Increased Insulin Sensitivity and Distorted Mitochondrial Adaptations during Muscle Unloading

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Gastrocnemius Relative Mass and Blood Metabolic Parameters
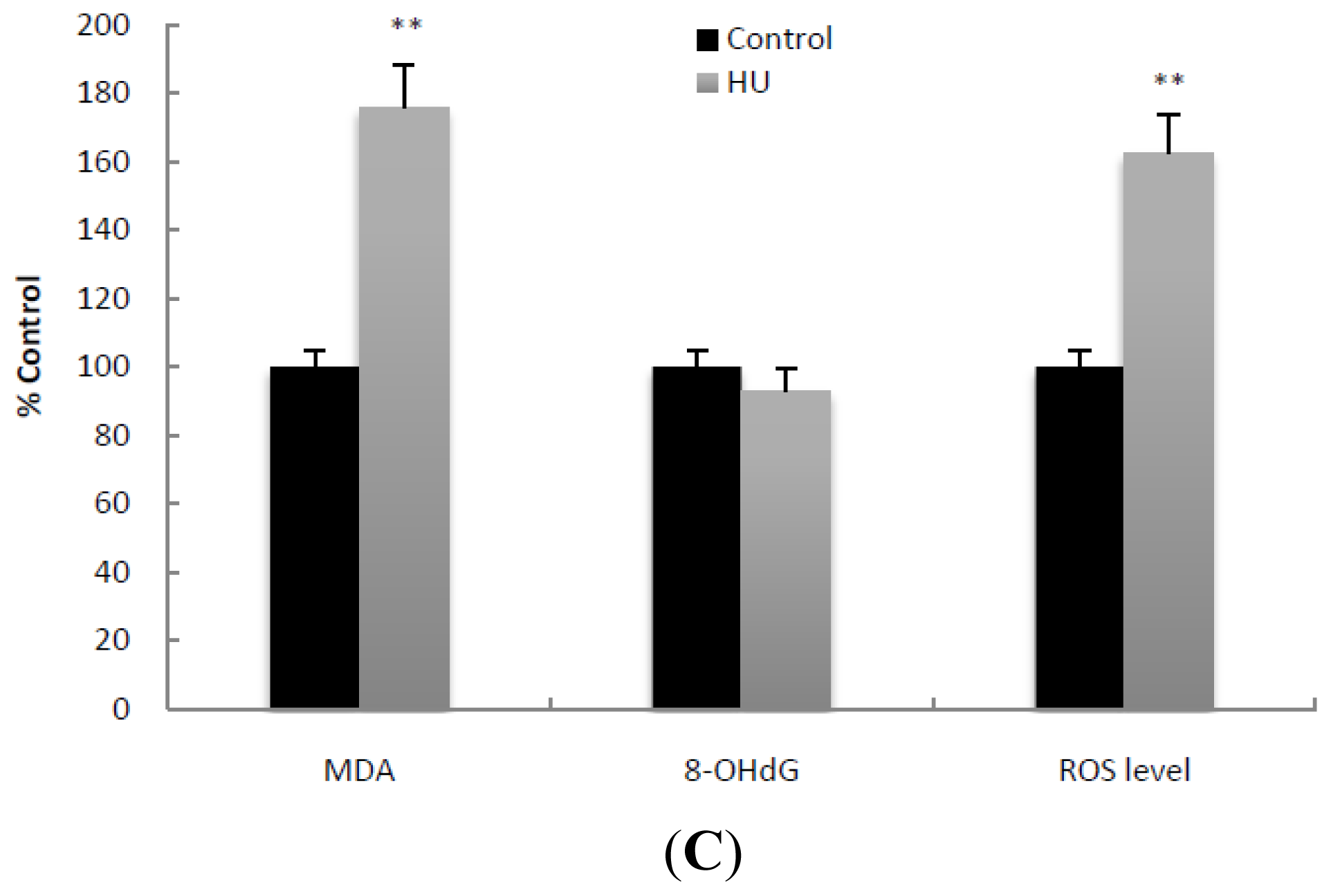
2.2. Mitochondrial Content and Oxidative Damage
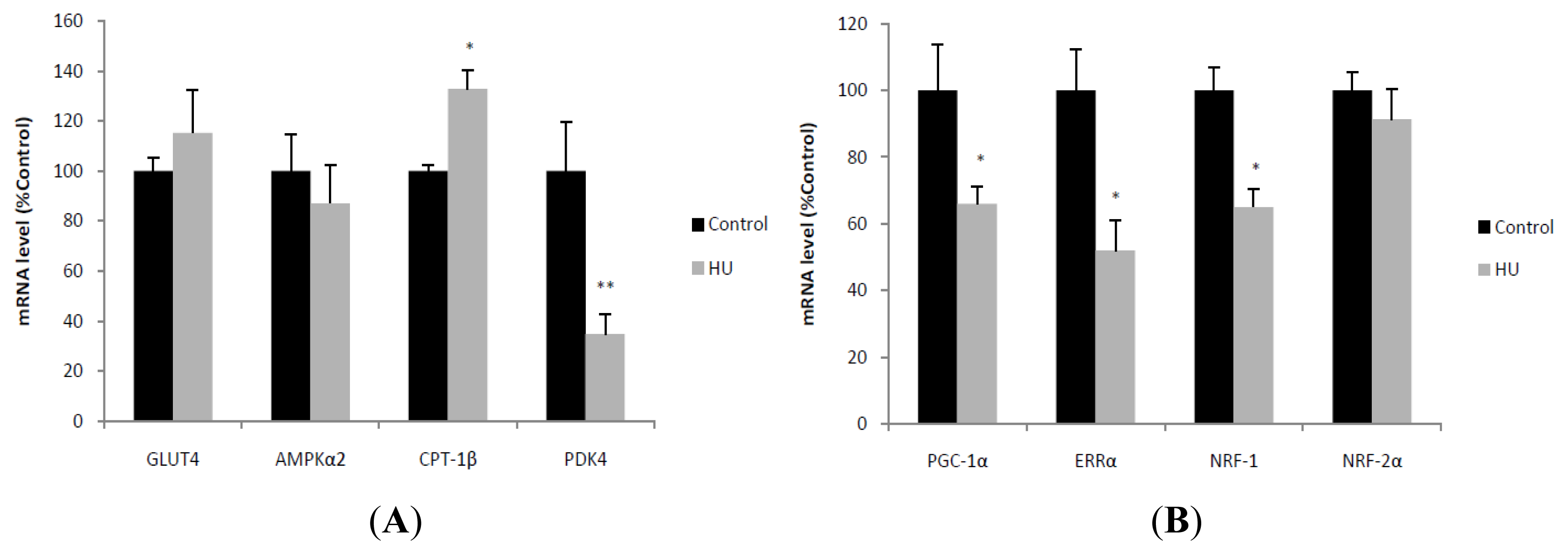
2.3. Expression of Genes Involved in Fuel Utilization, Mitochondrial Biogenesis and Dynamics
2.4. Mitochondrial Protein Level of PDK4, CPT-1beta, Mfn2 and Drp1
3. Materials and Methods
3.1. Animals and Hindlimb Unloading
3.2. Tissue Dissection and Blood Sampling
3.3. Subcellular Fractionation and ELISA for Cytochrome C and 8-OHdG
3.4. Immunoblotting
3.5. Real-time PCR and mtDNA Quantitation
3.6. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Information
ijms-13-16971-s001.pdfAcknowledgements
References
- Norman, T.L.; Bradley-Popovich, G.; Clovis, N.; Cutlip, R.G.; Brynerm, R.W. Aerobic exercise as a countermeasure for microgravity-induced bone loss and muscle atrophy in a rat hindlimb suspension model. Aviat. Space Environ. Med 2000, 71, 593–598. [Google Scholar]
- Shimada, Y.; Sakuraba, T.; Matsunaga, T.; Misawa, A.; Kawatani, M.; Itoi, E. Effects of therapeutic magnetic stimulation on acute muscle atrophy in rats after hindlimb suspension. Biomed. Res 2006, 27, 23–27. [Google Scholar]
- Eash, J.; Olsen, A.; Breur, G.; Gerrard, D.; Hannon, K. FGFR1 inhibits skeletal muscle atrophy associated with hindlimb suspension. BMC Musculoskelet Disord 2007, 8, 32. [Google Scholar]
- Brocca, L.; Pellegrino, M.A.; Desaphy, J.F.; Pierno, S.; Camerino, D.C.; Bottinelli, R. Is oxidative stress a cause or consequence of disuse muscle atrophy in mice? A proteomic approach in hindlimb-unloaded mice. Exp. Physiol 2010, 95, 331–350. [Google Scholar]
- Kyparos, A.; Feeback, D.L.; Layne, C.S.; Martinez, D.A.; Clarke, M.S. Mechanical stimulation of the plantar foot surface attenuates soleus muscle atrophy induced by hindlimb unloading in rats. J. Appl. Physiol 2005, 99, 739–746. [Google Scholar]
- Hurst, J.E.; Fitts, R.H. Hindlimb unloading-induced muscle atrophy and loss of function: Protective effect of isometric exercise. J. Appl. Physiol 2003, 95, 1405–1417. [Google Scholar]
- Carlson, C.J.; Booth, F.W.; Gordon, S.E. Skeletal muscle myostatin mRNA expression is fiber-type specific and increases during hindlimb unloading. Am. J. Physiol 1999, 277, R601–R606. [Google Scholar]
