The Increasing Complexity of the Oncofetal H19 Gene Locus: Functional Dissection and Therapeutic Intervention

{kind=link}
{kind=link}
Abstract
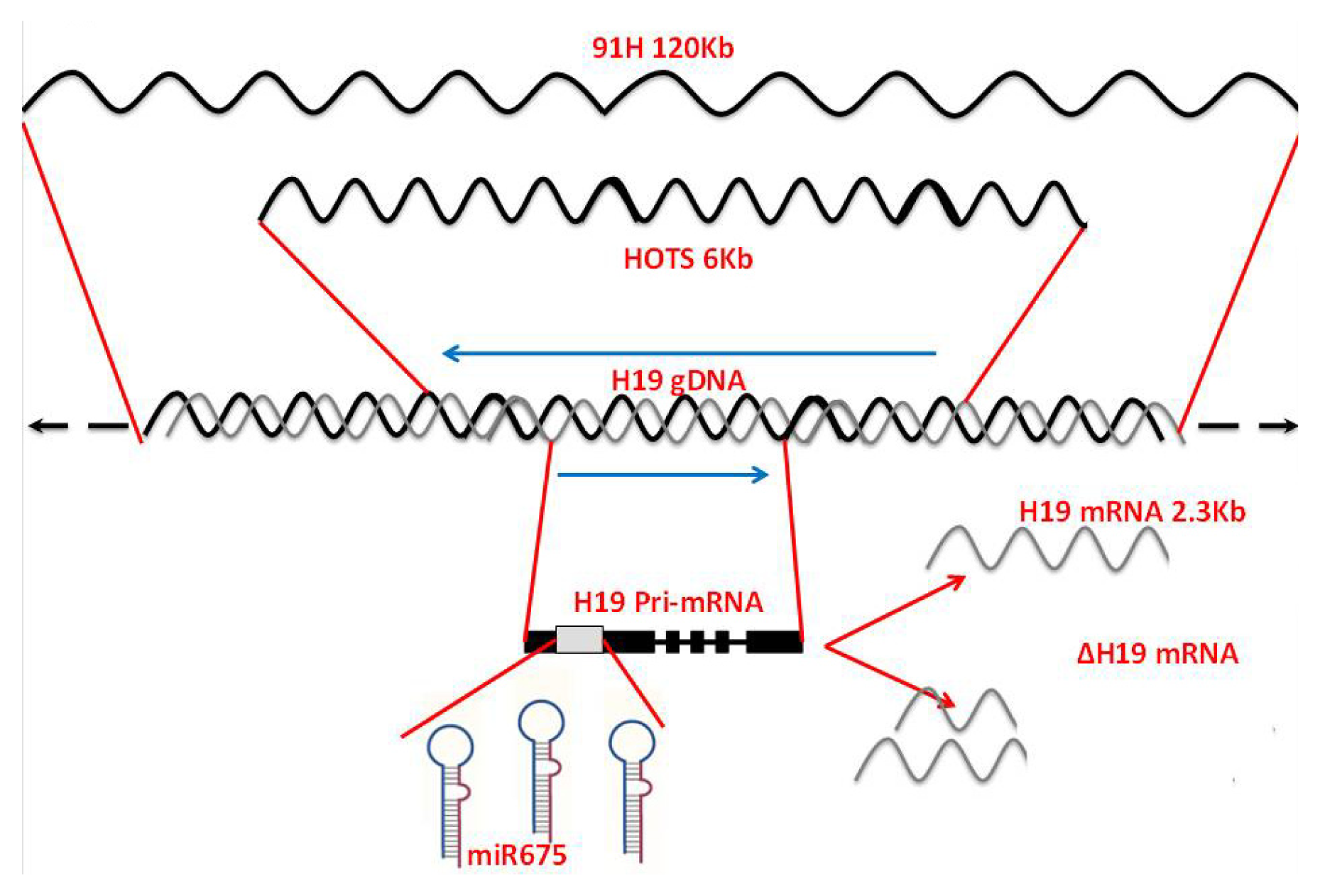
:1. The Increasing Complexity of the H19 Gene Locus
1.1. H19 Opposite Tumor Suppressor (HOTS)
1.2. Identification of 91H: A Long Intergenic Antisense RNA
1.3. H19 Is a Primary MicroRNA Precursor
1.4. Alternative Splice Isoforms of the H19 Gene
2. Transcriptional Control of the Oncofetal H19 Gene: Interplay of Factors Critical for the Tumorigenic Process
2.1. The Oncofetal H19 RNA Connection: Hypoxia, p53, HIF1-α and Cancer
2.2. c-Myc Oncogene Directly Induces H19 to Potentiate Tumorigenesis
2.3. E2F Activates H19 Promoter
3. H19 Gene Expression in Non-Cancerous Diseases
3.1. Beckwith–Wiedemann Syndrome
3.2. Russell—Silver Syndrome
3.3. Obesity, Overweight and H19
3.4. Inflammation, Rheumatoid Arthritis and H19
3.5. H19 Involvement in Pre-Eclampsia
4. Therapeutic Applications Based on H19 Gene
- transitional cell carcinoma (TCC) of the bladder;
- pancreatic cancer;
- ovarian cancer; and
- hepatic metastases
4.1. Bladder Cancer
4.2. Pancreatic Cancer
4.3. Ovarian Cancer
4.4. Colon Liver Metastases
5. Conclusions and Future Perspectives
Acknowledgments
Conflict of Interest
References
- Cai, X.; Cullen, B.R. The imprinted H19 noncoding RNA is a primary microRNA precursor. RNA 2007, 13, 313–316. [Google Scholar]
- Onyango, P.; Feinberg, A.P. A nucleolar protein, H19 opposite tumor suppressor (HOTS), is a tumor growth inhibitor encoded by a human imprinted H19 antisense transcript. Proc. Natl. Acad. Sci. USA 2011, 108, 16759–16764. [Google Scholar]
- Berteaux, N.; Aptel, N.; Cathala, G.; Genton, C.; Coll, J.; Daccache, A.; Spruyt, N.; Hondermarck, H.; Dugimont, T.; Curgy, J.J.; et al. A novel H19 antisense RNA overexpressed in breast cancer contributes to paternal IGF2 expression. Mol. Cell Biol 2008, 28, 6731–6745. [Google Scholar]
- Court, F.; Baniol, M.; Hagege, H.; Petit, J.S.; Lelay-Taha, M.N.; Carbonell, F.; Weber, M.; Cathala, G.; Forne, T. Long-range chromatin interactions at the mouse IGF2/H19 locus reveal a novel paternally expressed long non-coding RNA. Nucleic Acids Res 2011, 39, 5893–5906. [Google Scholar]
- Matouk, I.; Ayesh, B.; Schneider, T.; Ayesh, S.; Ohana, P.; de-Groot, N.; Hochberg, A.; Galun, E. Oncofetal splice-pattern of the human H19 gene. Biochem. Biophys. Res. Commun 2004, 318, 916–919. [Google Scholar]
- Lin, W.L.; He, X.B.; Svensson, K.; Adam, G.; Li, Y.M.; Tang, T.W.; Paldi, A.; Pfeifer, S.; Ohlsson, R. The genotype and epigenotype synergize to diversify the spatial pattern of expression of the imprinted H19 gene. Mech. Dev 1999, 82, 195–197. [Google Scholar]
