Biological Significance of Calbindin-D9k within Duodenal Epithelium



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
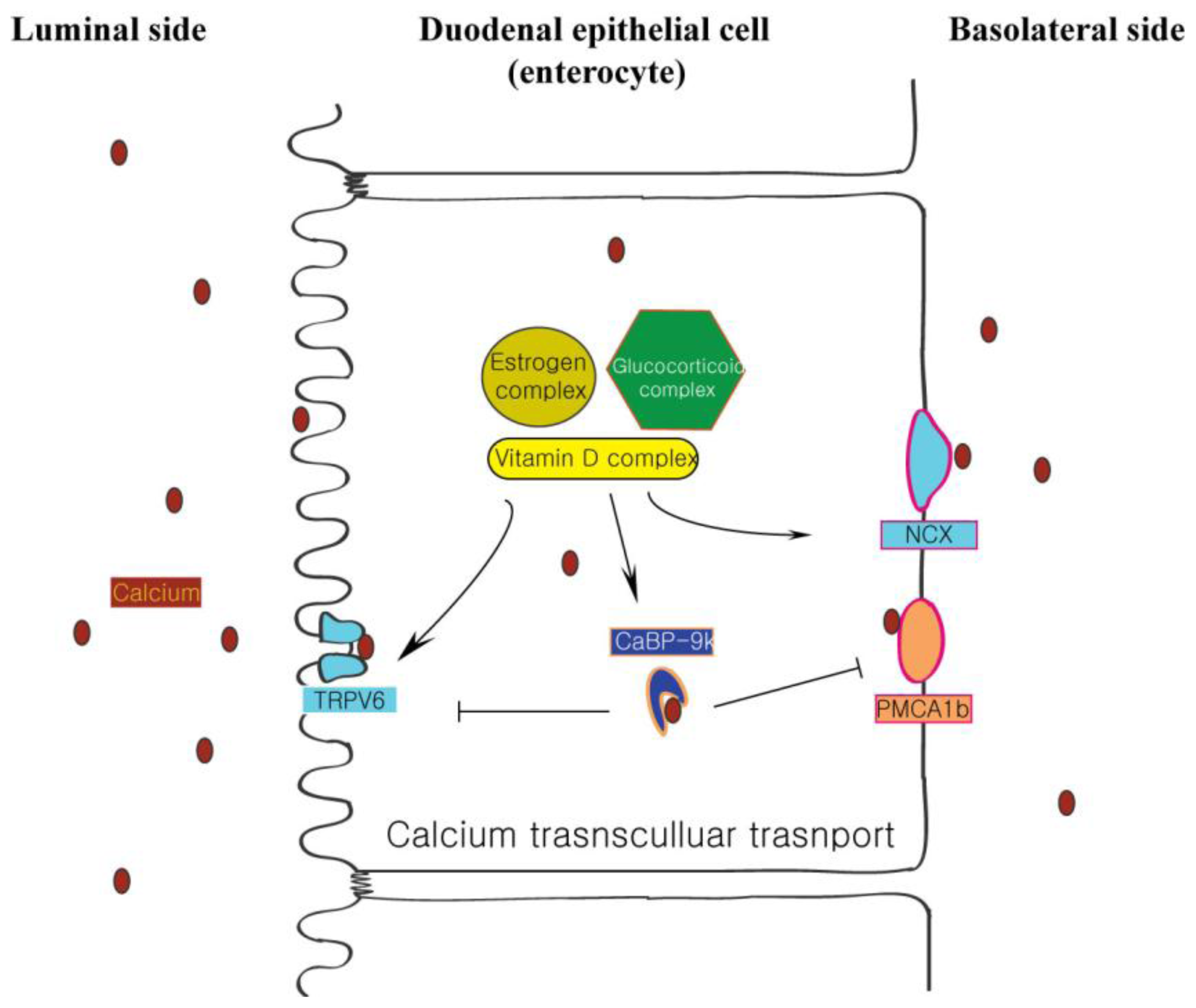
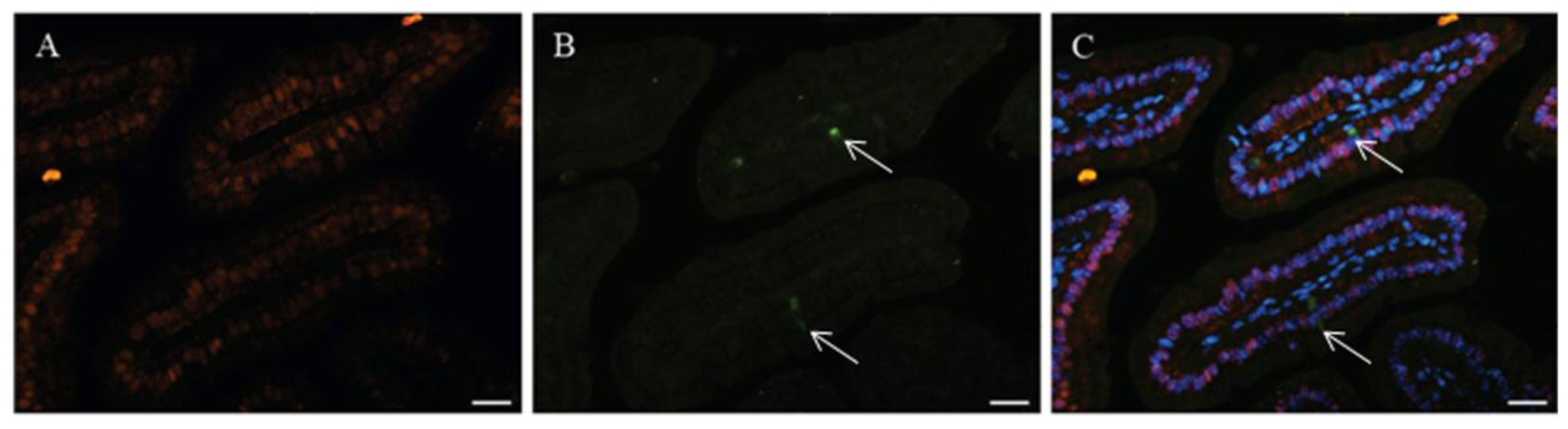
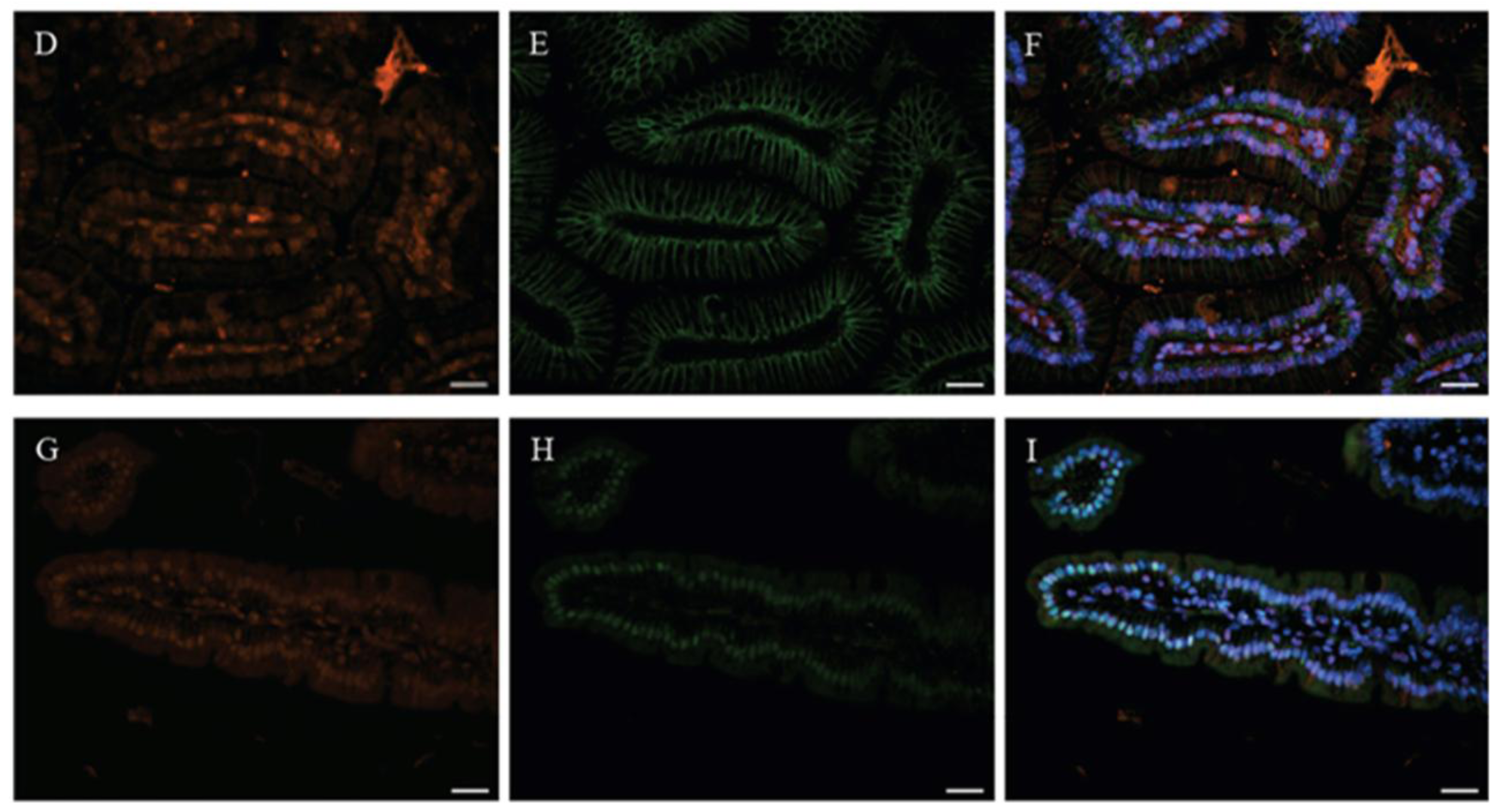
2. Vitamin D-Dependent CaBP-9k Expression in the Small Intestine
3. Other Physiological Factors that Affect CaBP-9k Expression
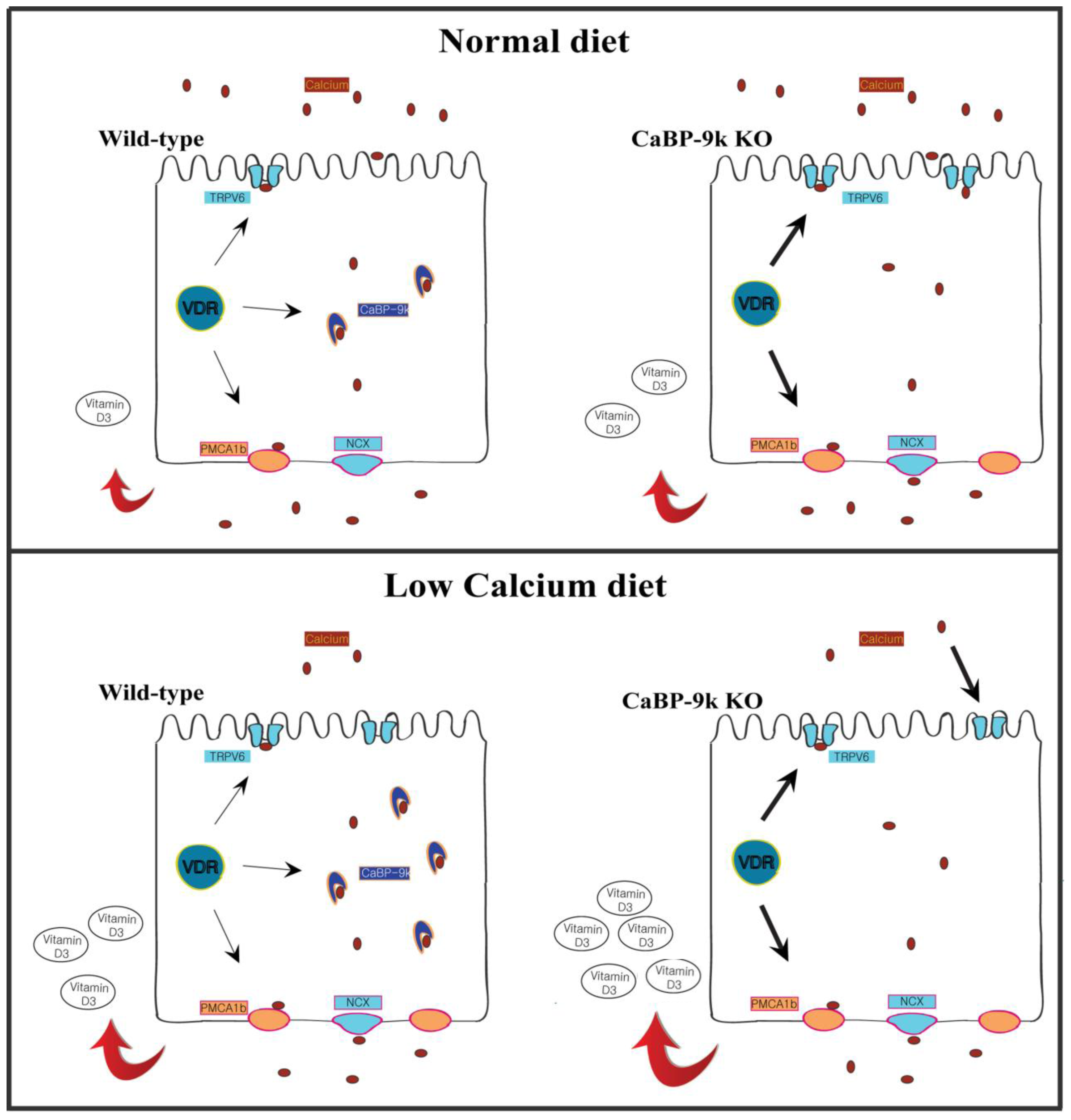
4. The Influence of Intracellular CaBP-9k in Expression of Other Calcium-Associated Proteins
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Wasserman, R.H.; Taylor, A.N. Vitamin D3-induced calcium-binding protein in chick intestinal mucosa. Science 1966, 152, 791–793. [Google Scholar]
- Ingersoll, R.J.; Wasserman, R.H. Vitamin D3-induced calcium-binding protein. Binding characteristics, conformational effects, and other properties. J. Biol. Chem 1971, 246, 2808–2814. [Google Scholar]
- Schwaller, B. Cytosolic Ca2+ buffers. Cold Spring Harb. Perspect. Biol 2010, 2, a004051. [Google Scholar]
- Hoenderop, J.G.; Nilius, B.; Bindels, R.J. Calcium absorption across epithelia. Physiol. Rev 2005, 85, 373–422. [Google Scholar]
- Jensen, T.P.; Buckby, L.E.; Empson, R.M. Expression of plasma membrane Ca2+ ATPase family members and associated synaptic proteins in acute and cultured organotypic hippocampal slices from rat. Brain Res. Dev. Brain Res 2004, 152, 129–136. [Google Scholar]
- DiPolo, R.; Beauge, L. Sodium/calcium exchanger: Influence of metabolic regulation on ion carrier interactions. Physiol. Rev 2006, 86, 155–203. [Google Scholar]
- Yu, S.P.; Choi, D.W. Na+–Ca2+ exchange currents in cortical neurons: Concomitant forward and reverse operation and effect of glutamate. Eur. J. Neurosci 1997, 9, 1273–1281. [Google Scholar]
- Bronner, F.; Pansu, D. Nutritional aspects of calcium absorption. J. Nutr 1999, 129, 9–12. [Google Scholar]
