Global Gene Expression Profiling Reveals Functional Importance of Sirt2 in Endothelial Cells under Oxidative Stress


Abstract
:Abbreviations
| eNOS | nitric oxide synthase |
| NF-κB | nuclear factor-κB |
| HUVEC | human umbilical vein endothelial cell |
| siRNA | small interfering RNA |
| qPCR | quantitative polymerase chain reaction |
| GO | Gene Ontology |
| GPx | glutathione peroxidase |
| SOD | superoxide dismutase |
1. Introduction
2. Results and Discussion
3. Experimental Section
3.1. Cell Culture
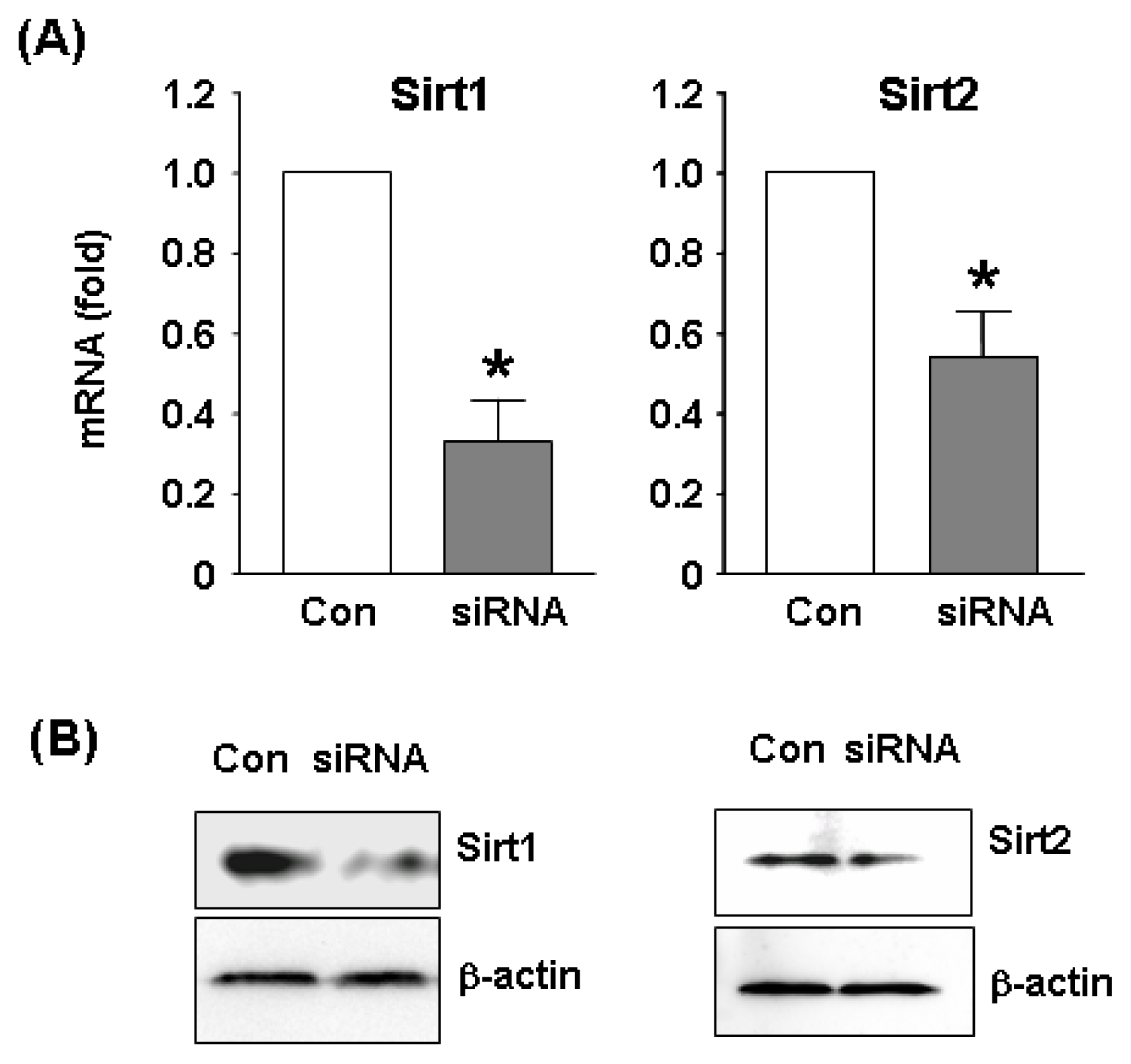
3.2. RNA Interference
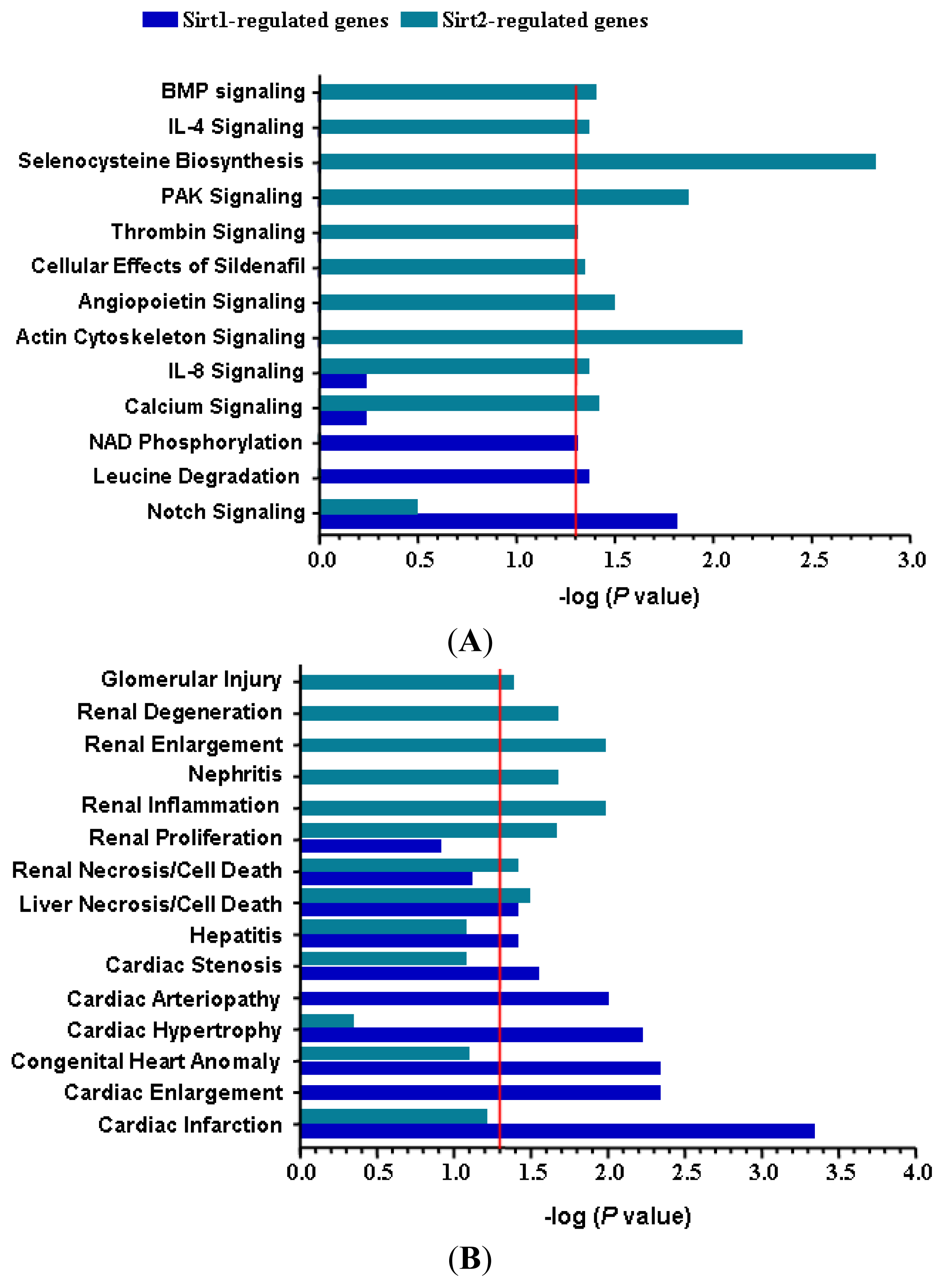
3.3. Microarray Experiments and Data Processing
3.4. Real-Time qPCR
3.5. Fluorescent Immunocytochemistry
3.6. Western Blot
3.7. Data and Statistics
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Haigis, M.C.; Sinclair, D.A. Mammalian sirtuins: Biological insights and disease relevance. Annu. Rev. Pathol 2010, 5, 253–295. [Google Scholar]
- Finkel, T.; Deng, C.X.; Mostoslavsky, R. Recent progress in the biology and physiology of sirtuins. Nature 2009, 460, 587–591. [Google Scholar]
- Saunders, L.R.; Verdin, E. Sirtuins: Critical regulators at the crossroads between cancer and aging. Oncogene 2007, 26, 5489–5504. [Google Scholar]
- Michan, S.; Sinclair, D. Sirtuins in mammals: Insights into their biological function. Biochem. J 2007, 404, 1–13. [Google Scholar]
- Bao, J.; Sack, M.N. Protein deacetylation by sirtuins: Delineating a post-translational regulatory program responsive to nutrient and redox stressors. Cell Mol. Life Sci 2010, 67, 3073–3087. [Google Scholar]
- Webster, B.R.; Lu, Z.; Sack, M.N.; Scott, I. The role of sirtuins in modulating redox stressors. Free Radic. Biol. Med 2012, 52, 281–290. [Google Scholar]
- Ota, H.; Eto, M.; Ogawa, S.; Iijima, K.; Akishita, M.; Ouchi, Y. SIRT1/eNOS axis as a potential target against vascular senescence, dysfunction and atherosclerosis. J. Atheroscler. Thromb 2010, 17, 431–435. [Google Scholar]
- Kelly, G.S. A review of the sirtuin system, its clinical implications, and the potential role of dietary activators like resveratrol: Part 2. Altern. Med. Rev 2010, 15, 313–328. [Google Scholar]
- Le Brocq, M.; Leslie, S.J.; Milliken, P.; Megson, I.L. Endothelial dysfunction: From molecular mechanisms to measurement, clinical implications, and therapeutic opportunities. Antioxid. Redox Signal 2008, 10, 1631–1674. [Google Scholar]
- Mattagajasingh, I.; Kim, C.S.; Naqvi, A.; Yamamori, T.; Hoffman, T.A.; Jung, S.B.; DeRicco, J.; Kasuno, K.; Irani, K. SIRT1 promotes endothelium-dependent vascular relaxation by activating endothelial nitric oxide synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 14855–14860. [Google Scholar]
- Yang, L.; Zhang, J.; Yan, C.; Zhou, J.; Lin, R.; Lin, Q.; Wang, W.; Zhang, K.; Yang, G.; Bian, X.; et al. SIRT1 Regulates CD40 Expression Induced by TNF-alpha via NF-κB Pathway in Endothelial Cells. Cell Physiol. Biochem 2012, 30, 1287–1298. [Google Scholar]
- Xia, L.; Ding, F.; Zhu, J.H.; Fu, G.S. Resveratrol attenuates apoptosis of pulmonary microvascular endothelial cells induced by high shear stress and proinflammatory factors. Hum. Cell 2011, 24, 127–133. [Google Scholar]
