. Association of adipose tissue arachidonic acid content with BMI and overweight status in children from Cyprus and Crete. Br. J. Nutr 2004, 91, 643–649. [Google Scholar]
- Williams, E.S.; Baylin, A.; Campos, H. Adipose tissue arachidonic acid and the metabolic syndrome in Costa Rican adults. Clin. Nutr 2007, 26, 474–482. [Google Scholar]
- Koeberle, A.; Shindou, H.; Harayama, T.; Yuki, K.; Shimizu, T. Polyunsaturated fatty acids are incorporated into maturating male mouse germ cells by lysophosphatidic acid acyltransferase 3. FASEB J 2011, 26, 169–180. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol 1959, 37, 911–917. [Google Scholar]



{kind=link}
{kind=link}
| Substrate | Preadipocyte (relative units) | Adipocyte (relative units) | p value |
|---|---|---|---|
| LPCAT activity | |||
| 16:0-CoA | 7.50 ± 0.29 | 12.36 ± 0.68 | p = 0.0008 |
| 18:1-CoA | 3.13 ± 0.19 | 18.24 ± 0.61 | p < 0.0001 |
| 18:2-CoA | 198.50 ± 13.60 | 1821.54 ± 80.62 | p < 0.0001 |
| 20:4-CoA | 267.49 ± 11.45 | 1882 ± 63.12 | p < 0.0001 |
| 22:6-CoA | 0.56 ± 0.04 | 2.30 ± 0.18 | p = 0.0002 |
| LPEAT activity | |||
| 16:0-CoA | 0.20 ± 0.02 | 0.82 ± 0.06 | p = 0.0001 |
| 18:1-CoA | 0.09 ± 0.01 | 3.10 ±0.17 | p < 0.0001 |
| 18:2-CoA | 1.36 ± 0.04 | 14.14 ± 0.63 | p < 0.0001 |
| 20:4-CoA | 2.44 ± 0.12 | 14.44 ± 0.40 | p < 0.0001 |
| 22:6-CoA | 0.02 ± 0.01 | 0.20 ± 0.02 | p = 0.0002 |
| LPSAT activity | |||
| 16:0-CoA | 0.61 ± 0.07 | 1.73 ± 0.05 | p < 0.0001 |
| 18:1-CoA | 0.35 ± 0.03 | 1.93 ± 0.03 | p < 0.0001 |
| 18:2-CoA | 2.65 ± 0.08 | 17.25 ± 0.24 | p < 0.0001 |
| 20:4-CoA | 6.97 ± 0.07 | 28.97 ± 0.88 | p < 0.0001 |
| 22:6-CoA | 0.043 ± 0.002 | 0.151 ± 0.007 | p < 0.0001 |
| PC species | Preadipocyte (%) | Adipocyte (%) | p value |
|---|---|---|---|
| PC | |||
| 30:0 PC | 3.13 ± 0.39 | 2.06 ± 0.17 | ns |
| 30:1 PC | 3.67 ± 0.21 | 2.45 ± 0.03 | 0.0086** |
| 32:0 PC | 10.55 ± 0.71 | 5.32 ± 0.56 | 0.0092** |
| 32:1 PC | 9.61 ± 0.28 | 18.83 ± 0.75 | 0.0007*** |
| 32:2 PC | 1.21 ± 0.17 | 2.95 ± 0.16 | 0.0037** |
| 34:0 PC | 1.98 ± 0.11 | 1.06 ± 0.05 | 0.0031** |
| 34:1 PC | 25.37 ± 1.37 | 22.74 ± 1.29 | ns |
| 34:2 PC | 6.49 ± 0.36 | 7.7 ± 0.66 | ns |
| 34:3 PC | 0.67 ± 0.08 | 1.13 ± 0.03 | 0.0114* |
