Plant Glandular Trichomes as Targets for Breeding or Engineering of Resistance to Herbivores



Abstract
:1. Introduction
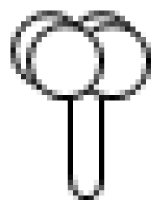
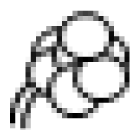
2. Trichome Morphology in Mint, Basil and Tomato
3. Biosynthesis and Function of Glandular Trichome-Produced Compounds
3.1. Hormonal Regulation of Induced Defenses in Trichomes
3.2. Terpenes
3.3. Phenylpropenes
3.4. Flavonoids
3.5. Methyl Ketones
3.6. Acyl Sugars
3.7. Defensive Proteins
4. Identification of Biochemical Pathways in Glandular Trichomes
5. Trichome Engineering to Increase Plant Resistance
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Wagner, G.J.; Wang, E.; Shepherd, R.W. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot 2004, 93, 3–11. [Google Scholar]
- Payne, W.W. A glossary of plant hair terminology. Brittonia 1978, 30, 239–255. [Google Scholar]
- Reis, C.; Sajo, M.G.; Stehmann, J.R. Leaf structure and taxonomy of Petunia and Calibrachoa (Solanaceae). Braz. Arch. Biol. Techol 2002, 45, 59–66. [Google Scholar]
- Werker, E. Trichome diversity and development. In Plant Trichomes; Hallahan, D.L., Gray, J.C., Eds.; Academic Press: New York, NY, USA, 2000; p. 1. [Google Scholar]
- Mathur, J.; Chua, N. Microtubule stabilization leads to growth reorientation in Arabidopsis trichomes. Plant Cell 2000, 12, 465–477. [Google Scholar]
- Schnittger, A.; Hülskamp, M. Trichome morphogenesis: A cell-cycle perspective. Phil. Trans. R. Soc. Lond. B 2002, 357, 823–826. [Google Scholar]
- Larkin, J.C.; Brown, M.L.; Schiefelbein, J. How do cells know what they want to be when they grow up? Lessons from epidermal patterning in Arabidopsis. Annu. Rev. Plant Biol 2003, 54, 403–430. [Google Scholar]
- Fahn, A. Structure and function of secretory cells. In Plant Trichomes; Hallahan, D.L., Gray, J.C., Eds.; Academic Press: New York, NY, USA, 2000; p. 37. [Google Scholar]
- Schilmiller, A.L.; Last, R.L.; Pichersky, E. Harnessing plant trichome biochemistry for the production of useful compounds. Plant J 2008, 54, 702–711. [Google Scholar]
- Duke, S.O.; Canel, C.; Rimando, A.M.; Tellez, M.R.; Duke, M.V.; Paul, R.N. Current and potential exploitation of plant glandular trichome productivity. Adv. Bot. Res 2000, 31, 121–151. [Google Scholar]
- Aharoni, A.; Jongsma, M.A.; Kim, T.; Ri, M.; Giri, A.P.; Verstappen, F.W.A.; Schwab, W.; Bouwmeester, H.J. Metabolic engineering of terpenoid biosynthesis in plants. Phytochem. Rev 2006, 5, 49–58. [Google Scholar]
- Weathers, P.J.; Arsenault, P.R.; Covello, P.S.; McMickle, A.; Teoh, K.H.; Reed, D.W. Artemisinin production in Artemisia annua: Studies in planta and results of a novel delivery method for treating malaria and other neglected diseases. Phytochem. Rev 2011, 10, 173–183. [Google Scholar]
- Mellon, J.E.; Zelaya, C.A.; Dowd, M.K.; Beltz, S.B.; Klich, M.A. Inhibitory effects of gossypol, gossypolone, and apogossypolone on a collection of economically important filamentous fungi. J. Agric. Food Chem 2012, 60, 2740–2745. [Google Scholar]
- Dayan, F.E.; Duke, S.O. Trichomes and root hairs: Natural pesticide factories. Pestic. Outlook 2003, 4, 175–178. [Google Scholar]
- Lange, B.M.; Mahmoud, S.S.; Wildung, M.R.; Turner, G.W.; Davis, E.M.; Lange, I.; Baker, R.C.; Boydston, R.A.; Croteau, R.B. Improving peppermint essential oil yield and composition by metabolic engineering. Proc. Natl. Acad. Sci. USA 2011, 108, 16944–16949. [Google Scholar]
- Tissier, A. Glandular trichomes: What comes after expressed sequence tags? Plant J 2012, 70, 51–68. [Google Scholar]
- Maffei, M.E. Sites of synthesis, biochemistry and functional role of plant volatiles. S. Afr. J. Bot 2010, 76, 612–631. [Google Scholar]
- Turner, G.W.; Gershenzon, J.; Croteau, R.B. Distribution of peltate glandular trichomes on developing leaves of peppermint. Plant Physiol 2000, 124, 655–663. [Google Scholar]
- Fahn, A. Secretory tissues in vascular plants. New Phytol 1988, 108, 229–257. [Google Scholar]
- Luckwill, L.C. The genus Lycopersicon: A historical, biological and taxonomic survey of the wild and cultivated tomato. Aberd. Univ. Stud 1943, 120, 1–44. [Google Scholar]
- Channarayappa, S.G.; Muniyappa, V.; Frist, R.H. Resistance of Lycopersicon species to Bemisia tabaci, a tomato leaf curl virus vector. Can. J. Bot 1992, 70, 2184–2192. [Google Scholar]
- McDowell, E.T.; Kapteyn, J.; Schmidt, A.; Li, C.; Kang, J.; Descour, A.; Shi, F.; Larson, M.; Schilmiller, A.; An, L.; et al. Comparative functional genomic analysis of Solanum glandular trichome types. Plant Physiol 2011, 155, 524–539. [Google Scholar]
- Li, L.; Zhao, Y.; McCaig, B.C.; Wingerd, B.A.; Wang, J.; Whalon, M.E.; Pichersky, E.; Howe, G.A. The tomato homolog of CORONATINE-INSENSITIVE1 is required for the maternal control of seed maturation, jasmonate-signaled defense responses, and glandular trichome development. Plant Cell 2004, 16, 126–143. [Google Scholar]