- Bronner, F.; Slepchenko, B.; Wood, R.J.; Pansu, D. The role of passive transport in calcium absorption. J. Nutr 2003, 133, 1426–1427. [Google Scholar]
- Jeung, E.B.; Krisinger, J.; Dann, J.L.; Leung, P.C. Cloning of the porcine Calbindin-D9k complementary deoxyribonucleic acid by anchored polymerase chain reaction technique. Biol. Reprod 1992, 47, 503–508. [Google Scholar]
- Crosnier, C.; Stamataki, D.; Lewis, J. Organizing cell renewal in the intestine: Stem cells, signals and combinatorial control. Nat. Rev. Genet 2006, 7, 349–359. [Google Scholar]
- Van Cromphaut, S.J.; Rummens, K.; Stockmans, I.; van Herck, E.; Dijcks, F.A.; Ederveen, A.G.; Carmeliet, P.; Verhaeghe, J.; Bouillon, R.; Carmeliet, G. Intestinal calcium transporter genes are upregulated by estrogens and the reproductive cycle through vitamin D receptor-independent mechanisms. J. Bone Miner. Res 2003, 18, 1725–1736. [Google Scholar]
- Kimmel-Jehan, C.; Jehan, F.; DeLuca, H.F. Salt concentration determines 1,25-dihydroxyvitamin D3 dependency of vitamin D receptor-retinoid X receptor—Vitamin D-responsive element complex formation. Arch. Biochem. Biophys 1997, 341, 75–80. [Google Scholar]
- Dardenne, O.; Prud’homme, J.; Arabian, A.; Glorieux, F.H.; St-Arnaud, R. Targeted inactivation of the 25-hydroxyvitamin D(3)-1(alpha)-hydroxylase gene (CYP27B1) creates an animal model of pseudovitamin D-deficiency rickets. Endocrinology 2001, 142, 3135–3141. [Google Scholar]
- Van Cromphaut, S.J.; Dewerchin, M.; Hoenderop, J.G.; Stockmans, I.; van Herck, E.; Kato, S.; Bindels, R.J.; Collen, D.; Carmeliet, P.; Bouillon, R.; et al. Duodenal calcium absorption in vitamin D receptor-knockout mice: Functional and molecular aspects. Proc. Natl. Acad. Sci. USA 2001, 98, 13324–13329. [Google Scholar]