- Ota, H.; Akishita, M.; Eto, M.; Iijima, K.; Kaneki, M.; Ouchi, Y. Sirt1 modulates premature senescence-like phenotype in human endothelial cells. J. Mol. Cell Cardiol 2007, 43, 571–579. [Google Scholar]
- Zu, Y.; Liu, L.; Lee, M.Y.; Xu, C.; Liang, Y.; Man, R.Y.; Vanhoutte, P.M.; Wang, Y. SIRT1 promotes proliferation and prevents senescence through targeting LKB1 in primary porcine aortic endothelial cells. Circ. Res 2010, 106, 1384–1393. [Google Scholar]
- Csiszar, A.; Labinskyy, N.; Podlutsky, A.; Kaminski, P.M.; Wolin, M.S.; Zhang, C.; Mukhopadhyay, P.; Pacher, P.; Hu, F.; de Cabo, R.; et al. Vasoprotective effects of resveratrol and SIRT1: Attenuation of cigarette smoke-induced oxidative stress and proinflammatory phenotypic alterations. Am. J. Physiol. Heart Circ. Physiol 2008, 294, H2721–H2735. [Google Scholar]
- Ota, H.; Eto, M.; Kano, M.R.; Ogawa, S.; Iijima, K.; Akishita, M.; Ouchi, Y. Cilostazol inhibits oxidative stress-induced premature senescence via upregulation of Sirt1 in human endothelial cells. Arterioscler. Thromb. Vasc. Biol 2008, 28, 1634–1639. [Google Scholar]
- Hou, J.; Wang, S.; Shang, Y.C.; Chong, Z.Z.; Maiese, K. Erythropoietin employs cell longevity pathways of SIRT1 to foster endothelial vascular integrity during oxidant stress. Curr. Neurovasc. Res 2011, 8, 220–235. [Google Scholar]
- Stein, S.; Schafer, N.; Breitenstein, A.; Besler, C.; Winnik, S.; Lohmann, C.; Heinrich, K.; Brokopp, C.E.; Handschin, C.; Landmesser, U.; et al. SIRT1 reduces endothelial activation without affecting vascular function in ApoE−/− mice. Aging (Albany N. Y.) 2010, 2, 353–360. [Google Scholar]
- Zhou, S.; Chen, H.Z.; Wan, Y.Z.; Zhang, Q.J.; Wei, Y.S.; Huang, S.; Liu, J.J.; Lu, Y.B.; Zhang, Z.Q.; Yang, R.F.; et al. Repression of P66Shc expression by SIRT1 contributes to the prevention of hyperglycemia-induced endothelial dysfunction. Circ. Res 2011, 109, 639–648. [Google Scholar]
- Nie, H.; Chen, H.; Han, J.; Hong, Y.; Ma, Y.; Xia, W.; Ying, W. Silencing of SIRT2 induces cell death and a decrease in the intracellular ATP level of PC12 cells. Int. J. Physiol. Pathophysiol. Pharmacol 2011, 3, 65–70. [Google Scholar]
- He, X.; Nie, H.; Hong, Y.; Sheng, C.; Xia, W.; Ying, W. SIRT2 activity is required for the survival of C6 glioma cells. Biochem. Biophys. Res. Commun 2011, 417, 468–472. [Google Scholar]
- Li, Y.; Matsumori, H.; Nakayama, Y.; Osaki, M.; Kojima, H.; Kurimasa, A.; Ito, H.; Mori, S.; Katoh, M.; Oshimura, M.; et al. SIRT2 down-regulation in HeLa can induce p53 accumulation via p38 MAPK activation-dependent p300 decrease, eventually leading to apoptosis. Genes Cells 2011, 16, 34–45. [Google Scholar]
- Liu, P.Y.; Xu, N.; Malyukova, A.; Scarlett, C.J.; Sun, Y.T.; Zhang, X.D.; Ling, D.; Su, S.P.; Nelson, C.; Chang, D.K.; et al. The histone deacetylase SIRT2 stabilizes Myc oncoproteins. Cell Death Differ 2013, 20, 503–514. [Google Scholar]
- Wang, F.; Nguyen, M.; Qin, F.X.; Tong, Q. SIRT2 deacetylates FOXO3a in response to oxidative stress and caloric restriction. Aging Cell 2007, 6, 505–514. [Google Scholar]
- Lynn, E.G.; McLeod, C.J.; Gordon, J.P.; Bao, J.; Sack, M.N. SIRT2 is a negative regulator of anoxia-reoxygenation tolerance via regulation of 14-3-3 zeta and BAD in H9c2 cells. FEBS Lett 2008, 582, 2857–2862. [Google Scholar]