- Schilmiller, A.L.; Schauvinhold, I.; Larson, M.; Xu, R.; Charbonneau, A.L.; Schmidt, A.; Wilkerson, C.; Last, R.L.; Pichersky, E. Monoterpenes in the glandular trichomes of tomato are synthesized from a neryl diphosphate precursor rather than geranyl diphosphate. Proc. Natl. Acad. Sci. USA 2009, 106, 10865–10870. [Google Scholar]
- Kang, J.; Shi, F.; Jones, A.D.; Marks, M.D.; Howe, G.A. Distortion of trichome morphology by the hairless mutation of tomato affects leaf surface chemistry. J. Exp. Bot 2010, 61, 1053–1064. [Google Scholar]
- Simmons, A.T.; Gurr, G.M. Trichomes of Lycopersicon species and their hybrids: Effects on pests and natural enemies. Agric. Forest Entomol 2005, 7, 265–276. [Google Scholar]
- Wilkens, R.T.; Shea, G.O.; Halbreich, S.; Stamp, N.E. Resource availability and the trichome defenses of tomato plants. Oecologia 1996, 106, 181–191. [Google Scholar]
- Fernández-Muñoz, R.; Salinas, M.; Álvarez, M.; Cuartero, J. Inheritance of resistance to two-spotted spider mite and glandular leaf trichomes in wild tomato Lycopersicon pimpinellifolium (Jusl.) Mill. J. Am. Soc. Hortic. Sci 2003, 128, 188–195. [Google Scholar]
- Fordyce, J.A.; Agrawal, A.A. The role of plant trichomes and caterpillar group size on growth and defence of the pipevine swallowtail Battus phineor. J. Anim. Ecol 2001, 70, 997–1005. [Google Scholar]
- Kennedy, G.G. Tomato, pests, parasitoids, and predators: Tritrophic interactions involving the genus Lycopersicon. Annu. Rev. Entomol 2003, 48, 51–72. [Google Scholar]
- Wei, J.; Yan, L.; Ren, Q.; Li, C.; Ge, F.; Kang, L.E. Antagonism between herbivore-induced plant volatiles and trichomes affects tritrophic interactions. Plant Cell Environ. 2012. [Google Scholar] [CrossRef]
- Cardoso, M.Z. Herbivore handling of a plant’s trichome: The case of Heliconius charithonia (L.) (Lepidoptera: Nymphalidae) and Passiflora lobata (Killip) Hutch. (Passifloraceae). Neotrop. Entomol 2008, 37, 247–252. [Google Scholar]
- Pott, C.; McLoughlin, S.; Shunqing, W.; Friis, E.M. Trichomes on the leaves of Anomozamites villosus sp. nov. (Bennettitales) from the Daohugou beds (Middle Jurassic), Inner Mongolia, China: Mechanical defence against herbivorous arthropods. Rev. Paleobot. Palyno 2012, 169, 48–60. [Google Scholar]
- Simmons, A.T.; Gurr, G.M.; McGrath, D.; Martin, P.M.; Nicol, H.I. Entrapment of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) on glandular trichomes of Lycopersicon species. Austr. J. Entomol 2004, 43, 196–200. [Google Scholar]
- Puterka, G.J.; Farone, W.; Palmer, T.; Barrington, A. Structure-function relationships affecting the insecticidal and miticidal activity of sugar esters. J. Econ. Entomol 2003, 96, 636–644. [Google Scholar]
- Ramirez, A.M.; Stoopen, G.; Menzel, T.R.; Gols, R.; Bouwmeester, H.J.; Dicke, M.; Jongsma, M.A. Bidirectional secretions from glandular trichomes of Pyrethrum enable immunization of seedlings. Plant Cell 2012, 24, 4252–4265. [Google Scholar]
- Shepherd, R.W.; Bass, W.T.; Houtz, R.L.; Wagner, G.J. Phylloplanins of tobacco are defensive proteins deployed on aerial surfaces by short glandular trichomes. Plant Cell 2005, 17, 1851–1861. [Google Scholar]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol 2007, 3, 408–414. [Google Scholar]
- Gang, D.R.; Wang, J.; Dudareva, N.; Nam, K.H.; Simon, J.E.; Lewinsohn, E.; Pichersky, E. An investigation of the storage and biosynthesis of phenylpropenes in sweet basil. Plant Physiol 2001, 125, 539–555. [Google Scholar]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett 2006, 4, 147–157. [Google Scholar]
- Fridman, E.; Wang, J.; Iijima, Y.; Froehlich, J.E.; Gang, D.R.; Ohlrogge, J.; Pichersky, E. Metabolic, genomic and biochemical analyses of glandular trichomes from the wild tomato species Lycopersicon hirsutum identify a key enzyme in the biosynthesis of methyl ketones. Plant Cell 2005, 17, 1252–1267. [Google Scholar]
- Kroumova, A.B.; Wagner, G.G. Different elongation pathways in the biosynthesis of acyl groups of trichome exudate sugar esters from various solanaceous plants. Planta 2003, 216, 1013–1021. [Google Scholar]
- Johnson, J.B. Plant pubescence: An ecological perspective. Bot. Rev 1975, 41, 233–258. [Google Scholar]
- Karabourniotis, G.; Kotsabassidis, D.; Manetas, Y. Trichome density and its protective potential against ultraviolet-B radiation damage during leaf development. Can. J. Bot 1995, 73, 376–383. [Google Scholar]
- Karabourniotis, G.; Kofidis, G.; Fasseas, C.; Liakoura, V.; Drossopoulos, I. Polyphenol deposition in leaf hairs of Olea europaea (Oleaceae) and Quercus ilex (Fagaceae). Am. J. Bot 1998, 85, 1007–1012. [Google Scholar]
- Ehleringer, J. Ecology and ecophysiology of leaf pubescence in North American desert plants. In Biology and Chemistry of Plant Trichomes; Rodriguez, E., Healey, P.L., Mehta, I., Eds.; Plenum Press: New York, NY, USA, 1984; pp. 113–132. [Google Scholar]
- De Silva, D.L.R.; Hetherington, A.M.; Mansfield, T.A. Where does all the calcium go? Evidence of an important regulatory role for trichomes in two calcicoles. Plant Cell Environ 1996, 19, 880–886. [Google Scholar]