- Wasserman, R.H.; Fullmer, C.S. Vitamin D and intestinal calcium transport: Facts, speculations and hypotheses. J. Nutr 1995, 125, 1971S–1979S. [Google Scholar]
- Colnot, S.; Ovejero, C.; Romagnolo, B.; Porteu, A.; Lacourte, P.; Thomasset, M.; Perret, C. Transgenic analysis of the response of the rat calbindin-D9k gene to vitamin D. Endocrinology 2000, 141, 2301–2308. [Google Scholar]
- Balesaria, S.; Sangha, S.; Walters, J.R. Human duodenum responses to vitamin D metabolites of TRPV6 and other genes involved in calcium absorption. Am. J. Physiol. Gastrointest Liver Physiol 2009, 297, 1193–1197. [Google Scholar]
- Kwiecinksi, G.G.; Petrie, G.I.; DeLuca, H.F. 1,25-Dihydroxyvitamin D3 restores fertility of vitamin D-deficient female rats. Am. J. Physiol 1989, 256, 483–487. [Google Scholar]
- Hoenderop, J.G.; Dardenne, O.; van Abel, M.; van der Kemp, A.W.; van Os, C.H.; St-Arnaud, R.; Bindels, R.J. Modulation of renal Ca2+ transport protein genes by dietary Ca2+ and 1,25-dihydroxyvitamin D3 in 25-hydroxyvitamin D3-1alpha-hydroxylase knockout mice. FASEB J 2002, 16, 1398–1406. [Google Scholar]
- Darwish, H.M.; DeLuca, H.F. Identification of a 1,25-dihydroxyvitamin D3-response element in the 5′-flanking region of the rat calbindin D9k gene. Proc. Natl. Acad. Sci. USA 1992, 89, 603–607. [Google Scholar]
- Li, Y.C.; Pirro, A.E.; Demay, M.B. Analysis of vitamin D-dependent calcium-binding protein messenger ribonucleic acid expression in mice lacking the vitamin D receptor. Endocrinology 1998, 139, 847–851. [Google Scholar]
- Uhland, A.M.; Kwiecinski, G.G.; DeLuca, H.F. Normalization of serum calcium restores fertility in vitamin D-deficient male rats. J. Nutr 1992, 122, 1338–1344. [Google Scholar]
- L’Horset, F.; Perret, C.; Brehier, A.; Thomasset, M. 17 beta-estradiol stimulates the calbindin-D9k (CaBP9k) gene expression at the transcriptional and posttranscriptional levels in the rat uterus. Endocrinology 1990, 127, 2891–2897. [Google Scholar]