- Liu, L.; Arun, A.; Ellis, L.; Peritore, C.; Donmez, G. Sirtuin 2 (SIRT2) enhances 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced nigrostriatal damage via deacetylating forkhead box O3a (Foxo3a) and activating Bim protein. J. Biol. Chem 2012, 287, 32307–32311. [Google Scholar]
- Luthi-Carter, R.; Taylor, D.M.; Pallos, J.; Lambert, E.; Amore, A.; Parker, A.; Moffitt, H.; Smith, D.L.; Runne, H.; Gokce, O.; et al. SIRT2 inhibition achieves neuroprotection by decreasing sterol biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 7927–7932. [Google Scholar]
- Zhang, Y.; Au, Q.; Zhang, M.; Barber, J.R.; Ng, S.C.; Zhang, B. Identification of a small molecule SIRT2 inhibitor with selective tumor cytotoxicity. Biochem. Biophys. Res. Commun 2009, 386, 729–733. [Google Scholar]
- Rothgiesser, K.M.; Erener, S.; Waibel, S.; Luscher, B.; Hottiger, M.O. SIRT2 regulates NF-kappaB dependent gene expression through deacetylation of p65 Lys310. J. Cell Sci 2010, 123, 4251–4258. [Google Scholar]
- Das, C.; Lucia, M.S.; Hansen, K.C.; Tyler, J.K. CBP/p300-mediated acetylation of histone H3 on lysine 56. Nature 2009, 459, 113–117. [Google Scholar]
- Black, J.C.; Mosley, A.; Kitada, T.; Washburn, M.; Carey, M. The SIRT2 deacetylase regulates autoacetylation of p300. Mol. Cell 2008, 32, 449–455. [Google Scholar]
- Bouras, T.; Fu, M.; Sauve, A.A.; Wang, F.; Quong, A.A.; Perkins, N.D.; Hay, R.T.; Gu, W.; Pestell, R.G. SIRT1 deacetylation and repression of p300 involves lysine residues 1020/1024 within the cell cycle regulatory domain 1. J. Biol. Chem 2005, 280, 10264–10276. [Google Scholar]
- Coussens, M.; Maresh, J.G.; Yanagimachi, R.; Maeda, G.; Allsopp, R. Sirt1 deficiency attenuates spermatogenesis and germ cell function. PLoS One 2008, 3, e1571. [Google Scholar]
- Potente, M.; Ghaeni, L.; Baldessari, D.; Mostoslavsky, R.; Rossig, L.; Dequiedt, F.; Haendeler, J.; Mione, M.; Dejana, E.; Alt, F.W.; et al. SIRT1 controls endothelial angiogenic functions during vascular growth. Genes Dev 2007, 21, 2644–2658. [Google Scholar]
- Outeiro, T.F.; Kontopoulos, E.; Altmann, S.M.; Kufareva, I.; Strathearn, K.E.; Amore, A.M.; Volk, C.B.; Maxwell, M.M.; Rochet, J.C.; McLean, P.J.; et al. Sirtuin 2 inhibitors rescue alpha-synuclein-mediated toxicity in models of Parkinson’s disease. Science 2007, 317, 516–519. [Google Scholar]
- De Oliveira, R.M.; Sarkander, J.; Kazantsev, A.G.; Outeiro, T.F. SIRT2 as a Therapeutic Target for Age-Related Disorders. Front Pharmacol 2012, 3, 82. [Google Scholar]
- Gal, J.; Bang, Y.; Choi, H.J. SIRT2 interferes with autophagy-mediated degradation of protein aggregates in neuronal cells under proteasome inhibition. Neurochem. Int 2012, 61, 992–1000. [Google Scholar]
- Liu, J.; Jiang, F. Shandong University, Jinan, Shandong Province, China. Personal communication, 2012. [Google Scholar]
- Pfister, J.A.; Ma, C.; Morrison, B.E.; D’Mello, S.R. Opposing effects of sirtuins on neuronal survival: SIRT1-mediated neuroprotection is independent of its deacetylase activity. PLoS One 2008, 3, e4090. [Google Scholar]
- Hashimoto-Komatsu, A.; Hirase, T.; Asaka, M.; Node, K. Angiotensin II induces microtubule reorganization mediated by a deacetylase SIRT2 in endothelial cells. Hypertens Res 2011, 34, 949–956. [Google Scholar]