- Sarret, G.; Harada, E.; Choi, Y.; Isaure, M.; Geoffroy, N.; Fakra, S.; Markus, M.A.; Birschwilks, M.; Clemens, S.; Manceau, A. Trichomes of tobacco excrete zinc as zinc-substituted calcium carbonate and other zinc-containing compounds. Plant Physiol 2006, 141, 1021–1034. [Google Scholar]
- Kant, M.R.; Ament, K.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Differential timing of spider mite-induced direct and indirect defenses in tomato plants. Plant Physiol 2004, 135, 483–495. [Google Scholar]
- Peiffer, M.; Tooker, J.F.; Luthe, D.S.; Felton, G.W. Plants on early alert: Glandular trichomes as sensors for insect herbivores. New Phytol 2009, 184, 644–656. [Google Scholar]
- Traw, M.B.; Dawson, T.E. Differential induction of trichomes by three herbivores of black mustard. Oecologia 2002, 131, 526–532. [Google Scholar]
- Boughton, A.J.; Hoover, K.; Felton, G.W. Methyl jasmonate application induces increased densities of glandular trichomes on tomato, Lycopersicon esculentum. J. Chem. Ecol 2005, 31, 2211–2216. [Google Scholar]
- Tian, D.; Tooker, J.; Peiffer, M.; Chung, S.; Felton, G.W. Role of trichomes in defense against herbivores: Comparison of herbivore response to woolly and hairless trichome mutants in tomato (Solanum lycopersicum). Planta 2012, 236, 1053–1066. [Google Scholar]
- Van Schie, C.C.N.; Haring, M.A.; Schuurink, R.C. Tomato linalool synthase is induced in glandular trichomes by jasmonic acid. Plant Mol. Biol 2007, 64, 251–263. [Google Scholar]
- Hare, J.D.; Walling, L.L. Constitutive and jasmonate-inducible traits of Datura wrightii. J. Chem. Ecol 2006, 32, 29–47. [Google Scholar]
- Laue, G.; Preston, C.A.; Baldwin, I.T. Fast track to the trichome: Induction of N-acyl nornicotines precedes nicotine induction in Nicotiana repanda. Planta 2000, 210, 510–514. [Google Scholar]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci 2006, 25, 417–440. [Google Scholar]
- Kant, M.R.; Bleeker, P.M.; Van Wijk, M.; Schuurink, R.C.; Haring, M.A. Plant volatiles in defence. In Advances in Botanical Research; Van Loon, L.C., Ed.; Academic Press: Burlington, VT, USA, 2009; Volume 51, pp. 613–666. [Google Scholar]
- Connolly, J.D.; Hill, R.A. Dictionary of Terpenoids; Chapman and Hall: London, UK; p. 1991.
- Matusova, R.; Rani, K.; Verstappen, F.W.A.; Franssen, M.C.R.; Beale, M.H.; Bouwmeester, H.J. The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche spp. are derived from the carotenoid pathway. Plant Physiol 2005, 139, 920–934. [Google Scholar]
- Vranová, E.; Coman, D.; Gruissem, W. Structure and dynamics of the isoprenoid pathway network. Mol. Plant 2012, 5, 318–333. [Google Scholar]
- Croteau, R.B.; Davis, E.M.; Ringer, K.L.; Wildung, M.R. (-)-Menthol biosynthesis and molecular genetics. Naturwissenschaften 2005, 92, 562–577. [Google Scholar]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol 2006, 9, 297–304. [Google Scholar]
- Hoeffler, J.F.; Herrerlin, A.; Grosdemange-Billiard, C.; Bach, T.J.; Rohmer, M. Isoprenoid biosynthesis in higher plants and in Escherichia coli. On the branching in the methylerythritol phosphate pathway and the independent biosynthesis of isopentenyl diphosphate and dimethylallyl diphosphate. Biochem J 2002, 366, 573–583. [Google Scholar]
- Rodríguez-Concepcíon, M.; Boronat, A. Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids. A metabolic milestone achieved through genomics. Plant Physiol 2002, 130, 1079–1089. [Google Scholar]
- McGarvey, D.J.; Croteau, R. Terpenoid metabolism. Plant Cell 1995, 7, 1015–1026. [Google Scholar]
- Tholl, D.; Lee, S. Terpene specialized metabolism inArabidopsis thaliana. Arabidopsis Book 2011, 9. [Google Scholar] [CrossRef]
- Nakamura, A.; Shimada, H.; Masuda, T.; Ohta, H.; Takamiya, K.-I. Two distinct isopentenyl diphosphate isomerases in cytosol and plastid are differentially induced by environmental stresses in tobacco. FEBS Lett 2001, 506, 61–64. [Google Scholar]
- Phillips, M.A.; D’Auria, J.C.; Gershenzon, J.; Pichersky, E. The Arabidopsis thaliana type I isopentenyl diphosphate isomerases are targeted to multiple subcellular compartments and have overlapping functions in isoprenoid biosynthesis. Plant Cell 2008, 20, 677–696. [Google Scholar]
- Sun, J.; Zhang, Y.-Y.; Liu, H.; Zou, Z.; Zhang, C.-J.; Zhang, X.-H.; Li, H.-X.; Ye, Z.-B. A novel cytoplasmic isopentenyl diphosphate isomerase gene from tomato (Solanum lycopersicum): Cloning, expression, and color complementation. Plant Mol. Biol. Rep 2010, 28, 473–480. [Google Scholar]
- Nielsen, M.T.; Akers, C.P.; Järlfors, U.E.; Wagner, G.J.; Berger, S. Comparative ultrastructural features of secreting and nonsecreting glandular trichomes of two genotypes of Nicotiana tabacum L. Bot. Gaz 1991, 152, 13–22. [Google Scholar]
- Wang, K.C.; Ohnuma, S.-I. Isoprenyl diphosphate synthases. Biochim. Biophys. Acta 2000, 1529, 33–48. [Google Scholar]
- Dewick, P.M. The biosynthesis of C5-C25 terpenoid compounds. Nat. Prod. Rep 2002, 19, 181–222. [Google Scholar]