- Lee, G.S.; Choi, K.C.; Park, S.M.; An, B.S.; Cho, M.C.; Jeung, E.B. Expression of human Calbindin-D(9k) correlated with age, vitamin D receptor and blood calcium level in the gastrointestinal tissues. Clin. BioChem 2003, 36, 255–261. [Google Scholar]
- Armbrecht, H.J.; Zenser, T.V.; Davis, B.B. Effect of vitamin D metabolites on intestinal calcium absorption and calcium-binding protein in young and adult rats. Endocrinology 1980, 106, 469–475. [Google Scholar]
- Lee, G.S.; Lee, K.Y.; Choi, K.C.; Ryu, Y.H.; Paik, S.G.; Oh, G.T.; Jeung, E.B. Phenotype of a calbindin-D9k gene knockout is compensated for by the induction of other calcium transporter genes in a mouse model. J. Bone Miner. Res 2007, 22, 1968–1978. [Google Scholar]
- Kimberg, D.V.; Baerg, R.D.; Gershon, E.; Graudusius, R.T. Effect of cortisone treatment on the active transport of calcium by the small intestine. J. Clin. Invest 1971, 50, 1309–1321. [Google Scholar]
- Shultz, T.D.; Bollman, S.; Kumar, R. Decreased intestinal calcium absorption in vivo and normal brush border membrane vesicle calcium uptake in cortisol-treated chickens: Evidence for dissociation of calcium absorption from brush border vesicle uptake. Proc. Natl. Acad. Sci. USA 1982, 79, 3542–3546. [Google Scholar]
- Lee, D.B. Unanticipated stimulatory action of glucocorticoids on epithelial calcium absorption. Effect of dexamethasone on rat distal colon. J. Clin. Invest 1983, 71, 322–328. [Google Scholar]
- Feher, J.J.; Wasserman, R.H. Intestinal calcium-binding protein and calcium absorption in cortisol-treated chicks: effects of vitamin D3 and 1,25-dihydroxyvitamin D3. Endocrinology 1979, 104, 547–551. [Google Scholar]