- Numaguchi, Y.; Huang, S.; Polte, T.R.; Eichler, G.S.; Wang, N.; Ingber, D.E. Caldesmon-dependent switching between capillary endothelial cell growth and apoptosis through modulation of cell shape and contractility. Angiogenesis 2003, 6, 55–64. [Google Scholar]
- Tang, J.; Hu, G.; Hanai, J.; Yadlapalli, G.; Lin, Y.; Zhang, B.; Galloway, J.; Bahary, N.; Sinha, S.; Thisse, B.; et al. A critical role for calponin 2 in vascular development. J. Biol. Chem 2006, 281, 6664–6672. [Google Scholar]
- Dennis, M.D.; Baum, J.I.; Kimball, S.R.; Jefferson, L.S. Mechanisms involved in the coordinate regulation of mTORC1 by insulin and amino acids. J. Biol. Chem 2011, 286, 8287–8296. [Google Scholar]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc 2009, 4, 44–57. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sirt1 | ||
|---|---|---|
| 11716338_a_at | U | INSIG1 |
| 11716395_a_at | U | GPR56 |
| 11718915_a_at | U | RGS19 |
| 11719218_at | U | SOCS3 |
| 11719513_a_at | U | ADAM15 |
| 11719745_s_at | U | ARHGAP27 |
| 11720832_x_at | U | SOX18 |
| 11722324_a_at | U | ZNF84 |
| 11722353_s_at | U | LDB2 |
| 11724390_x_at | U | ZDHHC16 |
| 11724394_at | U | C2orf55 |
| 11724395_a_at | U | ZNF83 |
| 11724396_x_at | U | ZNF83 |
| 11727984_at | U | ODZ3 |
| 11728347_at | U | ABI3 |
| 11729918_at | U | ADAMTS18 |
| 11729919_a_at | U | ADAMTS18 |
| 11730211_x_at | U | PFKL |
| 11731622_x_at | U | ZNF91 |
| 11733299_a_at | U | CLDN5 |
| 11736013_at | U | GLE1 |
| 11736029_a_at | U | ITGA6 |
| 11736458_x_at | U | KCNK6 |
| 11737089_a_at | U | TLL1 |
| 11737870_s_at | U | FAM78A |
| 11739146_a_at | U | MKNK2 |
| 11739491_a_at | U | MGAT4A |
| 11739492_a_at | U | MGAT4A |
| 11739493_at | U | MGAT4A |
| 11739650_at | U | DLL4 |
| 11740601_a_at | U | APH1B |
| 11722277_s_at | U | ZBED4 |
| 11722388_at | U | MTF1 |
| 11722391_at | U | MTF1 |
| 11722531_a_at | U | CALD1 |
| 11722532_s_at | U | CALD1 |
| 11722533_x_at | U | CALD1 |
| 11723092_at | U | FNIP2 |
| 11723414_a_at | U | CNN2 |
| 11723416_x_at | U | CNN2 |
| 11724389_a_at | U | ZDHHC16 |
| 11725054_a_at | U | HNMT |
| 11726328_x_at | U | GBP1 |
| 11726796_a_at | U | NEXN |
| 11726797_x_at | U | NEXN |
| 11726824_a_at | U | ZNF565 |
| 11727545_at | U | PANK3 |
| 11727784_x_at | U | TPM4 |
| 11728028_a_at | U | PWWP2A |
| 11728226_a_at | U | CASP7 |
| 11728276_s_at | U | PLA2G12A |
| 11728488_a_at | U | NRXN3 |
| 11728744_at | U | C4orf46 |
| 11729665_a_at | U | STK38L |
| 11730613_at | U | MBOAT7 |
| 11731665_a_at | U | PDGFB |
| 11732339_at | U | BCL11A |
| 11733043_a_at | U | SLC7A2 |
| 11733084_a_at | U | PALLD |
| 11734281_a_at | U | ZNF17 |
| 11734865_a_at | U | PSMC3IP |
| 11736812_at | U | PDCD1LG2 |
| 11736959_a_at | U | TNFSF15 |
| 11724786_s_at | D | C14orf129 |
| 11725510_a_at | D | SLC7A6 |
| 11725569_at | D | KLHL15 |
| 11725596_a_at | D | ZDHHC23 |
| 11725733_a_at | D | NFYA |
| 11725850_at | D | USP48 |
| 11726455_x_at | D | MYO5B |
| 11726551_s_at | D | RAET1L |
| 11726552_x_at | D | ULBP2 |
| 11727222_at | D | EVI5 |
| 11727286_a_at | D | ZNF323 |
| 11727361_a_at | D | MYLK |
| 11727406_a_at | D | TEK |
| 11727485_at | D | TPR |