- Sallaud, C.; Rontein, D.; Onillon, S.; Jabès, F.; Duffé, P.; Giacalone, C.; Thoraval, S.; Escoffier, C.; Herbette, G.; Leonhardt, N.; et al. A novel pathway for sesquiterpene biosynthesis from Z,Z-farnesyl pyrophosphate in the wild tomato Solanum habrochaites. Plant Cell 2009, 21, 301–317. [Google Scholar]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J 2011, 66, 212–229. [Google Scholar]
- Falara, V.; Akhtar, T.A.; Nguyen, T.T.H.; Spyropoulou, E.A.; Bleeker, P.M.; Schauvinhold, I.; Matsuba, Y.; Bonini, M.E.; Schilmiller, A.L.; Last, R.L.; et al. The tomato terpene synthase gene family. Plant Physiol 2011, 157, 770–789. [Google Scholar]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar]
- Tholl, D.; Sohrabi, R.; Huh, J.-H.; Lee, S. The biochemistry of homoterpenes—common constituents of floral and herbivore-induced plant volatile bouqets. Phytochemistry 2011, 72, 1635–1646. [Google Scholar]
- Dicke, M.; Sabelis, M.W. How plants obtain predatory mites as bodyguards. Neth. J. Zool 1988, 38, 148–165. [Google Scholar]
- Schnee, C.; Köllner, T.G.; Held, M.; Turlings, T.C.J.; Gershenzon, J.; Degenhardt, J. The products of a single maize sesquiterpene synthase form a volatile defense signal that attracts natural enemies of maize herbivores. Proc. Natl. Acad. Sci. USA 2006, 103, 1129–1134. [Google Scholar]
- De Moraes, C.M.; Lewis, W.J.; Pare, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 393, 570–573. [Google Scholar]
- Van Poecke, R.M.P.; Posthumus, M.A.; Dicke, M. Herbivore-induced volatile production by Arabidopsis thaliana leads to attraction of the parasitoid Cotesia rubecula: Chemical, behavioral and gene-expression analysis. J. Chem. Ecol 2001, 27, 1911–1928. [Google Scholar]
- Dicke, M. Local and systemic production of volatile herbivore-induced terpenoids—their role in plant-carnivore mutualism. J. Plant Physiol 1994, 143, 465–472. [Google Scholar]
- Gibson, R.W.; Pickett, J.A. Wild potato repels aphids by release of aphid alarm pheromone. Nature 1983, 302, 608–609. [Google Scholar]
- Beale, M.H.; Birkett, M.A.; Bruce, T.J.A.; Chamberlain, K.; Field, L.M.; Huttley, A.K.; Martin, J.L.; Parker, R.; Phillips, A.L.; Pickett, J.A.; et al. Aphid alarm pheromone produced by transgenic plants affects aphid and parasitoid behavior. Proc. Natl. Acad. Sci. USA 2006, 103, 10509–10513. [Google Scholar]
- Bleeker, P.M.; Diergaarde, P.J.; Ament, K.; Guerra, J.; Weidner, M.; Schutz, S.; De Both, M.T.J.; Haring, M.A.; Schuurink, R.C. The role of specific tomato volatiles in tomato-whitefly interaction. Plant Physiol 2009, 151, 925–935. [Google Scholar]
- Bleeker, P.M.; Diergaarde, P.J.; Ament, K.; Schütz, S.; Johne, B.; Dijkink, J.; Hiemstra, H.; de Gelder, R.; de Both, M.T.J.; Sabelis, M.W.; et al. Tomato-produced 7-epizingiberene and R-curcumene act as repellents to whiteflies. Phytochemistry 2011, 72, 68–73. [Google Scholar]
- Carter, C.D.; Sacalis, J.N.; Gianfagna, T.J. Zingiberene and resistance to Colorado potato beetle in Lycopersicon hirsutum f. hirsutum. J. Agric. Food Chem 1989, 37, 206–210. [Google Scholar]
- Eigenbrode, S.D.; Trumble, J.T.; Millar, J.G.; White, K.K. Topical toxicity of tomato sesquiterpenes to the beet armyworm and the role of these compounds in resistance derived from an accession of Lycopersicon hirusutum ftypicum. J. Agric. Food Chem 1994, 42, 807–810. [Google Scholar]
- De Azevedo, S.M.; Faria, M.V.; Maluf, W.R.; de Oliveira, A.C.B.; De Freitas, J.A. Zingiberene-mediated resistance to the South American tomato pinworm derived from Lycopersicon hirsutum var. hirsutum. Euphytica 2003, 134, 347–351. [Google Scholar]
- Maluf, W.R.; Campos, G.A.; Cardoso, C.M. Relationships between trichome types and spider mite (Tetranychus evansi) repellence in tomatoes with respect to foliar zingiberene contents. Euphytica 2001, 121, 73–80. [Google Scholar]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar]
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and distribution of floral scent. Bot. Rev 2006, 72, 1–120. [Google Scholar]
- Iijima, Y.; Davidovich-Rikanati, R.; Fridman, E.; Gang, D.R.; Bar, E.; Lewinsohn, E.; Pichersky, E. The biochemical and molecular basis for the divergent patterns in the biosynthesis of terpenes and phenylpropenes in the peltate glands of three cultivars of basil. Plant Physiol 2004, 136, 3724–3736. [Google Scholar]
- Boatright, J.; Negre, F.; Chen, X.; Kish, C.M.; Wood, B.; Peel, G.; Orlova, I.; Gang, D.; Rhodes, D.; Dudareva, N. Understanding in vivo benzenoids metabolism in petunia petal tissue. Plant Physiol 2004, 135, 1993–2011. [Google Scholar]
- Van Moerkercke, A.; Schauvinhold, I.; Pichersky, E.; Haring, M.A.; Schuurink, R.C. A plant thiolase involved in benzoic acid biosynthesis and volatile benzenoid production. Plant J 2009, 60, 292–302. [Google Scholar]
- Yang, D.S.; Sumner, L.W. Metabolic profiling of trichomes from potato leafhopper susceptible and resistant alfalfa lines. Available online: http://www.planttrichome.org accessed on 1 November 2012.