- Okutsu, T.; Kuroiwa, Y.; Kagitani, F.; Kai, M.; Aisaka, K.; Tsutsumi, O.; Kaneko, Y.; Yokomori, K.; Surani, M.A.; Kohda, T.; et al. Expression and imprinting status of human PEG8/IGF2AS, a paternally expressed antisense transcript from the IGF2 locus, in Wilms’ tumors. J. Biochem 2000, 127, 475–483. [Google Scholar]
- Lee, M.P.; DeBaun, M.R.; Mitsuya, K.; Galonek, H.L.; Brandenburg, S.; Oshimura, M.; Feinberg, A.P. Loss of imprinting of a paternally expressed transcript, with antisense orientation to KVLQT1, occurs frequently in Beckwith–Wiedemann syndrome and is independent of insulin-like growth factor II imprinting. Proc. Natl. Acad. Sci. USA 1999, 96, 5203–5208. [Google Scholar]
- Smilinich, N.J.; Day, C.D.; Fitzpatrick, G.V.; Caldwell, G.M.; Lossie, A.C.; Cooper, P.R.; Smallwood, A.C.; Joyce, J.A.; Schofield, P.N.; Reik, W.; et al. A maternally methylated CpG island in KvLQT1 is associated with an antisense paternal transcript and loss of imprinting in Beckwith–Wiedemann syndrome. Proc. Natl. Acad. Sci. USA 1999, 96, 8064–8069. [Google Scholar]
- Smits, G.; Mungall, A.J.; Griffiths-Jones, S.; Smith, P.; Beury, D.; Matthews, L.; Rogers, J.; Pask, A.J.; Shaw, G.; VandeBerg, J.L.; et al. Conservation of the H19 noncoding RNA and H19-IGF2 imprinting mechanism in therians. Nat. Genet 2008, 40, 971–976. [Google Scholar]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and IGF1R. Nat. Cell Biol 2012, 14, 659–665. [Google Scholar]
- Gao, W.L.; Liu, M.; Yang, Y.; Yang, H.; Liao, Q.; Bai, Y.; Li, Y.X.; Li, D.; Peng, C.; Wang, Y.L. The imprinted H19 gene regulates human placental trophoblast cell proliferation via encoding miR-675 that targets Nodal Modulator 1 (NOMO1). RNA Biol 2012, 9, 1002–1010. [Google Scholar]
- Tsang, W.P.; Ng, E.K.; Ng, S.S.; Jin, H.; Yu, J.; Sung, J.J.; Kwok, T.T. Oncofetal H19-derived miR-675 regulates tumor suppressor RB in human colorectal cancer. Carcinogenesis 2010, 31, 350–358. [Google Scholar]
- Dudek, K.A.; Lafont, J.E.; Martinez-Sanchez, A.; Murphy, C.L. Type II collagen expression is regulated by tissue-specific miR-675 in human articular chondrocytes. J. Biol. Chem 2010, 285, 24381–24387. [Google Scholar]
- Matouk, I.; Ohana, P.; Ayesh, S.; Hochberg, A. The oncofetal H19 RNA in human cancer, from the bench to the patient. Cancer Ther 2005, 3, 249–266. [Google Scholar]
- Matouk, I.J.; Ohana, P.; Galun, E.; Hochberg, A. The Pivotal Role of the Oncofetal H19 RNA in Human Cancer, A New Hope. Gene Therapy and Cancer Research Focus; Nova Science, Publisher: New York, NY, USA, 2008; pp. 241–260. [Google Scholar]
- Matouk, I.J.; deGroot, N.; Mezan, S.; Ayesh, S.; Abu-Lail, R.; Hochberg, A.; Galun, E. The H19 non-coding RNA is essential for human tumor growth. PLoS One 2007, 2, e845. [Google Scholar]