| 11727854_s_at | D | NUP50 |
| 11727905_a_at | D | IL13RA1 |
| 11727984_at | D | ODZ3 |
| 11728195_s_at | D | TRO |
| 11728288_a_at | D | KRT15 |
| 11728353_at | D | MSMP |
| 11728960_a_at | D | KRT80 |
| 11729449_s_at | D | TINF2 |
| 11729550_a_at | D | GPR126 |
| 11729840_s_at | D | ZCCHC2 |
| 11730033_a_at | D | RPS6KL1 |
| 11730195_at | D | SEC14L2 |
| 11730615_a_at | D | PLCD4 |
| 11730616_at | D | PLCD4 |
| 11730623_at | D | DSEL |
| 11732160_a_at | D | NUDCD1 |
| 11732713_at | D | FST |
| 11733051_a_at | D | SCAMP1 |
| 11740602_s_at | U | APH1B |
| 11740624_a_at | U | AZI1 |
| 11742830_a_at | U | PHF3 |
| 11744239_a_at | U | CCDC159 |
| 11744430_a_at | U | KIF9 |
| 11744505_x_at | U | TP53I11 |
| 11744948_x_at | U | SEMA3F |
| 11745450_a_at | U | AUH |
| 11745927_x_at | U | ADAM15 |
| 11746516_a_at | U | ADAM15 |
| 11747060_a_at | U | MGAT4A |
| 11747977_a_at | U | ZNF180 |
| 11748731_a_at | U | ZNF616 |
| 11749511_a_at | U | ZNF595 |
| 11751946_a_at | U | ARHGAP21 |
| 11752002_a_at | U | PLEKHG1 |
| 11752675_a_at | U | OGFR |
| 11754022_s_at | U | GAP43 |
| 11754446_x_at | U | ZNF761 |
| 11754754_s_at | U | ADAMTS18 |
| 11755219_a_at | U | THBD |
| 11755232_s_at | U | ZNF468 |
| 11755474_a_at | U | ADAM15 |
| 11757861_a_at | U | TSHZ1 |
| 11760814_x_at | U | KANK3 |
| 11715679_s_at | D | PRNP |
| 11715889_a_at | D | ATP5I |
| 11715890_x_at | D | ATP5I |
| 11716027_at | D | SELT |
| 11716028_x_at | D | SELT |
| 11716404_s_at | D | EIF4EBP2 |
| 11718102_at | D | CD59 |
| 11718141_at | D | DKK1 |
| 11718769_a_at | D | MAPKSP1 |
| 11719164_a_at | D | CLCN5 |
| 11719267_s_at | D | SERINC1 |
| 11719394_a_at | D | FBXO32 |
| 11719395_at | D | EXTL2 |
| 11719396_a_at | D | EXTL2 |
| 11719479_at | D | ALOX5AP |
| 11719712_at | D | PPM1B |
| 11719816_s_at | D | BET1 |
| 11720240_at | D | TMSB15A |
| 11720514_at | D | C9orf150 |
| 11721024_a_at | D | IL11RA |
| 11721112_a_at | D | ACP6 |
| 11739010_a_at | U | MYH9 |
| 11739086_x_at | U | MESDC2 |
| 11739088_at | U | MESDC2 |
| 11739451_a_at | U | FRMD6 |
| 11739672_x_at | U | ZNF253 |
| 11740133_a_at | U | CALD1 |
| 11740743_a_at | U | TPM2 |
| 11740744_x_at | U | TPM2 |
| 11741168_a_at | U | MSRB3 |
| 11741188_a_at | U | SLC30A6 |
| 11742483_a_at | U | C1orf110 |
| 11743015_a_at | U | DIP2C |
| 11743020_at | U | ZSCAN29 |
| 11743253_x_at | U | CALD1 |
| 11743458_a_at | U | FAM49A |
| 11743696_at | U | CLEC14A |
| 11743705_at | U | ETAA1 |
| 11744034_a_at | U | VASP |
| 11744323_s_at | U | PWWP2A |
| 11745608_a_at | U | WDR1 |
| 11745924_at | U | LOC220930 |
| 11746173_a_at | U | DDHD1 |
| 11746476_x_at | U | CALD1 |
| 11746548_s_at | U | CNN2 |
| 11747300_a_at | U | CDK14 |
| 11747469_x_at | U | KDM2B |
| 11747711_a_at | U | LDB1 |
| 11748208_a_at | U | ZNF317 |
| 11748391_x_at | U | ZDHHC16 |
| 11748400_s_at | U | LOC643634 |
| 11748401_x_at | U | TPM4 |
| 11748403_x_at | U | CNN2 |
| 11748527_a_at | U | ARHGAP24 |
| 11749172_x_at | U | NEXN |
| 11749732_a_at | U | LONP2 |
| 11749734_s_at | U | JRKL |
| 11749921_a_at | U | SDC4 |
| 11750198_a_at | U | CASP7 |
| 11750623_a_at | U | FILIP1L |
| 11751244_s_at | U | CNN2 |
| 11751245_x_at | U | CNN2 |
| 11751993_a_at | U | JAG1 |
| 11752164_x_at | U | KDM2B |
| 11752276_a_at | U | DIDO1 |