- Kapteyn, J.; Qualley, A.; Xie, Z.; Fridman, E.; Dudareva, N.; Gang, D.R. Evolution of cinnamate/p-coumarate carboxylmethyltransferases and their role in the biosynthesis of methylcinnamate. Plant Cell 2007, 19, 3212–3229. [Google Scholar]
- Koeduka, T.; Fridman, E.; Gang, D.R.; Vassão, D.G.; Jackson, B.L.; Kish, C.M.; Orlova, I.; Spassova, S.M.; Lewis, N.G.; Noel, J.P.; et al. Eugenol and isoeugenol, characteristic aromatic constituents of spices, are biosynthesized via reduction of coniferyl acohol ester. Proc. Natl. Acad. Sci. USA 2006, 103, 10128–10133. [Google Scholar]
- Gang, D.R.; Lavid, N.; Zubieta, C.; Chen, F.; Beuerle, T.; Lewinsohn, E.; Noel, J.P.; Pichersky, E. Characterization of phenylpropene O-methyltransferases from sweet basil: Facile change of substrate specificity and convergent evolution within a plant O-methyltransferase family. Plant Cell 2002, 14, 505–519. [Google Scholar]
- Tan, K.H.; Nishida, R.; Toong, Y.C. Floral synomone of a wild orchid, Bulbophyllum cheiri, lures Bactrocera fruit flies for pollination. J. Chem. Ecol 2002, 28, 1161–1172. [Google Scholar]
- Obeng-Ofori, D.; Reichmuth, C. Bioactivity of eugenol, a major component of essential oil of Ocimum suave (Wild.) against four species of stored-product Coleoptera. Int. J. Pest Manag 1997, 43, 89–94. [Google Scholar]
- Sangwan, N.; Verman, B.; Verma, K.; Dhindsa, K. Nematicidal activity of some essential plant oils. Pest Sci 1990, 28, 331–335. [Google Scholar]
- Adams, S.; Weidenborner, M. Mycelial deformations of Cladosporium herbarum due to the application of eugenol or carvacrol. J. Essential Oil Res 1996, 8, 535–540. [Google Scholar]
- Ferrer, J.; Austin, M.B.; Stewart, C., Jr; Noel, J.P. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar]
- Tattini, M.; Gravano, E.; Pinelli, P.; Mulinacci, N.; Romani, A. Flavonoids accumulate in leaves and glandular trichomes of Phillyrea latifolia exposed to excess solar radiation. New Phytol 2000, 148, 69–77. [Google Scholar]
- Schmidt, A.; Li, C.; Shi, F.; Jones, A.D.; Pichersky, E. Polymethylated myricetin in trichomes of the wild tomato species Solanum habrochaites and characterization of trichome-specific 3′/5′- and 7/4′-myricetin O-methyltransferases. Plant Physiol 2011, 155, 1999–2009. [Google Scholar]
- Kang, J.; Liu, G.; Shi, F.; Jones, A.D.; Beaudry, R.M.; Howe, G.A. The tomato odorless-2 mutant is defective in trichome-based production of diverse specialized metabolites and broad-spectrum resistance to insect herbivores. Plant Physiol 2010, 154, 262–272. [Google Scholar]
- Duffey, S.S.; Isman, M.B. Inhibition of insect larval growth by phenolics in glandular trichomes of tomato leaves. Experientia 1981, 37, 574–576. [Google Scholar]
- Ben-Israel, I.; Yu, G.; Austin, M.B.; Bhuiyan, N.; Auldridge, M.; Nguyen, T.; Schauvinhold, I.; Noel, J.P.; Pichersky, E.; Fridman, E. Multiple biochemical and morphological factors underlie the production of methylketones in tomato. Plant Physiol 2009, 151, 1952–1964. [Google Scholar]
- Yu, G.; Nguyen, T.T.H.; Guo, Y.; Schauvinhold, I.; Auldridge, M.E.; Bhuiyan, N.; Ben-Israel, I.; Iijima, Y.; Fridman, E.; Noel, J.P.; et al. Enzymatic functions of wild tomato methylketone synthases 1 and 2. Plant Physiol 2010, 154, 67–77. [Google Scholar]
- Williams, W.G.; Kennedy, G.G.; Yamamoto, R.T.; Thacker, J.D.; Bordner, J. 2-Tridecanone: A naturally occurring insecticide from the wild tomato Lycopersicon hirsutum f. glabratum. Science 1980, 207, 888–889. [Google Scholar]
- Antonious, G.F. Production and quantification of methyl ketones in wild tomato accessions. J. Environ. Sci. Health B 2001, 36, 835–848. [Google Scholar]
- Dimock, M.B.; Kennedy, G.G. The role of glandular trichomes in the resistance of Lycopersicon hirsutum f. glabratum to Heliothis zea. Entomol. Expl. Appl 1983, 33, 263–268. [Google Scholar]
- Chatzivasileiadis, E.A.; Sabelis, M.W. Toxicity of methyl ketones from tomato trichomes to Tetranychus urticae Koch. Exp. Appl. Acarol 1997, 21, 473–484. [Google Scholar]
- Musetti, L.; Neal, J.J. Toxicological effect of Lycopersicon hirsutum f. glabratum and behavioral response of Macrosiphum euphorbia. J. Chem. Ecol 1997, 23, 1321–1332. [Google Scholar]
- Farrar, R.R.; Kennedy, G.G. 2-Undecanone, a constituent of the glandular trichomes of Lycopersicon hirsutum f. glabratum: Effects on Heliothis zea and Manduca sexta growth and survival. Entomol. Exp. Appl 1987, 43, 17–23. [Google Scholar]
- Slocombe, S.P.; Schauvinhold, I.; McQuinn, R.P.; Besser, K.; Welsby, N.A.; Harper, A.; Aziz, N.; Li, Y.; Larson, T.R.; Giovannoni, J.; et al. Transcriptomic and reverse genetic analyses of branched-chain fatty acid and acyl sugar production in Solanum pennellii and Nicotiana benthamiana. Plant Physiol 2008, 148, 1830–1846. [Google Scholar]
