2.1. Preliminary Screening for the Best Carbon Sources and Supplementary Nitrogen and Carbon Sources for Enzymes Production by C. graminicola
The best carbon source for β-glucosidase production was wheat bran (
Table 1). The levels of enzyme reached 109.7 ± 7.7 U g
−1 dry substrate and the fungus showed vigorous growth in this substrate. Good enzyme levels were also obtained in peanut hulls, corn and rice husks. Although the fungus grew poorly in SCT, with a low enzyme production, the supplementation of wheat bran with 1% (
w/
w) SCT resulted in 1.6-fold increase in enzyme production levels. Enhanced production (1.4-fold) also resulted from wheat bran supplementation with 1% (
w/
w) cellobiose, but filter paper and carboxymethylcellulose (CMC) showed negligible effects. Addition of organic and inorganic nitrogen sources to wheat bran inhibited β-glucosidase production, except for yeast extract and soybean meal, without effect (
Table 1).
Maximal levels of β-xylosidase were obtained in wheat bran (57.9 ± 4.6 U g
−1), but milled corncob, soybean meal and corn husks were also acceptable sources for enzyme production (
Table 1). Wheat bran supplementation with 1% peanut hulls increased the production by 42%. Various supplementary carbon sources, including xylan, were somewhat inhibitory, and 1% glucose, xylose or cellobiose reduced the enzyme production by about 40%. Most supplementary nitrogen sources inhibited β-xylosidase production, except for casein and yeast extract, without effect (
Table 1).
Wheat bran was also the best carbon source for xylanase production, reaching 189.3 ± 9.5 U g
−1, and low to negligible enzyme levels were obtained in all other carbon sources (
Table 1). Wheat bran supplementation with 1% milled corncob slightly enhanced xylanase production. Most other supplementary carbon sources tested, including xylan, were without effect, while xylose inhibited about 33% the production of the enzyme. The addition of 0.8% (NH
4)
2SO
4 and 1% peptone to wheat bran increased xylanase production (30% and 20%, respectively), while all other nitrogen sources tested inhibited the production, except for casein (
Table 1).
Several studies show that wheat bran is a good carbon source for the production of lignocellulolytic enzymes by different fungi under SSF, even in the absence of any supplementary carbon or nitrogen source [
26–
29]. This has been attributed to its particularly rich nutritional composition: vitamin B, about 14% proteins, 27% carbohydrates (64% cellulose and 36% hemicellulose), 6% lipids, 5% minerals, and around 64% digestible nitrogen [
30,
31].
2.2. Optimization of Enzyme Production by C. graminicola Using Response Surface Methodology (RSM)
The optimization of culture media by varying one independent variable at a time maintaining the others at constant levels (OFAT) results in considerable increases in enzyme productivity by microorganisms. However, besides being expensive and time consuming, this methodology does not show the interactions that may occur between different variables that affect enzyme production. In contrast, statistical approaches using response surface methodology not only allow the identification of the optimal conditions by a set of independent variables, but also reveal the interactions between two or more variables [
32].
After preliminary studies by OFAT to define the experimental ranges for each independent variable, the culture conditions for the production of β-glucosidase, β-xylosidase and xylanase were optimized using a Central Composite Rotational Design (CCRD).
Data obtained for each enzyme (
Tables 2,
3 and
4) were analyzed by multiple regression analysis, and the predicted responses for β-glucosidase (βglu), β-xylosidase (βxyl) and xylanase (Xyl) production were:
where x, y, z, w, k and s were the coded values for temperature, culture time, initial moisture, peanut hulls, milled corncob and (NH4)2SO4 concentrations, respectively.
Preliminary analyses by OFAT revealed that the presence of SCT (0.5% to 10%, w/w) in the culture medium enhanced the production of β-glucosidase by about 60%, compared to that obtained in wheat bran and water only (not shown). However, similar production was obtained for all SCT concentrations tested in this range. In agreement, when the production of β-glucosidase was optimized using RSM the concentration of SCT and its interactions with the other independent variables tested (temperature, culture time and initial moisture) has not significantly influenced the production (not shown). Thus, the concentration of SCT in the culture medium was fixed at 1% for further experiments. Similarly, the temperature and its interactions with the other independent variables tested have not significantly influenced the production of β-xylosidase (not shown), and the production of this enzyme was optimized with respect to culture time, initial moisture and peanut hulls concentration only.