- Zhang, P.; Chen, X.; Fan, M. Signaling mechanisms involved in disuse muscle atrophy. Med. Hypotheses 2007, 69, 310–321. [Google Scholar]
- Stein, T.P.; Wade, C.E. Metabolic consequences of muscle disuse atrophy. J. Nutr 2005, 135, 1824S–1828S. [Google Scholar]
- Mazzatti, D.J.; Smith, M.A.; Oita, R.C.; Lim, F.L.; White, A.J.; Reid, M.B. Muscle unloading-induced metabolic remodeling is associated with acute alterations in PPARdelta and UCP-3 expression. Physiol. Genomics 2008, 34, 149–161. [Google Scholar]
- Leary, S.C.; Lyons, C.N.; Rosenberger, A.G.; Ballantyne, J.S.; Stillman, J.; Moyes, C.D. Fiber-type differences in muscle mitochondrial profiles. Am. J. Physiol. Regul. Integr. Comp. Physiol 2003, 285, R817–R826. [Google Scholar]
- Lin, J.; Wu, H.; Tarr, P.T.; Zhang, C.Y.; Wu, Z.; Boss, O.; Michael, L.F.; Puigserver, P.; Isotani, E.; Olson, E.N.; et al. Transcriptional co-activator PGC-1 alpha drives the formation of slow-twitch muscle fibres. Nature 2002, 418, 797–801. [Google Scholar]
- Handschin, C.; Chin, S.; Li, P.; Liu, F.; Maratos-Flier, E.; Lebrasseur, N.K.; Yan, Z.; Spiegelman, B.M. Skeletal muscle fiber-type switching, exercise intolerance, and myopathy in PGC-1alpha muscle-specific knock-out animals. J. Biol. Chem 2007, 282, 30014–30021. [Google Scholar]
- Zechner, C.; Lai, L.; Zechner, J.F.; Geng, T.; Yan, Z.; Rumsey, J.W.; Collia, D.; Chen, Z.; Wozniak, D.F.; Leone, T.C.; et al. Total skeletal muscle PGC-1 deficiency uncouples mitochondrial derangements from fiber type determination and insulin sensitivity. Cell Metab 2010, 12, 633–642. [Google Scholar]
- Yan, Z.; Okutsu, M.; Akhtar, Y.N.; Lira, V.A. Regulation of exercise-induced fiber type transformation, mitochondrial biogenesis, and angiogenesis in skeletal muscle. J. Appl. Physiol 2011, 110, 264–274. [Google Scholar]
- Geng, T.; Li, P.; Okutsu, M.; Yin, X.; Kwek, J.; Zhang, M.; Yan, Z. PGC-1alpha plays a functional role in exercise-induced mitochondrial biogenesis and angiogenesis but not fiber-type transformation in mouse skeletal muscle. Am. J. Physiol. Cell Physiol 2010, 298, C572–C579. [Google Scholar]
- Summermatter, S.; Thurnheer, R.; Santos, G.; Mosca, B.; Baum, O.; Treves, S.; Hoppeler, H.; Zorzato, F.; Handschin, C. Remodeling of calcium handling in skeletal muscle through PGC-1alpha: Impact on force, fatigability, and fiber type. Am. J. Physiol. Cell Physiol 2012, 302, C88–C99. [Google Scholar]