- Chortyk, O.T.; Kays, S.J.; Teng, Q. Characterization of insecticidal sugar esters of Petunia. J. Agric. Food Chem 1997, 45, 270–275. [Google Scholar]
- Fobes, J.F.; Mudd, J.B.; Marsden, M.P.F. Epicuticular lipid accumulation on the leaves of Lycopersicon pennellii (Corr.) D’Arcy and Lycopersicon esculentum Mill. Plant Physiol 1985, 77, 567–570. [Google Scholar]
- Schilmiller, A.L.; Charbonneau, A.L.; Last, R.L. Identification of a BAHD acetyltransferase that produces protective acyl sugars in tomato trichomes. Proc. Natl. Acad. Sci. USA 2012, 109, 16377–16382. [Google Scholar]
- Weinhold, A.; Baldwin, I.T. Trichome-derived O-acyl sugars are a first meal for caterpillars that tags them for predation. Proc. Natl. Acad. Sci. USA 2011, 108, 7855–7859. [Google Scholar]
- Burke, B.A.; Goldsby, G.; Mudd, J.B. Polar epicuticular lipids of Lycopersicon pennellii. Phytochemistry 1987, 26, 2567–2571. [Google Scholar]
- Walters, D.S.; Steffens, J.C. Branched chain amino acid metabolism in the biosynthesis of Lycopersicon pennellii glucose esters. Plant Physiol 1990, 93, 1544–1551. [Google Scholar]
- Li, A.X.; Eannetta, N.; Ghangas, G.S.; Steffens, J.C. Glucose polyester biosynthesis. Purification and characterization of a glucose acyltransferase. Plant Physiol 1999, 121, 453–460. [Google Scholar]
- Ghangas, G.S.; Steffens, J.C. UDP glucose:fatty acid transglucosylation and transacylation in triacylglucose biosynthesis. Proc. Natl. Acad. Sci. USA 1993, 90, 9911–9915. [Google Scholar]
- Li, A.X.; Steffens, J.C. An acyltransferase catalyzing the formation of diacylglucose is a serine carboxipeptidase-like protein. Proc. Natl. Acad. Sci. USA 2000, 97, 6902–6907. [Google Scholar]
- Goffreda, J.C.; Mutschler, M.A.; Avé, D.A.; Tingey, W.M.; Steffens, J.C. Aphid deterrence by glucose esters in glandular trichome exudate of the wild tomato, Lycopersicon pennellii. J. Chem. Ecol 1989, 15, 2135–2147. [Google Scholar]
- Saeidi, Z.; Mallik, B.; Kulkarni, R.S. Inheritance of glandular trichomes and two-spotted spider mite resistance in cross Lycopersicon esculentum “Nandi” and L. pennellii “LA2963”. Euphytica 2007, 154, 231–238. [Google Scholar]
- Alba, J.M.; Montserrat, M.; Fernández-Muñoz, R. Resistance to the two-spotted spider mite (Tetranychus urticae) by acylsucroses of wild tomato (Solanum pimpinellifolium) trichomes studied in a recombinant inbred line population. Exp. Appl. Acarol 2009, 47, 35–47. [Google Scholar]
- Rodríguez-López, M.J.; Garzo, E.; Bonani, J.P.; Fereres, A.; Fernández-Muñoz, R.; Moriones, E. Whitefly resistance traits derived from the wild tomato Solanum pimpinellifolium affect the preference and feeding behavior of Bemisia tabaci and reduce the spread of tomato yellow leaf curl virus. Phytopathology 2011, 101, 1191–1201. [Google Scholar]
- Juvik, J.A.; Shapiro, J.A.; Young, T.E.; Mutschler, M.A. Acylglucoses from wild tomatoes alter behavior and reduce growth and survival of Helicoverpa zea and Spodoptera exigua (Lepidoptera: Noctuidae). J. Econ. Entomol 1994, 87, 482–492. [Google Scholar]
- Hawthorne, D.J.; Shapiro, J.A.; Tingey, W.M.; Mutschler, M.A. Trichome-borne and artificially applied acylsugars of wild tomato deter feeding and oviposition of the leafminer Liriomyza trifolii. Entomol. Exp. Appl 1992, 65, 65–73. [Google Scholar]
- Liu, J.; Xia, K.F.; Zhu, J.C.; Deng, Y.G.; Huang, X.L.; Hu, B.L.; Xu, X.; Xu, Z.F. The nightshade proteinase inhibitor IIb gene is constitutively expressed in glandular trichomes. Plant Cell Physiol 2006, 47, 1274–1284. [Google Scholar]
- Yu, H.; Kowalski, S.P.; Steffens, J.C. Comparison of polyphenol oxidase expression in glandular trichomes of Solanum and Lycopersicon species. Plant Physiol 1992, 100, 1885–1890. [Google Scholar]
- Steppuhn, A.; Baldwin, I.T. Resistance management in a native plant: Nicotine prevents herbivores from compensating for plant protease inhibitors. Ecol. Lett 2007, 10, 499–511. [Google Scholar]
- Hartl, M.; Giri, A.P.; Kaur, H.; Baldwin, I.T. Serine protease inhibitors specifically defend Solanum nigrum against generalist herbivores but do not influence growth and development. Plant Cell 2010, 22, 4158–4175. [Google Scholar]
- Vaughn, K.C.; Lax, A.R.; Duke, S.O. Polyphenol oxidase: The chloroplast oxidase with no established function. Physiol. Plant 1988, 72, 659–665. [Google Scholar]
- Kowalski, S.P.; Eannetta, N.T.; Hirzel, A.T.; Steffens, J.C. Purification and characterization of polyphenol oxidase from glandular trichomes of Solanum berthaultii. Plant Physiol 1992, 100, 677–684. [Google Scholar]
- Thipyapong, P.; Joel, D.M.; Steffens, J.C. Differential expression and turnover of the tomato polyphenol oxidase gene familiy during vegetative and reproductive development. Plant Physiol 1997, 113, 707–718. [Google Scholar]
- Felton, G.W.; Donato, K.; del Vecchio, R.J.; Duffey, S.S. Activation of plant foliar oxidases by insect feeding reduces nutritive quality of foliage for noctuid herbivores. J. Chem. Ecol 1989, 15, 2667–2694. [Google Scholar]