- Bastl, C.P.; Schulman, G.; Cragoe, E.J., Jr. Low-dose glucocorticoids stimulate electroneutral NaCl absorption in rat colon. Am. J. Physiol 1989, 257, 1027–1038. [Google Scholar]
- Sheppard, K.E.; Li, K.X.; Autelitano, D.J. Corticosteroid receptors and 11beta-hydroxysteroid dehydrogenase isoforms in rat intestinal epithelia. Am. J. Physiol 1999, 277, 541–547. [Google Scholar]
- Lee, G.S.; Choi, K.C.; Jeung, E.B. Glucocorticoids differentially regulate expression of duodenal and renal calbindin-D9k through glucocorticoid receptor-mediated pathway in mouse model. Am. J. Physiol. Endocrinol. Metab 2006, 290, 299–307. [Google Scholar]
- Lambers, T.T.; Mahieu, F.; Oancea, E.; Hoofd, L.; de Lange, F.; Mensenkamp, A.R.; Voets, T.; Nilius, B.; Clapham, D.E.; Hoenderop, J.G.; et al. Calbindin-D28K dynamically controls TRPV5-mediated Ca2+ transport. EMBO J 2006, 25, 2978–2988. [Google Scholar]
- Lee, G.S.; Choi, K.C.; Kim, H.J.; Jeung, E.B. Effect of genistein as a selective estrogen receptor beta agonist on the expression of Calbindin-D9k in the uterus of immature rats. Toxicol. Sci 2004, 82, 451–457. [Google Scholar]
- Nguyen, T.H.; Lee, G.S.; Ji, Y.K.; Choi, K.C.; Lee, C.K.; Jeung, E.B. A calcium binding protein, calbindin-D9k, is mainly regulated by estrogen in the pituitary gland of rats during estrous cycle. Brain Res. Mol. Brain Res 2005, 141, 166–173. [Google Scholar]
- Seifert, M.F.; Gray, R.W.; Bruns, M.E. Elevated levels of vitamin D-dependent calcium-binding protein (calbindin-D9k) in the osteosclerotic (oc) mouse. Endocrinology 1988, 122, 1067–1073. [Google Scholar]
- Darwish, H.; Krisinger, J.; Furlow, J.D.; Smith, C.; Murdoch, F.E.; DeLuca, H.F. An estrogen-responsive element mediates the transcriptional regulation of calbindin D9K gene in rat uterus. J. Biol. Chem 1991, 266, 551–558. [Google Scholar]