- Rangwala, S.M.; Li, X.; Lindsley, L.; Wang, X.; Shaughnessy, S.; Daniels, T.G.; Szustakowski, J.; Nirmala, N.R.; Wu, Z.; Stevenson, S.C. Estrogen-related receptor alpha is essential for the expression of antioxidant protection genes and mitochondrial function. Biochem. Biophys. Res. Commun 2007, 357, 231–236. [Google Scholar]
- Schreiber, S.N.; Emter, R.; Hock, M.B.; Knutti, D.; Cardenas, J.; Podvinec, M.; Oakeley, E.J.; Kralli, A. The estrogen-related receptor alpha (ERRalpha) functions in PPARgamma coactivator 1alpha (PGC-1alpha)-induced mitochondrial biogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 6472–6477. [Google Scholar]
- Cartoni, R.; Leger, B.; Hock, M.B.; Praz, M.; Crettenand, A.; Pich, S.; Ziltener, J.L.; Luthi, F.; Deriaz, O.; Zorzano, A.; et al. Mitofusins 1/2 and ERRalpha expression are increased in human skeletal muscle after physical exercise. J. Physiol 2005, 567, 349–358. [Google Scholar]
- Soriano, F.X.; Liesa, M.; Bach, D.; Chan, D.C.; Palacin, M.; Zorzano, A. Evidence for a mitochondrial regulatory pathway defined by peroxisome proliferator-activated receptor-gamma coactivator-1 alpha, estrogen-related receptor-alpha, and mitofusin 2. Diabetes 2006, 55, 1783–1791. [Google Scholar]
- Araki, M.; Motojima, K. Identification of ERRalpha as a specific partner of PGC-1alpha for the activation of PDK4 gene expression in muscle. FEBS J 2006, 273, 1669–1680. [Google Scholar]
- Wende, A.R.; Huss, J.M.; Schaeffer, P.J.; Giguere, V.; Kelly, D.P. PGC-1alpha coactivates PDK4 gene expression via the orphan nuclear receptor ERRalpha: A mechanism for transcriptional control of muscle glucose metabolism. Mol. Cell Biol 2005, 25, 10684–10694. [Google Scholar]
- Pellegrino, M.A.; Desaphy, J.F.; Brocca, L.; Pierno, S.; Camerino, D.C.; Bottinelli, R. Redox homeostasis, oxidative stress and disuse muscle atrophy. J. Physiol 2011, 589, 2147–2160. [Google Scholar]
- Siu, P.M.; Pistilli, E.E.; Always, S.E. Apoptotic responses to hindlimb suspension in gastrocnemius muscles from young adult and aged rats. Am. J. Physiol. Regul. Integr. Comp. Physiol 2005, 289, R1015–R1026. [Google Scholar]
- Quadrilatero, J.; Rush, J.W. Increased DNA fragmentation and altered apoptotic protein levels in skeletal muscle of spontaneously hypertensive rats. J. Appl. Physiol 2006, 101, 1149–1161. [Google Scholar]
- Ding, H.; Jiang, N.; Liu, H.; Liu, X.; Liu, D.; Zhao, F.; Wen, L.; Liu, S.; Ji, L.L.; Zhang, Y. Response of mitochondrial fusion and fission protein gene expression to exercise in rat skeletal muscle. Biochim. Biophys. Acta 2010, 1800, 250–256. [Google Scholar]