| 11752361_s_at | U | NEXN |
| 11752499_a_at | U | CALD1 |
| 11733054_a_at | D | SCAMP1 |
| 11733929_a_at | D | ARMC8 |
| 11734059_a_at | D | PSTK |
| 11734150_x_at | D | PLAGL1 |
| 11735224_a_at | D | KLRG1 |
| 11735991_at | D | LARS |
| 11736192_at | D | RRM2B |
| 11736343_x_at | D | OPA3 |
| 11736345_x_at | D | OPA3 |
| 11736528_a_at | D | SMC2 |
| 11736785_at | D | HOXA2 |
| 11737052_x_at | D | PLAGL1 |
| 11737816_x_at | D | FAM119B |
| 11739064_s_at | D | GNG12 |
| 11739245_a_at | D | ANKFY1 |
| 11739596_a_at | D | KIAA1429 |
| 11739640_at | D | DIP2A |
| 11739942_s_at | D | SEPSECS |
| 11740096_a_at | D | TMCC1 |
| 11740176_at | D | ARSK |
| 11740213_a_at | D | TBL1X |
| 11741152_x_at | D | PLAGL1 |
| 11742720_at | D | LRRC58 |
| 11742722_at | D | LRRC58 |
| 11742962_a_at | D | IP6K2 |
| 11743404_at | D | ZMAT2 |
| 11743573_at | D | TMEM184C |
| 11743574_x_at | D | TMEM184C |
| 11743648_a_at | D | DCAF6 |
| 11743649_a_at | D | DCAF6 |
| 11743763_at | D | GTF3C3 |
| 11744083_at | D | ANKIB1 |
| 11744415_s_at | D | MFSD6 |
| 11744788_x_at | D | TMEM68 |
| 11745010_a_at | D | DCBLD2 |
| 11745230_a_at | D | C3orf23 |
| 11745231_a_at | D | C3orf23 |
| 11745700_s_at | D | ULBP2 |
| 11746163_a_at | D | LARP4B |
| 11746536_a_at | D | WSB2 |
| 11747146_s_at | D | TMBIM6 |
| 11748416_a_at | D | DCAF6 |
| 11749027_x_at | D | HERPUD2 |
| 11750354_a_at | D | TMEM184C |
| 11750993_x_at | D | MAP3K7 |
| 11751165_a_at | D | RBMS2 |
| 11722843_a_at | D | ENAH |
| 11723533_x_at | D | BRMS1L |
| 11723534_at | D | BRMS1L |
| 11723580_at | D | LOC221710 |
| 11724238_at | D | CYB5R4 |
| 11726140_s_at | D | SIRT1 |
| 11726750_a_at | D | GTF3C4 |
| 11727022_at | D | TMEM64 |
| 11727370_at | D | TSN |
| 11727935_at | D | C4orf49 |
| 11729128_at | D | CPA4 |
| 11729710_a_at | D | MARS2 |
| 11731195_at | D | SEMA3D |
| 11731263_a_at | D | ZNF365 |
| 11734371_a_at | D | SCML1 |
| 11734955_a_at | D | SCML1 |
| 11736470_at | D | SLC35D1 |
| 11738893_s_at | D | TPM1 |
| 11739119_s_at | D | CNPY3 |
| 11742308_s_at | D | TPM1 |
| 11742483_a_at | D | C1orf110 |
| 11742743_a_at | D | CNN3 |
| 11743092_at | D | THEM4 |
| 11743197_at | D | TLR4 |
| 11743334_a_at | D | MRPL35 |
| 11743973_a_at | D | MRPL38 |
| 11743974_at | D | MRPL38 |
| 11745482_s_at | D | PRNP |
| 11746622_a_at | D | PHC1 |
| 11746928_a_at | D | ENC1 |
| 11747834_a_at | D | SPATA5 |
| 11748315_s_at | D | PRNP |
| 11750985_a_at | D | EXTL2 |
| 11751191_a_at | D | LIPG |
| 11752333_a_at | D | ITGAV |
| 11754940_s_at | D | TSN |
| 11754976_x_at | D | CNPY3 |
| 11755458_a_at | D | HDDC2 |
| 11755848_a_at | D | C17orf51 |
| 11756471_a_at | D | MFSD2A |
| 11756882_a_at | D | RTTN |
| 11757738_s_at | D | FAT1 |
| 11758013_s_at | D | C8orf4 |
| 11758101_s_at | D | EIF4EBP2 |
| 11758326_s_at | D | THEM4 |
| 11758872_at | D | CDC37L1 |
| 11752930_a_at | U | GBP1 |
| 11754084_x_at | U | MYL9 |
| 11754644_x_at | U | CNN2 |
| 11754887_a_at | U | MSRB3 |
| 11754911_x_at | U | NEXN |
| 11755122_a_at | U | PALLD |
| 11755734_x_at | U | CCDC107 |
| 11757637_a_at | U | MUS81 |
| 11758013_s_at | U | C8orf4 |
| 11759169_a_at | U | C1orf222 |
| 11759711_a_at | U | RBM25 |
| 11760202_at | U | IGFBP7 |
| 11760611_x_at | U | SETD6 |
| 11760918_a_at | U | MRRF |