The analyses of variance (ANOVA) for the response surface quadratic models for the production of each enzyme are summarized in
Table 5. The coefficients of the full model were analyzed for their significance and the non-significant ones were eliminated from the model. According to the
F test, the three models were predictive, since the calculated
F values were greater than the listed
F values. The low
P-values also indicate the significance of the models. Further,
P-values greater than 0.05 indicated that the lack of fit for the models was non-significant. The R
2 coefficients of 0.99 for β-glucosidase, 0.90 for β-xylosidase and 0.88 for xylanase production confirm the goodness of the three models, indicating that 99%, 90% and 88%, respectively, of the responses variability could be explained by them.
The regression coefficients obtained for β-glucosidase, β-xylosidase and xylanase production are listed in
Table 5. The analysis of the
P-values showed that the quadratic terms of all independent variables tested had significant effects on β-glucosidase production, but not their linear terms nor the interactions between them. The quadratic effects of culture time and initial moisture were the most important factors affecting β-glucosidase production.
Significant effects on β-xylosidase production were exerted by culture time and initial moisture, the interaction between them, and the interaction between peanut hulls concentration and initial moisture. The quadratic terms of all three independent variables also had significant effects. The linear effects of culture time and initial moisture were the most critical variables that affected the production of β-xylosidase.