- O’Neill, B.T.; Kim, J.; Wende, A.R.; Theobald, H.A.; Tuinei, J.; Buchanan, J.; Guo, A.; Zaha, V.G.; Davis, D.K.; Schell, J.C.; et al. A conserved role for phosphatidylinositol 3-kinase but not Akt signaling in mitochondrial adaptations that accompany physiological cardiac hypertrophy. Cell Metab 2007, 6, 294–306. [Google Scholar]
- Langfort, J.; Zernicka, E.; Mayet-Sornay, M.H.; Dubaniewicz, A.; Desplanches, D. Effects of acute and chronic hindlimb suspension on sensitivity and responsiveness to insulin in the rat soleus muscle. Biochem. Cell Biol 1997, 75, 41–44. [Google Scholar]
- Henriksen, E.J.; Tischler, M.E. Time course of the response of carbohydrate metabolism to unloading of the soleus. Metabolism 1988, 37, 201–208. [Google Scholar]
- Loh, K.; Deng, H.; Fukushima, A.; Cai, X.; Boivin, B.; Galic, S.; Bruce, C.; Shields, B.J.; Skiba, B.; Ooms, L.M.; et al. Reactive oxygen species enhance insulin sensitivity. Cell Metab 2009, 10, 260–272. [Google Scholar]
- Strobel, N.A.; Peake, J.M.; Matsumoto, A.; Marsh, S.A.; Coombes, J.S.; Wadley, G.D. Antioxidant supplementation reduces skeletal muscle mitochondrial biogenesis. Med. Sci. Sports Exerc 2011, 43, 1017–1024. [Google Scholar]
- Gomez-Cabrera, M.C.; Domenech, E.; Romagnoli, M.; Arduini, A.; Borras, C.; Pallardo, F.V.; Sastre, J.; Vina, J. Oral administration of vitamin C decreases muscle mitochondrial biogenesis and hampers training-induced adaptations in endurance performance. Am. J. Clin. Nutr 2008, 87, 142–149. [Google Scholar]
- Canepari, M.; Pellegrino, M.A.; D’Antona, G.; Bottinelli, R. Single muscle fiber properties in aging and disuse. Scand. J. Med. Sci. Sports 2010, 20, 10–19. [Google Scholar]
- Ohira, Y.; Yasui, W.; Kariya, F.; Wakatsuki, T.; Nakamura, K.; Asakura, T.; Edgerton, V.R. Metabolic adaptation of skeletal muscles to gravitational unloading. Acta Astronaut 1994, 33, 113–117. [Google Scholar]
- Grichko, V.P.; Heywood-Cooksey, A.; Kidd, K.R.; Fitts, R.H. Substrate profile in rat soleus muscle fibers after hindlimb unloading and fatigue. J. Appl. Physiol 2000, 88, 473–478. [Google Scholar]
- Nahle, Z.; Hsieh, M.; Pietka, T.; Coburn, C.T.; Grimaldi, P.A.; Zhang, M.Q.; Das, D.; Abumrad, N.A. CD36-dependent regulation of muscle FoxO1 and PDK4 in the PPAR delta/beta-mediated adaptation to metabolic stress. J. Biol. Chem 2008, 283, 14317–14326. [Google Scholar]
- Zhang, Y.; Ma, K.; Sadana, P.; Chowdhury, F.; Gaillard, S.; Wang, F.; Mcdonnell, D.P.; Unterman, T.G.; Elam, M.B.; Park, E.A. Estrogen-related receptors stimulate pyruvate dehydrogenase kinase isoform 4 gene expression. J. Biol. Chem 2006, 281, 39897–39906. [Google Scholar]