| 11763500_a_at | U | ZNF93 |
| 11715207_at | D | WDFY4 |
| 11715265_at | D | FIG4 |
| 11715477_at | D | TFRC |
| 11715545_at | D | TMED10 |
| 11715550_at | D | DAG1 |
| 11715651_s_at | D | FSTL1 |
| 11715761_a_at | D | TBC1D14 |
| 11716015_a_at | D | CMTM3 |
| 11716208_s_at | D | GLUD1 |
| 11716288_s_at | D | ESYT1 |
| 11716391_a_at | D | BACE1 |
| 11716620_a_at | D | TNPO1 |
| 11716626_at | D | KIF3B |
| 11716787_a_at | D | B3GNT1 |
| 11716788_at | D | B3GNT1 |
| 11718184_a_at | D | FCHSD2 |
| 11718287_at | D | UBL3 |
| 11718288_at | D | UBL3 |
| 11718406_s_at | D | TMBIM6 |
| 11718439_at | D | NSUN4 |
| 11718734_a_at | D | POGK |
| 11718900_a_at | D | TGFBR3 |
| 11718901_at | D | TGFBR3 |
| 11719088_at | D | MMGT1 |
| 11719353_s_at | D | GCC2 |
| 11719397_a_at | D | RRAGC |
| 11719398_s_at | D | RRAGC |
| 11719409_a_at | D | HIPK2 |
| 11719628_a_at | D | HDHD1A |
| 11719786_at | D | SMCR7L |
| 11719912_a_at | D | KDM1B |
| 11751297_s_at | D | SUB1 |
| 11751305_a_at | D | B3GNT1 |
| 11751353_a_at | D | RRAGC |
| 11751354_a_at | D | HDHD1A |
| 11753308_s_at | D | ULBP2 |
| 11753549_a_at | D | CMTM3 |
| 11754031_s_at | D | CKS1B |
| 11754827_x_at | D | FBXO17 |
| 11755251_x_at | D | FADS1 |
| 11756152_s_at | D | PCNP |
| 11756156_s_at | D | TFRC |
| 11756181_x_at | D | YWHAZ |
| 11756205_x_at | D | DCAF6 |
| 11756254_a_at | D | GGT5 |
| 11756259_s_at | D | NFATC1 |
| 11756285_s_at | D | IGF2BP3 |
| 11756497_a_at | D | VWCE |
| 11756603_a_at | D | C9orf6 |
| 11756861_s_at | D | ULBP2 |
| 11757430_s_at | D | TMED5 |
| 11757523_s_at | D | WDR35 |
| 11757542_s_at | D | SSR1 |
| 11757787_x_at | D | FTL |
| 11757799_s_at | D | VAMP7 |
| 11757810_s_at | D | TMED10 |
| 11757880_s_at | D | DAG1 |
| 11757989_s_at | D | ANKRD46 |
| 11758200_x_at | D | CKS1B |
| 11758212_s_at | D | KIAA0494 |
| 11758452_s_at | D | CENPQ |
| 11758454_s_at | D | FAM116A |
| 11758750_x_at | D | YWHAZ |
| 11758751_at | D | YWHAZ |
| 11758809_at | D | RRAGC |
| 11758820_at | D | DNAJC10 |
| 11758873_a_at | D | HPSE |
| 11758995_at | D | LOX |
| 11758999_s_at | D | UBE2H |
| 11759720_x_at | D | LOC400590 |
| 11763276_a_at | D | PWP1 |
| 11763395_a_at | D | ZC3HAV1L |
| 11764275_s_at | D | SLC11A2 |
| Co-regulated | ||
| 11715698_a_at | U | NOLC1 |
| 11717961_at | U | MED11 |
| 11718908_s_at | U | CHST2 |
| 11758929_at | D | TFDP2 |
| 11759503_at | D | FAM103A1 |
| 11759776_at | D | GTDC1 |
| 11759943_at | D | FAM13A |
| Sirt2 | ||
| 11715270_s_at | U | KLF7 |
| 11715586_at | U | MAPRE1 |
| 11715713_a_at | U | VOPP1 |
| 11715803_a_at | U | ANXA6 |
| 11716299_a_at | U | ITGAV |
| 11716344_a_at | U | ZCCHC3 |
| 11716413_x_at | U | MT1E |
| 11716582_a_at | U | G3BP2 |
| 11716771_s_at | U | SIK1 |
| 11717055_a_at | U | CORO1C |
| 11717056_a_at | U | CORO1C |
| 11717952_at | U | ZDHHC18 |
| 11717968_at | U | TMEM216 |
| 11719647_a_at | U | CASP7 |
| 11719648_a_at | U | CASP7 |
| 11719833_at | U | MPZL2 |
| 11720028_x_at | U | LDLR |
| 11720063_a_at | U | GLIPR2 |
| 11720223_at | U | GFPT1 |
| 11720823_at | U | JAG1 |
| 11720849_a_at | U | RAB23 |
| 11721218_a_at | U | MSRB3 |
| 11721525_s_at | U | LOC440354 |
| 11721684_a_at | U | ZW10 |
| 11721778_a_at | U | ACBD5 |
| 11722193_a_at | U | C12orf75 |