- Matouk, I.J.; Mezan, S.; Mizrahi, A.; Ohana, P.; Abu-Lail, R.; Galun, E.; Hochberg, A. The oncofetal H19 RNA connection: Hypoxia, p53 and cancer. Biochim. Biophys. Acta 2010, 1803, 443–451. [Google Scholar]
- Barsyte-Lovejoy, D.; Lau, S.K.; Boutros, P.C.; Khosravi, F.; Jurisica, I.; Andrulis, I.L.; Tsao, M.S.; Penn, L.Z. The c-Myc oncogene directly induces the H19 noncoding RNA by allele-specific binding to potentiate tumorigenesis. Cancer Res 2006, 66, 5330–5337. [Google Scholar]
- Ayesh, S.; Matouk, I.; Schneider, T.; Ohana, P.; Laster, M.; Hochberg, A. Possible physiological role of H19 RNA. Mol. Carcinog 2002, 35, 63–74. [Google Scholar]
- Berteaux, N.; Lottin, S.; Monté, D.; Pinte, S.; Quatannens, B.; Coll, J.; Hondermarck, H.; Curgy, J.J.; Dugimont, T.; Adriaenssens, E. H19 mRNA-like noncoding RNA promotes breast cancer cell proliferation through positive control by E2F1. J. Biol. Chem 2005, 280, 29625–26636. [Google Scholar]
- Dugimont, T.; Montpellier, C.; Adriaenssens, E.; Lottin, S.; Dumont, L.; Iotsova, V.; Lagrou, C.; Stéhelin, D.; Coll, J.; Curgy, J.J. The H19 TATA-less promoter is efficiently repressed by the wild- type tumor suppressor gene product p53. Oncogene 1998, 16, 2395–2401. [Google Scholar]
- Choufani, S.; Shuman, C.; Weksberg, R. Beckwith–Wiedemann syndrome. Am. J. Med. Genet. C 2010, 154, 343–354. [Google Scholar]
- Joyce, J.A.; Lam, W.K.; Catchpoole, D.J.; Jenks, P.; Reik, W.; Maher, E.R.; Schofield, P.N. Imprinting of IGF2 and H19: Lack of reciprocity in sporadic Beckwith–Wiedemann Syndrome. Hum. Mol. Genet 1997, 6, 1543–1548. [Google Scholar]
- Eggermann, T.; Begemann, M.; Binder, G.; Spengler, S. Silver-Russell syndrome: Genetic basis and molecular genetic testing. Orphanet. J. Rare Dis 2010, 5, 19. [Google Scholar]
- Gabory, A.; Ripoche, M.A.; le Digarcher, A.; Watrin, F.; Ziyyat, A.; Forné, T.; Jammes, H.; Ainscough, J.F.; Surani, M.A.; Journot, L.; Dandolo, L. H19 acts as a trans regulator of the imprinted gene network controlling growth in mice. Development 2009, 136, 3413–3421. [Google Scholar]
- Schonherr, N.; Binder, G.; Korsch, E.; Kammerer, E.; Wollmann, H.A.; Eggermann, T. Are H19 variants associated with Silver-Russell syndrome? J. Pediatr. Endocrinol. Metab 2008, 21, 985–993. [Google Scholar]
- Perkins, E.; Murphy, S.K.; Murtha, A.P.; Schildkraut, J.; Jirtle, R.L.; Demark-Wahnefried, W.; Forman, M.R.; Kurtzberg, J.; Overcash, F.; Huang, Z.; Hoyo, C. Insulin-like growth factor 2/H19 methylation at birth and risk of overweight and obesity in children. J. Pediatr 2012, 161, 31–39. [Google Scholar]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar]