Analysis of the P-values for the regression coefficients obtained for xylanase production revealed that temperature, culture time, initial moisture and (NH4)2SO4 concentration, and their quadratic terms, significantly affected the enzyme production. The quadratic term of milled corncob concentration and the interaction between temperature and initial moisture also had significant effects. The most important factors that affected xylanase production were the quadratic terms of temperature, culture time and initial moisture.
Among the factors that influence the enzymatic production under SSF, water content is well recognized as one of the most critical. The ideal moisture varies with the organism, the operational conditions and the solid substrate, affecting both the microbial growth and the production and secretion of enzymes. At low moisture content, lower substrate swelling occurs, reducing the superficial area. Moreover, the solubility and diffusion of the nutrients are reduced, as well as the stability of the extracellular enzymes, possibly impairing the growth of the microorganism. In contrast, moisture levels that are too high may cause aggregation of substrate particles, reducing the porosity and thus limiting oxygen availability, also hindering microbial growth and lowering enzyme production [
28,
33,
34].
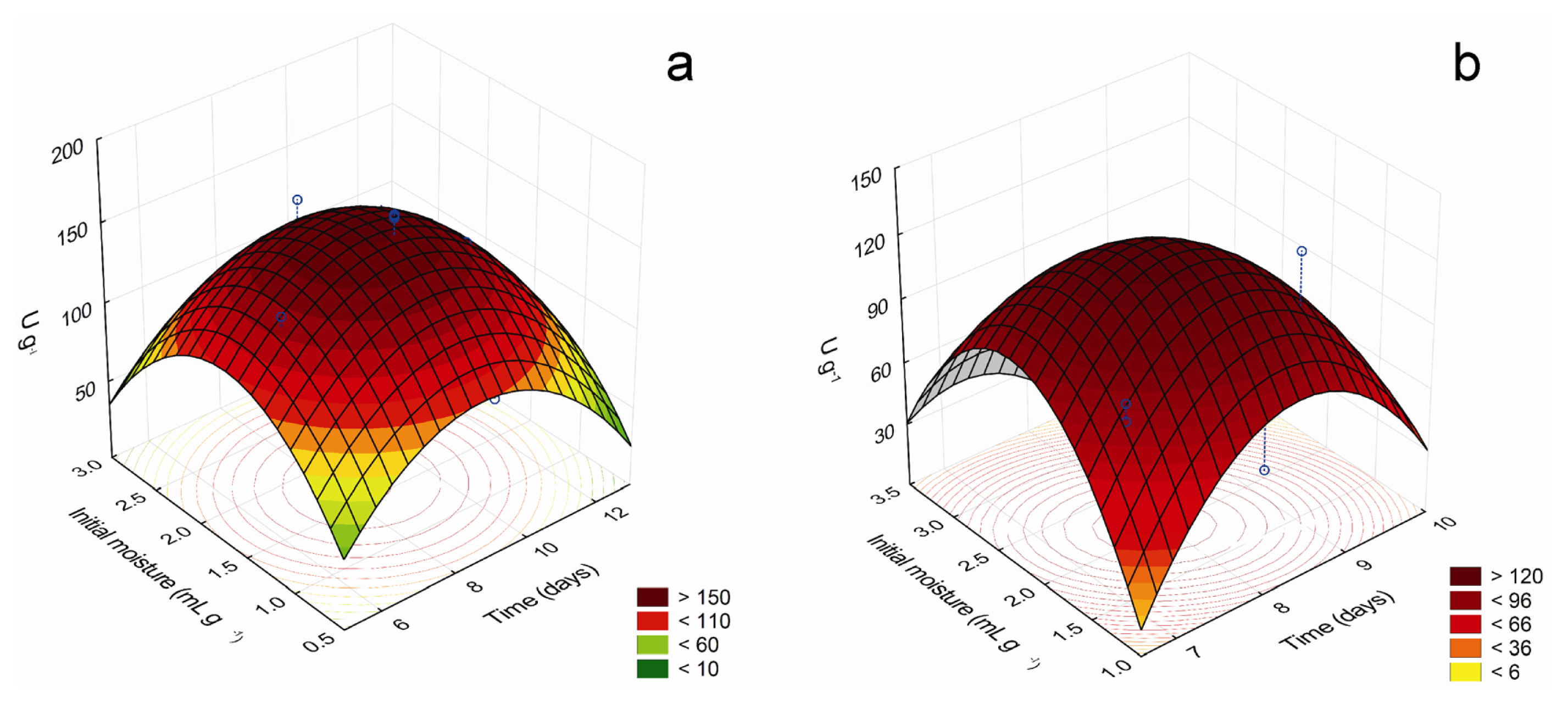
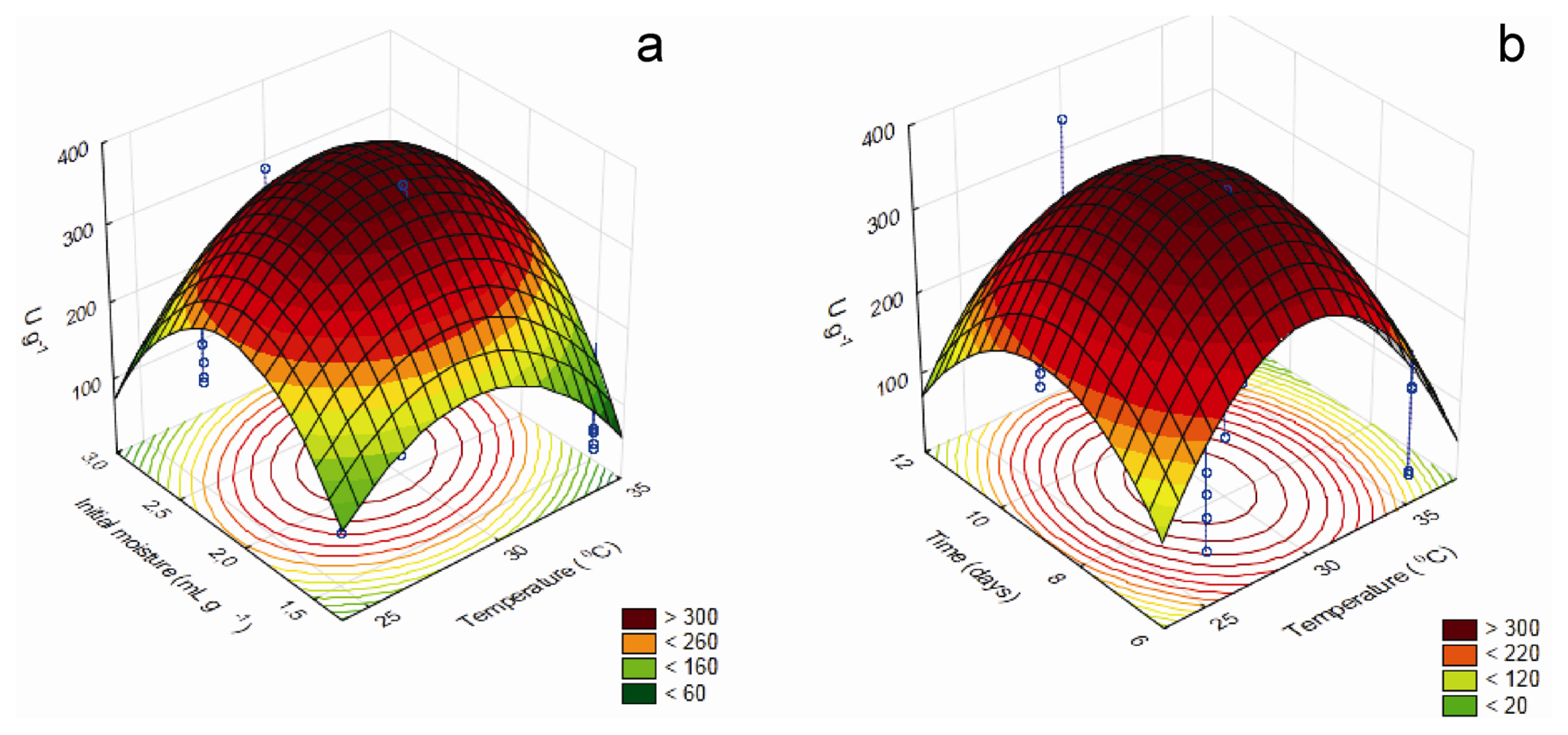
Tridimensional response surface curves graphically represent regression equations, demonstrating the relationships between the response and experimental levels of each variable and allowing a better understanding of the interactions between the significant independent variables. Moreover, they provide working ranges for each variable, depicting the robustness of a process,
i.e., its capability to bring the expected results in the presence of deviations from the optimal conditions or unexpected adverse factors. The 3D response curves for the most important factors that affect the production of β-glucosidase and β-xylosidase are shown in
Figure 1, while
Figure 2 brings those for the factors that influence xylanase production.