- Peters, S.J.; Harris, R.A.; Heigenhauser, G.J.; Spriet, L.L. Muscle fiber type comparison of PDH kinase activity and isoform expression in fed and fasted rats. Am. J. Physiol. Regul. Integr. Comp. Physiol 2001, 280, R661–R668. [Google Scholar]
- Nikolic, N.; Rhedin, M.; Rustan, A.C.; Storlien, L.; Thoresen, G.H.; Stromstedt, M. Overexpression of PGC-1alpha increases fatty acid oxidative capacity of human skeletal muscle cells. Biochem. Res. Int 2012, 2012, 714074. [Google Scholar]
- Han, B.; Zhu, M.J.; Ma, C.; Du, M. Rat hindlimb unloading down-regulates insulin like growth factor-1 signaling and AMP-activated protein kinase, and leads to severe atrophy of the soleus muscle. Appl. Physiol. Nutr. Metab 2007, 32, 1115–1123. [Google Scholar]
- Nakao, R.; Hirasaka, K.; Goto, J.; Ishidoh, K.; Yamada, C.; Ohno, A.; Okumura, Y.; Nonaka, I.; Yasutomo, K.; Baldwin, K.M.; et al. Ubiquitin ligase Cbl-b is a negative regulator for insulin-like growth factor 1 signaling during muscle atrophy caused by unloading. Mol. Cell Biol 2009, 29, 4798–4811. [Google Scholar]
- Pesce, V.; Cormio, A.; Fracasso, F.; Lezza, A.M.; Cantatore, P.; Gadaleta, M.N. Rat hindlimb unloading: Soleus and extensor digitorum longus histochemistry, mitochondrial DNA content and mitochondrial DNA deletions. Biosci. Rep 2002, 22, 115–125. [Google Scholar]
- Wagatsuma, A.; Kotake, N.; Kawachi, T.; Shiozuka, M.; Yamada, S.; Matsuda, R. Mitochondrial adaptations in skeletal muscle to hindlimb unloading. Mol. Cell Biochem 2011, 350, 1–11. [Google Scholar]
- Oishi, Y.; Ogata, T.; Yamamoto, K.I.; Terada, M.; Ohira, T.; Ohira, Y.; Taniguchi, K.; Roy, R.R. Cellular adaptations in soleus muscle during recovery after hindlimb unloading. Acta Physiol. (Oxf.) 2008, 192, 381–395. [Google Scholar]
- Pitts, K.R.; Mcniven, M.A.; Yoon, Y. Mitochondria-specific function of the dynamin family protein DLP1 is mediated by its C-terminal domains. J. Biol. Chem 2004, 279, 50286–50294. [Google Scholar]
- Lackner, L.L.; Horner, J.S.; Nunnari, J. Mechanistic analysis of a dynamin effector. Science 2009, 325, 874–877. [Google Scholar]
- Romanello, V.; Guadagnin, E.; Gomes, L.; Roder, I.; Sandri, C.; Petersen, Y.; Milan, G.; Masiero, E.; Del, P.P.; Foretz, M.; et al. Mitochondrial fission and remodelling contributes to muscle atrophy. EMBO J 2010, 29, 1774–1785. [Google Scholar]