- Stuhlmüller, B.; Kunisch, E.; Franz, J.; Martinez-Gamboa, L.; Hernandez, M.M.; Pruss, A.; Ulbrich, N.; Erdmann, V.A.; Burmester, G.R.; Kinne, R.W. Detection of oncofetal H19 RNA in rheumatoid arthritis synovial tissue. Am. J. Pathol 2003, 163, 901–911. [Google Scholar]
- Smitten, A.L.; Simon, T.A.; Hochberg, M.C.; Suissa, S. A meta-analysis of the incidence of malignancy in adult patients with rheumatoid arthritis. Arthritis Res. Ther 2008, 10, R45. [Google Scholar]
- Love, T.; Solomon, D.H. The relationship between cancer and rheumatoid arthritis: Still a large research agenda. Arthritis Res. Ther 2008, 10, 109. [Google Scholar]
- Bergamaschi, S.; Morato, E.; Bazzo, M.; Neves, F.; Fialho, S.; Castro, G.; Zimmermann, A.; Pereira, I. Tumor markers are elevated in patients with rheumatoid arthritis and do not indicate presence of cancer. Int. J. Rheum. Dis 2012, 15, 179–182. [Google Scholar]
- Carty, D.M.; Delles, C.; Dominiczak, A.F. Novel biomarkers for predicting preeclampsia. Trends Cardiovasc. Med 2008, 18, 186–194. [Google Scholar]
- Eggenschwiler, J.; Ludwig, T.; Fisher, P.; Leighton, P.A.; Tilghman, S.M.; Efstratiadis, A. Mouse mutant embryos overexpressing IGF-II exhibit phenotypic features of the Beckwith–Wiedemann and Simpson-Golabi-Behmel syndromes. Genes Dev 1997, 11, 3128–3142. [Google Scholar]
- Yu, L.; Chen, M.; Zhao, D.; Yi, P.; Lu, L.; Han, J.; Zheng, X.; Zhou, Y.; Li, L. The H19 gene imprinting in normal pregnancy and pre-eclampsia. Placenta 2009, 30, 443–347. [Google Scholar]
- Vaiman, D.; Mondon, F.; Garcès-Duran, A.; Mignot, T.M.; Robert, B.; Rebourcet, R.; Jammes, H.; Chelbi, S.T.; Quetin, F.; Marceau, G. Hypoxia-activated genes from early placenta are elevated in Preeclampsia, but not in Intra-Uterine Growth Retardation. BMC Genomics 2005, 6, 111. [Google Scholar]
- Bourque, D.K.; Avila, L.; Peñaherrera, M.; von Dadelszen, P.; Robinson, W.P. Decreased placental methylation at the H19/IGF2 imprinting control region is associated with normotensive intrauterine growth restriction but not preeclampsia. Placenta 2010, 31, 197–202. [Google Scholar]
- Gao, W.L.; Li, D.; Xiao, Z.X.; Liao, Q.P.; Yang, H.X.; Li, Y.X.; Ji, L.; Wang, Y.L. Detection of global DNA methylation and paternally imprinted H19 gene methylation in preeclamptic placentas. Hypertens Res 2011, 34, 655–661. [Google Scholar]
- Rachmilewitz, J.; Gileadi, O.; Eldar-Geva, T.; Schneider, T.; De-Groot, N.; Hochberg, A. Transcription of the H19 gene in differentiating cytotrophoblasts from human placenta. Mol. Reprod. Dev 1992, 32, 196–202. [Google Scholar]
- Adam, G.I.; Cui, H.; Miller, S.J.; Flam, F.; Ohlsson, R. Allele-specific in situ hybridization (ASISH) analysis: A novel technique which resolves differential allelic usage of H19 within the same cell lineage during human placental development. Development 1996, 122, 839–847. [Google Scholar]