Statistical analysis showed that maximum β-glucosidase production could be achieved after 7.07 days at 28.13 °C, with initial moisture of 2.38 mL g−1, using wheat bran as carbon source and 1% (w/w) sugarcane trash as supplementary carbon source. Maximum predicted value was 156.63 U g−1, 1.4-fold higher than observed before optimization. The mean value of the experimental validation of the optimized conditions (159.3 ± 12.7 U g−1) was in excellent correlation with the predicted value, confirming the validity of the model. Interestingly, the production of β-glucosidase under optimal temperature, initial moisture and culture time decreased about 50% in the absence of SCT, but near maximal levels were obtained with SCT concentrations ranging from 1% to 10% (w/w) (not shown). Considering that SCT does not support the growth of C. graminicola, it is possible that when grown in wheat bran, the fungus partially degrades SCT, liberating little amounts of some compounds that stimulate enzyme production.
For β-xylosidase production, the maximum point predicted was 7.71 days, peanut hulls concentration of 4.66% (w/w) and initial moisture of 2.1 mL g−1, at 25 °C, using wheat bran as carbon source. Maximum predicted production was 125.88 U g−1, about 2.2-fold higher than that obtained initially. The mean value of the experimental validation was 128.1 ± 6.4 U g−1, in good agreement with the predicted value, proving the model’s adequacy.
According to the model, maximum xylanase production occurred at 29.7 °C, 8.66 days, initial moisture of 2.21 mL g−1, milled corncob concentration of 0.87% (w/w) and (NH4)2SO4 concentration of 0.378% (w/w), using wheat bran as carbon source. The maximal predicted value, 396.65 U g−1, was 2.1-fold higher than initially estimated, and showed good correlation with the result of the experimental validation (378.1 ± 23.3 U g−1), establishing the model’s validity.