- Delorme, A.C.; Danan, J.L.; Acker, M.G.; Ripoche, M.A.; Mathieu, H. In rat uterus 17 beta-estradiol stimulates a calcium-binding protein similar to the duodenal vitamin D-dependent calcium-binding protein. Endocrinology 1983, 113, 1340–1347. [Google Scholar]
- Price, N.T.; Smith, A.J.; Rogers, L.J. Relationship of the flavodoxin isoforms from. Porphyra umbilicalis. Phytochemistry 1991, 30, 2841–2843. [Google Scholar]
- Kim, S.; An, B.S.; Yang, H.; Jeung, E.B. Effects of octylphenol and bisphenol A on the expression of calcium transport genes in the mouse duodenum and kidney during pregnancy. Toxicology 2013, 303, 99–106. [Google Scholar]
- Brown, A.J.; Krits, I.; Armbrecht, H.J. Effect of age, vitamin D, and calcium on the regulation of rat intestinal epithelial calcium channels. Arch. BioChem. Biophys 2005, 437, 51–58. [Google Scholar]
- Vennekens, R.; Hoenderop, J.G.; Prenen, J.; Stuiver, M.; Willems, P.H.; Droogmans, G.; Nilius, B.; Bindels, R.J. Permeation and gating properties of the novel epithelial Ca2+ channel. J. Biol. Chem 2000, 275, 3963–3969. [Google Scholar]
- Li, Y.C.; Pirro, A.E.; Amling, M.; Delling, G.; Baron, R.; Bronson, R.; Demay, M.B. Targeted ablation of the vitamin D receptor: An animal model of vitamin D-dependent rickets type II with alopecia. Proc. Natl. Acad. Sci. USA 1997, 94, 9831–9835. [Google Scholar]
- Walters, J.R. Calbindin-D9k stimulates the calcium pump in rat enterocyte basolateral membranes. Am. J. Physiol 1989, 256, 124–128. [Google Scholar]
- Walters, J.R.; Howard, A.; Charpin, M.V.; Gniecko, K.C.; Brodin, P.; Thulin, E.; Forsen, S. Stimulation of intestinal basolateral membrane calcium-pump activity by recombinant synthetic calbindin-D9k and specific mutants. BioChem. Biophys. Res. Commun 1990, 170, 603–608. [Google Scholar]
- Lee, G.S.; Jung, E.M.; Choi, K.C.; Oh, G.T.; Jeung, E.B. Compensatory induction of the TRPV6 channel in a calbindin-D9k knockout mouse: Its regulation by 1,25-hydroxyvitamin D3. J. Cell Biochem 2009, 108, 1175–1183. [Google Scholar]