| 11722218_a_at | U | WBP4 |
| 11722220_a_at | U | WBP4 |
| 11720046_x_at | D | DNAJC10 |
| 11720111_at | D | SNTB2 |
| 11720112_at | D | SNTB2 |
| 11720146_a_at | D | DAPK1 |
| 11720273_at | D | SFT2D3 |
| 11720570_a_at | D | PHF15 |
| 11720599_s_at | D | SUB1 |
| 11720602_at | D | SYT11 |
| 11720798_at | D | RAB8B |
| 11720799_s_at | D | RAB8B |
| 11720800_a_at | D | RAB8B |
| 11720893_s_at | D | SOS1 |
| 11721524_s_at | D | ZNF706 |
| 11721585_a_at | D | TMCC1 |
| 11721622_a_at | D | KATNAL1 |
| 11721834_a_at | D | GET4 |
| 11721993_at | D | SLC6A6 |
| 11722111_at | D | HLX |
| 11722273_s_at | D | KIAA1826 |
| 11722338_at | D | PEX7 |
| 11722377_at | D | PNPO |
| 11722425_s_at | D | NEDD4L |
| 11722460_at | D | PCDH12 |
| 11722475_a_at | D | ARID3B |
| 11722662_a_at | D | HPSE |
| 11722969_s_at | D | TRPV4 |
| 11722977_at | D | HOXB5 |
| 11723228_s_at | D | SGCB |
| 11723230_a_at | D | RNF138 |
| 11723586_a_at | D | GRB14 |
| 11723639_s_at | D | C11orf58 |
| 11724011_at | D | SIKE1 |
| 11724171_a_at | D | FCHO2 |
| 11718909_x_at | U | CHST2 |
| 11721860_s_at | U | STX12 |
| 11721862_a_at | U | STX12 |
| 11722853_a_at | U | HABP4 |
| 11724388_at | U | ZNF721 |
| 11724959_s_at | U | CDK14 |
| 11727522_a_at | U | ZNF267 |
| 11727523_x_at | U | ZNF267 |
| 11727782_a_at | U | TPM4 |
| 11727783_s_at | U | TPM4 |
| 11728155_s_at | U | FUT4 |
| 11731868_a_at | U | BICD2 |
| 11734554_a_at | U | BCL7B |
| 11738988_a_at | U | GANAB |
| 11738989_a_at | U | GANAB |
| 11738990_x_at | U | GANAB |
| 11743449_a_at | U | BICD2 |
| 11744786_x_at | U | OGFR |
| 11748926_a_at | U | GANAB |
| 11752163_a_at | U | KDM2B |
| 11752676_x_at | U | OGFR |
| 11753089_a_at | U | OGFR |
| 11753090_x_at | U | OGFR |
| 11754869_s_at | U | ZNF267 |
| 11755270_a_at | U | GANAB |
| 11757525_s_at | U | BICD2 |
| 11759004_at | U | SLC33A1 |
| 11759757_a_at | U | SLC33A1 |
| 11741758_x_at | D | TRPV4 |
| 11754398_at | D | LOC644538 |
| 11758147_s_at | D | MAPK13 |
| 11758902_at | D | ZNF641 |
Supplementary Files
{kind=link}
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, J.; Wu, X.; Wang, X.; Zhang, Y.; Bu, P.; Zhang, Q.; Jiang, F. Global Gene Expression Profiling Reveals Functional Importance of Sirt2 in Endothelial Cells under Oxidative Stress. Int. J. Mol. Sci. 2013, 14, 5633-5649. https://doi.org/10.3390/ijms14035633
Liu J, Wu X, Wang X, Zhang Y, Bu P, Zhang Q, Jiang F. Global Gene Expression Profiling Reveals Functional Importance of Sirt2 in Endothelial Cells under Oxidative Stress. International Journal of Molecular Sciences. 2013; 14(3):5633-5649. https://doi.org/10.3390/ijms14035633
Chicago/Turabian StyleLiu, Junni, Xiao Wu, Xi Wang, Yun Zhang, Peili Bu, Qunye Zhang, and Fan Jiang. 2013. "Global Gene Expression Profiling Reveals Functional Importance of Sirt2 in Endothelial Cells under Oxidative Stress" International Journal of Molecular Sciences 14, no. 3: 5633-5649. https://doi.org/10.3390/ijms14035633
APA StyleLiu, J., Wu, X., Wang, X., Zhang, Y., Bu, P., Zhang, Q., & Jiang, F. (2013). Global Gene Expression Profiling Reveals Functional Importance of Sirt2 in Endothelial Cells under Oxidative Stress. International Journal of Molecular Sciences, 14(3), 5633-5649. https://doi.org/10.3390/ijms14035633