The optimal production levels of β-glucosidase by
C. graminicola were higher than most reported in the literature for SSF processes, but values of the same magnitude were found for
Aspergillus fumigatus [
35] and
Trichoderma koningi F244 [
36] cultured in wheat bran moistened with mineral salt medium or supplemented with ammonium sulfate, respectively. Similar levels were also obtained for
Thermoascus aurantiacus grown in wheat straw [
37] and
Aspergillus terreus M11 in corn stover [
38], both supplemented with mineral solutions. Few fungi are good producers of β-xylosidases under SSF, but the production levels of the mutant strain
Aspergillus niger KK2 in rice straw supplemented with salts, yeast extract and corn steep liquor were similar to those found for
C. graminicola [
26]. Higher production levels were reported for a Brazilian strain of
Aspergillus tamarii cultured in wheat bran supplemented with Vogel salts [
39] and for the mutant
Humicola lanuginosa M7D cultured in corncob supplemented with Vogel salts and corn steep liquor [
40]. Similar levels of xylanase, compared to
C. graminicola, were produced by
A. niger in wheat bran supplemented with mineral salts [
36]. However, 6-fold higher levels of xylanase were produced by some strains of
A. niger and
T. reesei [
11] in wheat bran supplemented with Mandel Weber medium [
41].
Due to the high enzyme loads required for an efficient hydrolysis of lignocellulosic materials, it is estimated that enzyme costs contribute 23%–40% to cellulosic ethanol production cost, directly impacting the economic viability of the process [
11,
42]. The carbon source is one of the major factors affecting enzyme production costs [
38], and thus, there is a rising interest in fungi that produce cellulases and hemicellulases with good yields in inexpensive carbon sources [
11,
21]. In this context, our results characterize
C. graminicola as a very promising microorganism for industrial purposes, since few fungal strains produce similar or higher levels of β-glucosidase, β-xylosidase and xylanase under SSF [
26,
35,
38,
40]. Moreover, this good production was obtained in quite cheap media mostly composed of wheat bran, a low-cost industrial byproduct, without addition of any expensive nutritional supplement. The simultaneous production of significant levels of three enzymes involved in the hydrolysis of lignocellulose may be considered another advantage of this fungus, aimed at the reduction of production costs. However, it is well established that the hydrolysis of different lignocellulosic residues, which are highly heterogeneous, requires enzymatic cocktails of distinct compositions for maximal yields [
8,
25]. Thus, the use of multi-enzyme crude extracts has the disadvantage of limiting the possibility of varying enzyme proportions, making difficult to optimize a cocktail for a specific lignocellulosic residue. Moreover, the presence of contaminant enzymes may have deleterious effects for specific industrial processes.
2.3. Effect of Temperature and pH on β-Glucosidase, β-Xylosidase and Xylanase Activities and Thermal Stabilities
The effects of pH and temperature on the enzymatic activities in the crude extracts from
C. graminicola cultured under optimal conditions were evaluated using a CCRD for each enzyme, and RSM analyses. The experimental conditions and results of the experimental designs are summarized in
Table 6. Data analyses by multiple regressions led to predicted responses for β-glucosidase (βglu), β-xylosidase (βxyl) and xylanase (Xyl) activities as follows:
The analyses of variance (ANOVA) for the response surface quadratic models are summarized in
Table 7. Based on
F test, the models were predictive of the enzymatic activities as a function of pH and temperature, given the higher values for calculated
F values compared to listed ones. The low
p-values also indicate the models’ significance. Further, the
p-values greater than 0.05 indicated that the lack of fit for the models was non-significant. The R
2 coefficients of 0.99 for β-glucosidase, 0.99 for β-xylosidase and 0.98 for xylanase activities confirm the goodness of the models.