- Felton, G.W.; Donato, K.K.; Broadway, R.M.; Duffey, S.S. Impact of oxidized plant phenolics on the nutrional quality of dietary protein to a noctuid herbivore, Spodoptera exigua. J. Insect Physiol 1992, 38, 277–285. [Google Scholar]
- Li, L.; Steffens, J.C. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta 2002, 215, 239–247. [Google Scholar]
- Thipyapong, P.; Hunt, M.D.; Steffens, J.C. Antisense downregulation of polyphenol oxidase results in enhanced disease susceptibility. Planta 2004, 220, 105–117. [Google Scholar]
- Richter, C.; Dirks, M.E.; Gronover, C.S.; Prűfer, D.; Moerschbacher, B.M. Silencing and heterologous expression of ppo-2 indicate a specific function of a single polyphenol oxidase isoform in resistance of Dandelion (Taraxacum officinale) against Pseudomonas syringae pv. tomato. Mol. Plant Microbe Int 2012, 25, 200–210. [Google Scholar]
- Shepherd, R.W.; Wagner, G.J. Phylloplane proteins: Emerging defenses at the aerial frontline. Trends Plant Sci 2007, 12, 51–56. [Google Scholar]
- Tissier, A. Trichome specific expression: Promoters and their applications. In Transgenic plants – Advances and limitations; Çiftçi, Y.O., Ed.; InTech: Rijeka, Croatia, 2012; pp. 353–378. [Google Scholar]
- Shepherd, R.W.; Wagner, G.J. Fungi and leaf surfaces. In Biocomplexity of plant-fungal interactions; Southworth, D., Ed.; John Wiley & Sons: Chichester, West Sussex, UK, 2012; p. 131. [Google Scholar]
- Harada, E.; Kim, J.; Meyer, A.J.; Hell, R.; Clemens, S.; Choi, Y. Expression profiling of tobacco leaf trichomes identifies genes for biotic and abiotic stress. Plant Cell Physiol 2010, 51, 1627–1637. [Google Scholar]
- Cui, H.; Zhang, S.; Yang, H.; Ji, H.; Wang, X. Gene expression profile analysis of tobacco leaf trichomes. BMC Plant Biol 2011, 11, 76–86. [Google Scholar]
- Keene, C.K.; Wagner, G.J. Direct demonstration of duvatrienediol biosynthesis in glandular heads of tobacco trichomes. Plant Physiol 1985, 79, 1026–1032. [Google Scholar]
- Kandra, L.; Wagner, G.J. Studies of the site and mode of biosynthesis of tobacco trichome exudate components. Arch. Biochem. Biophys 1988, 265, 425–432. [Google Scholar]
- Lange, B.M.; Wildung, M.R.; Stauber, E.J.; Sanchez, C.; Pouchnick, D.; Croteau, R. Probing essential oil biosynthesis and secretion by functional evaluation of expressed sequence tags from mint glandular trichomes. Proc. Natl. Acad. Sci. USA 2000, 97, 2934–2939. [Google Scholar]
- Wagner, G.J. Secreting glandular trichomes: More than just hairs. Plant Physiol 1991, 96, 675–679. [Google Scholar]
- Franceschi, V.R.; Giaquinta, R.T. Glandular trichomes of soybean leaves: Cytological differentiation from initiation through senescence. Bot. Gaz 1983, 144, 175–184. [Google Scholar]
- Iijima, Y.; Gang, D.R.; Fridman, E.; Lewinsohn, E.; Pichersky, E. Characterization of geraniol synthase from the peltate glands of sweet basil. Plant Physiol 2004, 134, 370–379. [Google Scholar]
- Schilmiller, A.L.; Miner, D.P.; Larson, M.; McDowell, E.; Gang, D.R.; Wilkerson, C.; Last, R.L. Studies of a biochemical factory: Tomato trichome deep expressed sequence tag sequencing and proteomics. Plant Physiol 2010, 153, 1212–1223. [Google Scholar]
- Bleeker, P.M.; Spyropoulou, E.A.; Diergaarde, P.J.; Volpin, H.; De Both, M.T.J.; Zerbe, P.; Bohlmann, J.; Falara, V.; Matsuba, Y.; Pichersky, E.; et al. RNA-seq discovery, functional characterization, and comparison of sesquiterpene synthases from Solanum lycopersicum and Solanum habrochaites trichomes. Plant Mol. Biol 2011, 77, 323–336. [Google Scholar]
- Nagel, J.; Culley, L.K.; Lu, Y.; Liu, E.; Matthews, P.D.; Stevens, J.F.; Page, J.E. EST analysis of hop glandular trichomes identifies an O-methyltransferase that catalyzes the biosynthesis of xanthohumol. Plant Cell 2008, 20, 186–200. [Google Scholar]
- Choi, Y.; Lim, S.; Kim, H.; Han, J.; Lee, M.; Yang, Y.; Kim, J.; Kim, Y. Tobacco NtLTP1, a glandular-specific lipid transfer protein, is required for lipid secretion from glandular secretions. Plant J 2012, 70, 480–491. [Google Scholar]
- Xie, Z.; Kapteyn, J.; Gang, D.R. A systems biology investigation of the MEP/terpenoid and shikimate/phenylpropanoid pathways points to multiple levels of metabolic control in sweet basil glandular trichomes. Plant J 2008, 54, 349–361. [Google Scholar]
- Besser, K.; Harper, A.; Welsby, N.; Schauvinhold, N.; Slocombe, S.; Li, Y.; Dixon, R.A.; Broun, P. Divergent regulation of terpenoid metabolism in the trichomes of wild and cultivated tomato species. Plant Physiol 2009, 149, 499–514. [Google Scholar]
- Wang, S.; Wang, J.; Yu, N.; Li, C.; Luo, B.; Gou, J.; Wang, L.; Chen, X. Control of plant trichome development by a cotton fiber MYB gene. Plant Cell 2004, 16, 2323–2334. [Google Scholar]