| 36:0 PC | 0.18 ± 0.02 | 0.12 ± 0.01 | ns |
| 36:1 PC | 6.54 ± 0.56 | 7.32 ± 0.47 | ns |
| 36:2 PC | 12.34 ± 0.88 | 7.75 ± 0.17 | 0.0138* |
| 36:3 PC | 2.81 ± 0.12 | 3.51 ± 0.16 | 0.0452* |
| 36:4 PC | 2.09 ± 0.26 | 4.02 ± 0.31 | 0.0178* |
| 36:5 PC | 0.25 ± 0.04 | 0.73 ± 0.08 | 0.0144* |
| 38:1 PC | 1.78 ± 0.03 | 0.99 ± 0.03 | p < 0.0001*** |
| 38:2 PC | 2.32 ± 0.08 | 1.71 ± 0.12 | 0.0298* |
| 38:3 PC | 1.12 ± 0.06 | 1.22 ± 0.12 | ns |
| 38:4 PC | 3.08 ± 0.17 | 4.36 ± 0.3 | 0.0395* |
| 38:5 PC | 1.75 ± 0.22 | 2.01 ± 0.24 | ns |
| 38:6 PC | 0.35 ± 0.04 | 0.37 ± 0.03 | ns |
| 38:7 PC | 0.04 ± 0.01 | 0.05 ± 0.01 | ns |
| 40:1 PC | 0.22 ± 0.07 | 0.11 ± 0.01 | ns |
| 40:2 PC | 0.27 ± 0.02 | 0.08 ± 0.01 | 0.0044** |
| 40:3 PC | 0.19 ± 0.02 | 0.11 ± 0.01 | ns |
| 40:4 PC | 0.49 ± 0.05 | 0.34 ± 0.04 | ns |
| 40:5 PC | 0.72 ± 0.06 | 0.46 ± 0.06 | ns |
| 40:6 PC | 0.52 ± 0.05 | 0.35 ± 0.03 | ns |
| 40:7 PC | 0.24 ± 0.02 | 0.11 ± 0.02 | 0.0102* |
| PE | |||
| 30:0 PE | 0.1 ± 0.01 | 0.05 ± 0.01 | 0.0237* |
| 32:0 PE | 0.31 ± 0.04 | 0.24 ± 0.01 | ns |
| 32:1 PE | 1.19 ± 0.12 | 5.74 ± 0.06 | p < 0.0001*** |
| 32:2 PE | 0.18 ± 0.02 | 2.66 ± 0.16 | 0.0002*** |
| 34:0 PE | 0.32 ± 0.01 | 0.26 ± 0.01 | 0.0117* |
| 34:1 PE | 7.81 ± 0.25 | 9.71 ± 0.49 | 0.0487* |
| 34:2 PE | 2.95 ± 0.30 | 6.00 ± 0.23 | 0.0027** |
| 34:3 PE | 0.20 ± 0.02 | 0.93 ± 0.07 | 0.001*** |
| 36:1 PE | 14.01 ± 0.06 | 7.02 ± 0.40 | 0.0001*** |
| 36:2 PE | 9.93 ± 0.69 | 7 ± 0.03 | 0.0254* |
| 36:3 PE | 1.05 ± 0.10 | 1.32 ± 0.02 | ns |
| 36:4 PE | 2.80 ± 0.11 | 4.78 ± 0.14 | 0.0009*** |
| 36:5 PE | 0.34 ± 0.02 | 1.64 ± 0.08 | 0.0002*** |
| 38:1 PE | 0.69 ± 0.04 | 0.21 ± 0.02 | 0.0009*** |
| 38:2 PE | 1.55 ± 0.24 | 1.09 ± 0.07 | ns |
| 38:3 PE | 5.09 ± 0.54 | 5.28 ± 0.02 | ns |
| 38:4 PE | 30.45 ± 1.32 | 29.95 ± 0.17 | ns |
| 38:5 PE | 5.32 ± 0.16 | 5.13 ± 0.20 | ns |
| 38:6 PE | 0.7 ± 0.06 | 1.05 ± 0.09 | ns |
| 38:7 PE | 0.06 ± 0.01 | 0.29 ± 0.02 | 0.0006*** |
| 40:2 PE | 0.81 ± 0.06 | 0.24 ± 0.03 | 0.0021** |
| 40:3 PE | 1.14 ± 0.04 | 0.89 ± 0.15 | ns |
| 40:4 PE | 6.49 ± 0.49 | 2.88 ± 0.19 | 0.0048** |
| 40:5 PE | 1.62 ± 0.10 | 1.37 ± 0.04 | ns |
| 40:6 PE | 3.53 ± 0.15 | 3.11 ± 0.06 | ns |