The regression coefficients (
Table 7) showed low
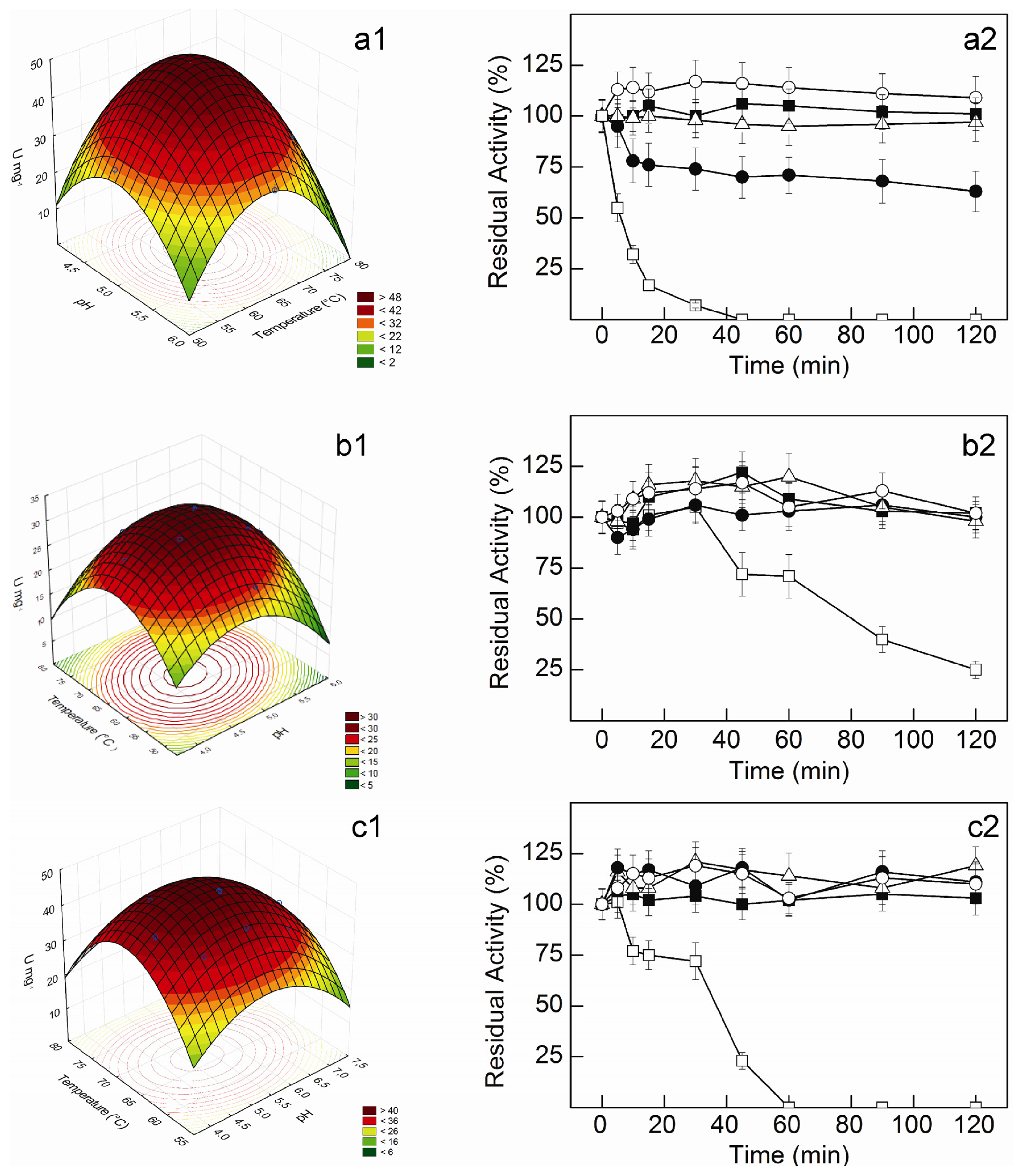
p-values for all linear and quadratic effects, and also for the interaction between pH and temperature, indicating their significant effect on enzymatic activities. The pH effects were higher than the temperature effects for the three activities, and working ranges for near optimal activity may be easily anticipated from the response surface curves for each enzymatic activity (
Figure 3). Indeed, this is a clear advantage of using RSM to estimate optimal temperature and pH for enzymatic activity, providing higher flexibility in the development of different bioprocesses [
43].
The statistical analysis showed that optimum pH and temperature for β-glucosidase activity were 4.94 and 64.61 °C, and the maximum predicted value was 51.16 U mg−1, very close to that obtained in the experimental validation (51.2 ± 1.6 U mg−1), confirming the model validity. β-xylosidase activity was maximal at pH 4.77 and 63.33 °C, with a predicted value of 32.59 U mg−1, in excellent correlation with the value of experimental validation (32.4 ± 0.8 U mg−1). Further, optimal pH and temperature for xylanase activity were 5.21 and 66.91 °C with a predicted value of 49.1 U mg−1, in close agreement with the experimental validation, 49.3 ± 1.4 U mg−1.
pH optima determined for
C. graminicola enzymes are in the range found for most fungal β-glucosidases [
27,
44], β-xylosidases [
17] and xylanases [
27]. Moreover, their elevated optimal temperatures are highly desirable for industrial applications.