- Walsh, C.; Miller, S.J.; Flam, F.; Fisher, R.A.; Ohlsson, R. Paternally derived H19 is differentially expressed in malignant and nonmalignant trophoblast. Cancer Res 1995, 55, 1111–1116. [Google Scholar]
- Smets, E.M.; Visser, A.; Go, A.T.; van Vugt, J.M.; Oudejans, C.B. Novel biomarkers in preeclampsia. Clin. Chim. Acta 2006, 364, 22–32. [Google Scholar]
- Sidi, A.; Ohana, P.; Benjamin, S.; Shalev, M.; Ransom, J.H.; Lamm, D.; Hochberg, A.; Leibovitch, I. Phase I/II marker lesion study of intravesical BC-819 DNA plasmid in H19 overexpressing superficial bladder cancer refractory to Bacillus calmette guerin. J. Urology 2008, 180, 2379–2383. [Google Scholar]
- Hanna, N.; Ohana, P.; Konikoff, F.M.; Leichtmann, G.; Hubert, A.; Appelbaum, L.; Kopelman, Y.; Czerniak, A.; Hochberg, A. Phase 1/2a, dose-escalation, safety, pharmacokinetic, and preliminary efficacy study of intratumoral administration of BC-819 in patients with unresectable pancreatic vancer. Cancer Gene Ther 2012, 19, 374–381. [Google Scholar]
- Mizrahi, A.; Czerniak, A.; Ohana, P.; Amiur, S.; Gallula, J.; Matouk, I.; Abu-lail, R.; Birman, T.; Hochberg, A.; Levy, T. Treatment of ovarian cancer ascites by intra-peritoneal injection of diphtheria toxin A chain-H19 vector: A case report. J. Med. Case Rep 2010, 4, 228. [Google Scholar]


© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Matouk, I.; Raveh, E.; Ohana, P.; Lail, R.A.; Gershtain, E.; Gilon, M.; De Groot, N.; Czerniak, A.; Hochberg, A. The Increasing Complexity of the Oncofetal H19 Gene Locus: Functional Dissection and Therapeutic Intervention. Int. J. Mol. Sci. 2013, 14, 4298-4316. https://doi.org/10.3390/ijms14024298
Matouk I, Raveh E, Ohana P, Lail RA, Gershtain E, Gilon M, De Groot N, Czerniak A, Hochberg A. The Increasing Complexity of the Oncofetal H19 Gene Locus: Functional Dissection and Therapeutic Intervention. International Journal of Molecular Sciences. 2013; 14(2):4298-4316. https://doi.org/10.3390/ijms14024298
Chicago/Turabian StyleMatouk, Imad, Eli Raveh, Patricia Ohana, Rasha Abu Lail, Eitan Gershtain, Michal Gilon, Nathan De Groot, Abraham Czerniak, and Abraham Hochberg. 2013. "The Increasing Complexity of the Oncofetal H19 Gene Locus: Functional Dissection and Therapeutic Intervention" International Journal of Molecular Sciences 14, no. 2: 4298-4316. https://doi.org/10.3390/ijms14024298
APA StyleMatouk, I., Raveh, E., Ohana, P., Lail, R. A., Gershtain, E., Gilon, M., De Groot, N., Czerniak, A., & Hochberg, A. (2013). The Increasing Complexity of the Oncofetal H19 Gene Locus: Functional Dissection and Therapeutic Intervention. International Journal of Molecular Sciences, 14(2), 4298-4316. https://doi.org/10.3390/ijms14024298