| 40:7 PE | 0.69 ± 0.16 | 0.74 ± 0.05 | ns |
| 42:9 PE | 0.38 ± 0.03 | 0.28 ± 0.01 | 0.0491* |
| 42:10 PE | 0.08 ± 0.01 | 0.08 ± 0.01 | ns |
| PS | |||
| 34:0 PS | 0.67 ± 0.16 | 0.37 ± 0.05 | ns |
| 34:1 PS | 3.76 ± 1.87 | 5.65 ± 3.54 | ns |
| 34:2 PS | 0.79 ± 0.01 | 0.83 ± 0.1 | ns |
| 36:1 PS | 37.45 ± 1.23 | 34.94 ± 1.87 | ns |
| 36:2 PS | 6.82 ± 0.40 | 5.8 ± 0.4 | ns |
| 36:4 PS | 1.01 ± 0.04 | 1.27 ± 0.09 | ns |
| 38:1 PS | 2.37 ± 0.16 | 1.86 ± 0.08 | ns |
| 38:2 PS | 1.85 ± 0.26 | 1.77 ± 0.21 | ns |
| 38:3 PS | 6.64 ± 0.40 | 7.8 ± 0.38 | ns |
| 38:4 PS | 12.17 ± 0.96 | 14.7 ± 1.12 | ns |
| 38:5 PS | 1.30 ± 0.15 | 1.14 ± 0.08 | ns |
| 40:1 PS | 1.54 ± 0.10 | 1.11 ± 0.10 | ns |
| 40:2 PS | 1.28 ± 0.14 | 1.41 ± 0.16 | ns |
| 40:3 PS | 1.61 ± 0.32 | 2.95 ± 0.42 | ns |
| 40:4 PS | 9.86 ± 0.91 | 8.41 ± 0.36 | ns |
| 40:5 PS | 4.00 ± 0.80 | 4.05 ± 0.71 | ns |
| 40:6 PS | 4.27 ± 0.16 | 3.8 ± 0.15 | ns |
| 40:7 PS | 0.23 ± 0.02 | 0.19 ± 0.01 | ns |
| 42:5 PS | 0.77 ± 0.06 | 0.48 ± 0.07 | ns |
| 42:7 PS | 0.64 ± 0.07 | 0.49 ± 0.06 | ns |
| 42:8 PS | 0.54 ± 0.08 | 0.59 ± 0.05 | ns |
| 42:9 PS | 0.42 ± 0.02 | 0.39 ± 0.04 | ns |
| PC species | Preadipocyte (%) | Adipocyte (%) |
|---|---|---|
| PC | ||
| 16:0/16:0 PC | 12.78 | 8.97 |
| 16:0/16:1 PC | 4.13 | 7.45 |
| 16:0/18:0 PC | 13.16 | 12.58 |
| 16:0/18:1 PC | 24.17 | 20.64 |
| 16:0/18:2 PC | 3.58 | 4.32 |
| 16:0/18:3 PC | 2.51 | 3.33 |
| 16:0/20:4 PC | 1.12 | 2.11 |
| 16:0/22:6 PC | 0.43 | 0.42 |
| 18:0/18:1 PC | 4.57 | 4.73 |
| 18:0/18:2 PC | 2.61 | 4.21 |
| 18:0/18:3 PC | 6.63 | 7.31 |
| 18:0/20:4 PC | 1.37 | 2.61 |
| 18:0/22:6 PC | 2.06 | 1.71 |
| 18:1/18:1 PC | 7.90 | 5.51 |
| 18:1/18:2 PC | 10.91 | 11.24 |
| 18:1/18:3 PC | 1.66 | 2.28 |
| 18:1/20:4 PC | 0.42 | 0.60 |
| 18:1/22:6 PC | 12.78 | 8.97 |
| PE | ||
| 16:0/16:0 PE | 0.30 | 0.22 |
| 16:0/16:1 PE | 0.93 | 6.12 |
| 16:0/18:0 PE | 7.08 | 3.63 |
| 16:0/18:1 PE | 6.96 | 6.05 |
| 16:0/18:2 PE | 0.38 | 0.85 |
| 16:0/18:3 PE | 0.05 | 0.17 |
| 16:0/20:4 PE | 2.00 | 4.16 |
| 16:0/22:6 PE | 0.24 | 0.43 |
| 18:0/18:1 PE | 26.75 | 19.09 |
| 18:0/18:2 PE | 2.83 | 3.41 |
| 18:0/18:3 PE | 0.37 | 0.54 |
| 18:0/20:4 PE | 34.77 | 40.20 |
| 18:0/22:6 PE | 1.22 | 1.18 |
| 18:1/18:1 PE | 10.21 | 7.83 |