The β-glucosidase from
C. graminicola maintained about 100% of the initial activity up to 72 h at 50 °C. Moreover, the enzyme was fully stable for 120 min at 55–60 °C. At 65 °C, a rapid decrease to about 80% of the initial activity occurred after 10 min, but a residual activity of 63% remained after 120 min (
Figure 3a2). Higher thermal stabilities were observed for the β-xylosidase and the xylanase. Besides retaining full activity for at least 72 h at 50 °C, these enzymes remained stable for 120 min at 55–65 °C (
Figure 3b2,c2). At 70 °C, however, half-lives of 6.1, 79.0 and 36.9 min were estimated for β-glucosidase, β-xylosidase and xylanase activities, respectively.
The β-glucosidase from
C. graminicola was more thermostable than most fungal enzymes studied to date, including those from various thermophilic fungi [
44–
48]; Few β-glucosidases with higher thermal stabilities were reported [
31,
49,
50]. The β-xylosidase also showed higher thermal stability than reported for enzymes from different thermophilic species [
51–
56], but somewhat more thermostable enzymes were produced by a few
Aspergillus genera [
17] and
Humicola lanuginosa [
40]. Similarly,
C. graminicola xylanase was more thermostable than several enzymes from different thermophiles, with half-lives in the range of 4–115 min at 60 °C [
57–
60]. Some highly thermostable xylanases are, however, produced by
Thermomyces lanuginosus THKU-49 [
61] and
Talaromyces thermophilus [
62], with half-lives of 336 min at 70 °C and 1.0 h at 100 °C, respectively. The high thermal stability of
C. graminicola β-glucosidase, β-xylosidase and xylanase constitutes another very interesting feature for their employment in biotechnological processes.
2.4. Synergic Action of C. graminicola Crude Extract and T. reesei Cellulases on Raw Sugarcane Trash Hydrolysis
Initially, the optima of temperature and pH, and the thermal stability of the cellulases in the crude extract from T. reesei were studied. The optimal pH for cellulase activity was 4.5 to 5.0, and the optimal temperature was 50–55 °C, with a sharp decrease at 60 °C. About 70% of T. reesei cellulases activity was retained after 72 h at 50 °C, but at higher temperatures, low residual activities were estimated after 24 h (not shown). Considering that the β-glucosidase, β-xylosidase and xylanase in C. graminicola crude extract showed optimal pH around 5.0, where they were fully stable at 50 °C up to 72 h and presented about 55% of their maximal activity at 50 °C, the conditions chosen for RSCT hydrolysis were 50 °C and pH 5.0.
The crude extracts from T. reesei and C. graminicola were then characterized for specific activities at 50 °C and pH 5.0. Trichoderma reesei extract showed good levels of CMCase (3.7 ± 0.5 U mL−1), FPA (0.29 ± 0.04 U mL−1) and avicelase (0.30 ± 0.05 U mL−1), but low levels of cellobiase (0.002 U mL−1) and undetectable levels of the other activities tested. In contrast, low levels of cellulases were found in the extract from C. graminicola (0.28 ± 0.04 U mL−1 CMCase; 0.08 ± 0.02 FPU mL−1; 0.044 ± 0.008 U mL−1 avicelase), which showed high levels of cellobiase (7.6 ± 0.9 U mL−1), β-xylosidase (5.0 ± 0.8 U mL−1) and xylanase (34.6 ± 4.9 U mL−1) activities, as well as some α-L-arabinofuranosidase activity (0.11 ± 0.02 U mL−1). Similar profiles were obtained for C. graminicola crude extracts produced at optimized conditions for β-glucosidase, β-xylosidase or xylanase production, despite some variation in the proportions of the main enzymes (not shown).