- Wang, E.; Wang, R.; DeParasis, J.; Loughrin, J.H.; Gan, S.; Wagner, G.J. Suppression of a P450 hydroxylase gene in plant trichome glands enhances natural-product-based aphid resistance. Nature Biotech 2001, 19, 371–374. [Google Scholar]
- Wang, E.; Gan, S.; Wagner, G.J. Isolation and characterization of the CYP71D16 trichome-specific promoter of Nicotiana tabacum L. J. Exp. Bot 2002, 53, 1891–1897. [Google Scholar]
- Ennajdaoui, H.; Vachon, G.; Giacalone, C.; Besse, I.; Sallaud, C.; Herzog, M.; Tissier, A. Trichome specific expression of the tobacco (Nicotiana sylvestris) cembratrien-ol synthase genes is controlled by both activating and repressing cis-regions. Plant Mol. Biol 2010, 73, 673–685. [Google Scholar]
- Rontein, D.; Onillon, S.; Herbette, G.; Lesot, A.; Werck-Reichhart, D.; Sallaud, C.; Tissier, A. CYP725A4 from yew catalyzes complex structural rearrangement of taxa-4(5),11(12)-diene into the cyclic ether 5(12)-oxa-3(11)-cyclotaxane. J. Biol. Chem 2008, 283, 6067–6075. [Google Scholar]
- Anandan, A.; Gatehouse, L.N.; Marshall, R.K.; Murray, C.; Christeller, J.T. Two highly homologous promoters of a squash aspartic protease inhibitor (SQAPI) multigene family exhibit differential expression in transgenic tobacco phloem and trichome cells. Plant Mol. Biol. Rep 2009, 27, 355–364. [Google Scholar]
- Liu, H.; Creech, R.G.; Jenkins, J.N.; Ma, D. Cloning and promoter analysis of the cotton lipid transfer protein gene Ltp3. Biochim. Biophys. Acta 2000, 1487, 106–111. [Google Scholar]
- Bleeker, P.M.; Mirabella, R.; Diergaarde, P.J.; Van Doorn, A.; Tissier, A.; Kant, M.R.; Prins, M.; de Vos, M.; Haring, M.A.; Schuurink, R.C. Improved herbivore resistance in cultivated tomato with the sesquiterpene biosynthetic pathway from a wild relative. Proc. Natl. Acad. Sci. USA 2012. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
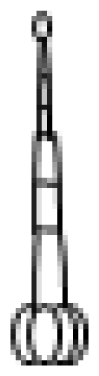{kind=link}
{kind=link}
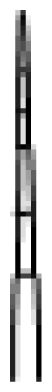{kind=link}
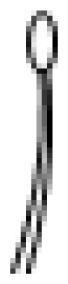{kind=link}
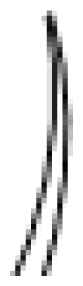{kind=link}
{kind=link}
{kind=link}
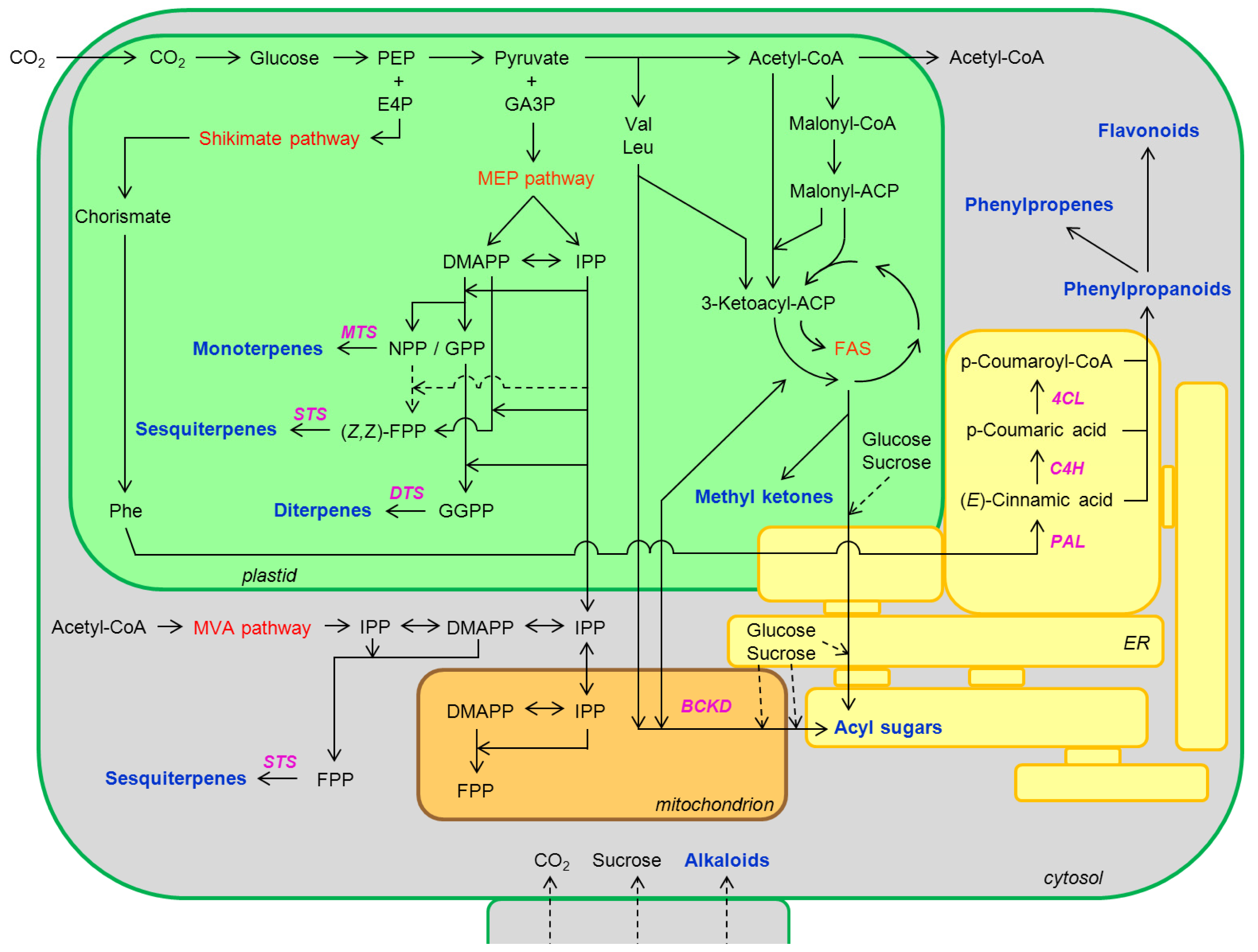
| Type | Description |
|---|---|
I
 | Thin glandular trichomes consisting of 6–10 cells and 2–3 mm long. Globular and multicellular base with a small and round glandular cell in the trichome tip. |
II
 | Similar to trichome I but non-glandular and shorter (0.2–1.0 mm). Globular and multicellular base. |
III
 | Thin non-glandular trichome consisting of 4–8 cells and 0.4–1.0 mm long with a unicellular and flat base. External walls lack intercellular sections. |
IV
 | Similar to trichome I but shorter (0.2–0.4 mm) and with a glandular cell in the tip. Trichome base is unicellular and flat. |
V
 | Very similar to type IV with respect to height and thickness but non-glandular. |
VI
 | Thick and short glandular trichomes composed of two stalk cells and a head made up of 4 secretory cells. |
VII
 | Very small glandular trichomes (0.05 mm) with a head consisting of 4–8 cells. |
VIII
 | Non-glandular trichome composed of one basal and thick cell with a leaning cell in the tip. |
| Species | I | II | III | IV | V | VI | VII | VIII |
|---|---|---|---|---|---|---|---|---|
| S. habrochaites | + | + | + | + | + | |||
| S. lycopersicum | + | + | + | + | + | + | ||
| S. pennellii | + | + | ||||||
| S. cheesmaniae, S. galapagense | + | |||||||
| S. pimpinellifolium | + | + b | + | + | ||||
| S. peruvianum, S. arcanum, S. corneliomuelleri, S. huylasense | + | + a | + | + | + | |||
| S. chilense | + | + | + | |||||
| S. chmielewski | + | + | ||||||
| S. neorickii | + | + |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Glas, J.J.; Schimmel, B.C.J.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant Glandular Trichomes as Targets for Breeding or Engineering of Resistance to Herbivores. Int. J. Mol. Sci. 2012, 13, 17077-17103. https://doi.org/10.3390/ijms131217077
Glas JJ, Schimmel BCJ, Alba JM, Escobar-Bravo R, Schuurink RC, Kant MR. Plant Glandular Trichomes as Targets for Breeding or Engineering of Resistance to Herbivores. International Journal of Molecular Sciences. 2012; 13(12):17077-17103. https://doi.org/10.3390/ijms131217077
Chicago/Turabian StyleGlas, Joris J., Bernardus C. J. Schimmel, Juan M. Alba, Rocío Escobar-Bravo, Robert C. Schuurink, and Merijn R. Kant. 2012. "Plant Glandular Trichomes as Targets for Breeding or Engineering of Resistance to Herbivores" International Journal of Molecular Sciences 13, no. 12: 17077-17103. https://doi.org/10.3390/ijms131217077
APA StyleGlas, J. J., Schimmel, B. C. J., Alba, J. M., Escobar-Bravo, R., Schuurink, R. C., & Kant, M. R. (2012). Plant Glandular Trichomes as Targets for Breeding or Engineering of Resistance to Herbivores. International Journal of Molecular Sciences, 13(12), 17077-17103. https://doi.org/10.3390/ijms131217077