| 18:1/18:2 PE | 0.94 | 0.91 |
| 18:1/18:3 PE | 0.12 | 0.13 |
| 18:1/20:4 PE | 3.41 | 3.72 |
| 18:1/22:6 PE | 1.44 | 1.36 |
| Primers | Sequence |
|---|---|
| LPCAT1 forward | GTGCACGAGCTGCGACT |
| LPCAT1 reverse | GCTGCTCTGGCTCCTTATCA |
| LPCAT2 forward | GTCCAGCAGACTACGATCAGTG |
| LPCAT2 reverse | CTTATTGGATGGGTCAGCTTTTC |
| LPCAT3 forward | TCAGGATACCTGATTTGCTTCCA |
| LPCAT3 reverse | GGATGGTCTGTTGCACCAAGTAG |
| LPCAT4 forward | TTCGGTTTCAGAGGATACGACAA |
| LPCAT4 reverse | AATGTCTGGATTGTCGGACTGAA |
| LPEAT1 forward | CTGAAATGTGTGTGCTATGAGCG |
| LPEAT1 reverse | TGGAAGAGAGGAAGTGGTGTCTG |
| PPARγ2 forward | TATGCTGTTATGGGTGAAACTCTGG |
| PPARγ2 reverse | GTCAAAGGAATGCGAGTGGTCT |
| 36B4 forward | CTGAGATTCGGGATATGCTGTTG |
| 36B4 reverse | AAAGCCTGGAAGAAGGAGGTCTT |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Eto, M.; Shindou, H.; Koeberle, A.; Harayama, T.; Yanagida, K.; Shimizu, T. Lysophosphatidylcholine Acyltransferase 3 Is the Key Enzyme for Incorporating Arachidonic Acid into Glycerophospholipids during Adipocyte Differentiation. Int. J. Mol. Sci. 2012, 13, 16267-16280. https://doi.org/10.3390/ijms131216267
Eto M, Shindou H, Koeberle A, Harayama T, Yanagida K, Shimizu T. Lysophosphatidylcholine Acyltransferase 3 Is the Key Enzyme for Incorporating Arachidonic Acid into Glycerophospholipids during Adipocyte Differentiation. International Journal of Molecular Sciences. 2012; 13(12):16267-16280. https://doi.org/10.3390/ijms131216267
Chicago/Turabian StyleEto, Miki, Hideo Shindou, Andreas Koeberle, Takeshi Harayama, Keisuke Yanagida, and Takao Shimizu. 2012. "Lysophosphatidylcholine Acyltransferase 3 Is the Key Enzyme for Incorporating Arachidonic Acid into Glycerophospholipids during Adipocyte Differentiation" International Journal of Molecular Sciences 13, no. 12: 16267-16280. https://doi.org/10.3390/ijms131216267
APA StyleEto, M., Shindou, H., Koeberle, A., Harayama, T., Yanagida, K., & Shimizu, T. (2012). Lysophosphatidylcholine Acyltransferase 3 Is the Key Enzyme for Incorporating Arachidonic Acid into Glycerophospholipids during Adipocyte Differentiation. International Journal of Molecular Sciences, 13(12), 16267-16280. https://doi.org/10.3390/ijms131216267