The time course of RSCT hydrolysis using
C. graminicola and
T. reesei crude extracts is shown in
Figure 4. Treatment with
C. graminicola extract only resulted in maximal yields of 11.3% reducing sugars and 4.4% glucose after 72 h. Higher yields were obtained using
T. reesei extract, attaining 18.3% reducing sugars and 18.7% glucose. However, when using a mixture of both extracts, a synergic effect was observed, and yields of 24.6% reducing sugars and 25.5% glucose were attained just after 24 h. Moreover, maximal yields reached 37.6% reducing sugars and 33.1% glucose, after 48 h. The low yields of glucose obtained using
C. graminicola crude extract reflect its low levels of cellulases. The inefficient hydrolysis of cellulose results in low production of cellobiose and, consequently, low glucose release by β-glucosidases, although present at high levels. The higher levels of reducing sugars compared to glucose thus possibly reflect the action of xylanases and β-xylosidases on RSCT, generating xyloligosaccharides and xylose. In contrast, the reducing sugars obtained using the cellulase-rich crude extract from
T. reesei must derive from cellulose hydrolysis. Some glucose was also obtained, since this extract presents low levels of β-glucosidases, but the plateau of reducing sugars observed after 48 h suggests accumulation of cellobiose, inhibiting endo- and exo-glucanases, as reported by others [
15,
25]. The synergic effect observed when a mixture of the two crude extracts was used may be explained by the presence of good levels of xylanases, β-xylosidases and β-glucosidases (
C. graminicola extract), as well as cellulases (
T reesei extract), as proposed by other authors [
8,
10,
13,
14]. The hydrolysis of xylan by the action of xylanases exposes the cellulosic fibers to cellulases, resulting in higher efficiency of cellulose hydrolysis and liberation of good levels of cellobiose. Efficient cellobiose hydrolysis by β-glucosidases then generates good levels of glucose and releases cellulases from product inhibition. Simultaneously, cellulose degradation increases xylan exposure to xylanases, amplifying the synergic effect. Moreover, the hydrolysis of xylan and xyloligosaccharides to xylose releases the cellulases from inhibition by these compounds [
13]. The net result is a faster hydrolysis of RSCT with higher yields of both glucose and reducing sugars. The maximal hydrolysis yield, however, is limited by the high content of lignin in RSCT (20%–36%), that hinders an efficient action of the hydrolytic enzymes [
3,
42,
63,
64].
Despite its increasing importance as a sustainable feedstock for cellulosic ethanol production, little information is available on SCT, and few authors have studied its enzymatic hydrolysis. Moreover, most studies reported were carried out using SCT submitted to different pre-treatments to increase the accessibility of cellulose and hemicelluloses to the enzymes [
3,
42,
63–
65]. Glucose yields of about 80% to 90% were attained using SCT pre-treated with dilute acid [
3,
42], steam [
63] or ammonia [
64], or submitted to milling [
65]. In contrast, to the best of our knowledge, a single study provided some data on RSCT hydrolysis [
65], reporting a maximal glucose yield of 23.5% after 72 h using a cocktail of commercial enzymes.
The higher efficiency of our cocktail for RSCT hydrolysis, compared to that used by Sant’Ana da Silva
et al. [
65], may be at least in part attributed to the high content of xylanases and β-xylosidases, besides β-glucosidases, in
C. graminicola crude extract. Similar FPU loads (15 FPU g
−1 RSCT) were used in both studies. Further, a CMCase load about 1.7-fold lower was employed in this study. In contrast, the cellobiase and xylanase loads were about 3-fold higher, and our cocktail also contained around 225 U g
−1 of β-xylosidase activity, almost absent (1.4 U g
−1) from the other cocktail. Thus, our higher glucose yield may be directly related to a higher efficiency of xylan degradation, resulting in an improved action of cellulases. Indeed, the xylan content of SCT is around 23% (
w/
w) [
5,
64], and the composition of
C. graminicola crude extract seems to be naturally very well suited for the hydrolysis of this particular lignocellulosic residue, with the addition of crude cellulases only.
Finally, considering that the conditions for the hydrolysis of RSCT (incubation time, biomass loading, enzyme loading,
etc.) using the
C. graminicola cocktail were not yet optimized, increases in glucose yields are to be expected. Moreover, the addition of some other enzymes to the cocktail, such as ligninases, may increase the hydrolysis efficiency, as well as some mild, low-cost pre-treatment of SCT. Thus, this low-cost cellulolytic-hemicelulolytic enzyme system seems promising for future industrial use. However, its real practical applicability depends on several factors, including the success of scaling up the processes of enzyme production and SCT hydrolysis. Further, the implementation of any industrial process of enzymatic hydrolysis of a lignocellulosic residue for ethanol production depends on a complete techno-economic evaluation of the final production costs [
5].

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}