- Kim, M.H.; Lee, G.S.; Jung, E.M.; Choi, K.C.; Oh, G.T.; Jeung, E.B. Dexamethasone differentially regulates renal and duodenal calcium-processing genes in calbindin-D9k and-D28k knockout mice. Exp. Physiol 2009, 94, 138–151. [Google Scholar]
- Bianco, S.D.; Peng, J.B.; Takanaga, H.; Suzuki, Y.; Crescenzi, A.; Kos, C.H.; Zhuang, L.; Freeman, M.R.; Gouveia, C.H.; Wu, J.; et al. Marked disturbance of calcium homeostasis in mice with targeted disruption of the Trpv6 calcium channel gene. J. Bone Miner. Res 2007, 22, 274–285. [Google Scholar]
- Benn, B.S.; Ajibade, D.; Porta, A.; Dhawan, P.; Hediger, M.; Peng, J.B.; Jiang, Y.; Oh, G.T.; Jeung, E.B.; Lieben, L.; et al. Active intestinal calcium transport in the absence of transient receptor potential vanilloid type 6 and calbindin-D9k. Endocrinology 2008, 149, 3196–3205. [Google Scholar]




© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hong, E.-J.; Jeung, E.-B. Biological Significance of Calbindin-D9k within Duodenal Epithelium. Int. J. Mol. Sci. 2013, 14, 23330-23340. https://doi.org/10.3390/ijms141223330
Hong E-J, Jeung E-B. Biological Significance of Calbindin-D9k within Duodenal Epithelium. International Journal of Molecular Sciences. 2013; 14(12):23330-23340. https://doi.org/10.3390/ijms141223330
Chicago/Turabian StyleHong, Eui-Ju, and Eui-Bae Jeung. 2013. "Biological Significance of Calbindin-D9k within Duodenal Epithelium" International Journal of Molecular Sciences 14, no. 12: 23330-23340. https://doi.org/10.3390/ijms141223330
APA StyleHong, E. -J., & Jeung, E. -B. (2013). Biological Significance of Calbindin-D9k within Duodenal Epithelium. International Journal of Molecular Sciences, 14(12), 23330-23340. https://doi.org/10.3390/ijms141223330