- Zorzano, A.; Liesa, M.; Sebastian, D.; Segales, J.; Palacin, M. Mitochondrial fusion proteins: Dual regulators of morphology and metabolism. Semin. Cell Dev. Biol 2010, 21, 566–574. [Google Scholar]
- Pich, S.; Bach, D.; Briones, P.; Liesa, M.; Camps, M.; Testar, X.; Palacin, M.; Zorzano, A. The Charcot-Marie-Tooth type 2A gene product, Mfn2, up-regulates fuel oxidation through expression of OXPHOS system. Hum. Mol. Genet 2005, 14, 1405–1415. [Google Scholar]
- Chen, X.; Xu, Y. Liver-specific reduction of Mfn2 protein by RNAi results in impaired glycometabolism and lipid homeostasis in BALB/c mice. J. Huazhong Univ. Sci. Technol. Med. Sci 2009, 29, 689–696. [Google Scholar]
- Bach, D.; Pich, S.; Soriano, F.X.; Vega, N.; Baumgartner, B.; Oriola, J.; Daugaard, J.R.; Lloberas, J.; Camps, M.; Zierath, J.R.; et al. Mitofusin-2 determines mitochondrial network architecture and mitochondrial metabolism. A novel regulatory mechanism altered in obesity. J. Biol. Chem 2003, 278, 17190–17197. [Google Scholar]
- Zorzano, A. Regulation of mitofusin-2 expression in skeletal muscle. Appl. Physiol. Nutr. Metab 2009, 34, 433–439. [Google Scholar]
- Zorzano, A.; Hernandez-Alvarez, M.I.; Palacin, M.; Mingrone, G. Alterations in the mitochondrial regulatory pathways constituted by the nuclear co-factors PGC-1alpha or PGC-1beta and mitofusin 2 in skeletal muscle in type 2 diabetes. Biochim. Biophys. Acta 2010, 1797, 1028–1033. [Google Scholar]







© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Qi, Z.; Zhang, Y.; Guo, W.; Ji, L.; Ding, S. Increased Insulin Sensitivity and Distorted Mitochondrial Adaptations during Muscle Unloading. Int. J. Mol. Sci. 2012, 13, 16971-16985. https://doi.org/10.3390/ijms131216971
Qi Z, Zhang Y, Guo W, Ji L, Ding S. Increased Insulin Sensitivity and Distorted Mitochondrial Adaptations during Muscle Unloading. International Journal of Molecular Sciences. 2012; 13(12):16971-16985. https://doi.org/10.3390/ijms131216971
Chicago/Turabian StyleQi, Zhengtang, Yuan Zhang, Wei Guo, Liu Ji, and Shuzhe Ding. 2012. "Increased Insulin Sensitivity and Distorted Mitochondrial Adaptations during Muscle Unloading" International Journal of Molecular Sciences 13, no. 12: 16971-16985. https://doi.org/10.3390/ijms131216971
APA StyleQi, Z., Zhang, Y., Guo, W., Ji, L., & Ding, S. (2012). Increased Insulin Sensitivity and Distorted Mitochondrial Adaptations during Muscle Unloading. International Journal of Molecular Sciences, 13(12), 16971-16985. https://doi.org/10.3390/ijms131216971