Combinatorial Analysis of Secretory Immunoglobulin A (sIgA) Expression in Plants

Abstract
:1. Introduction
2. Results
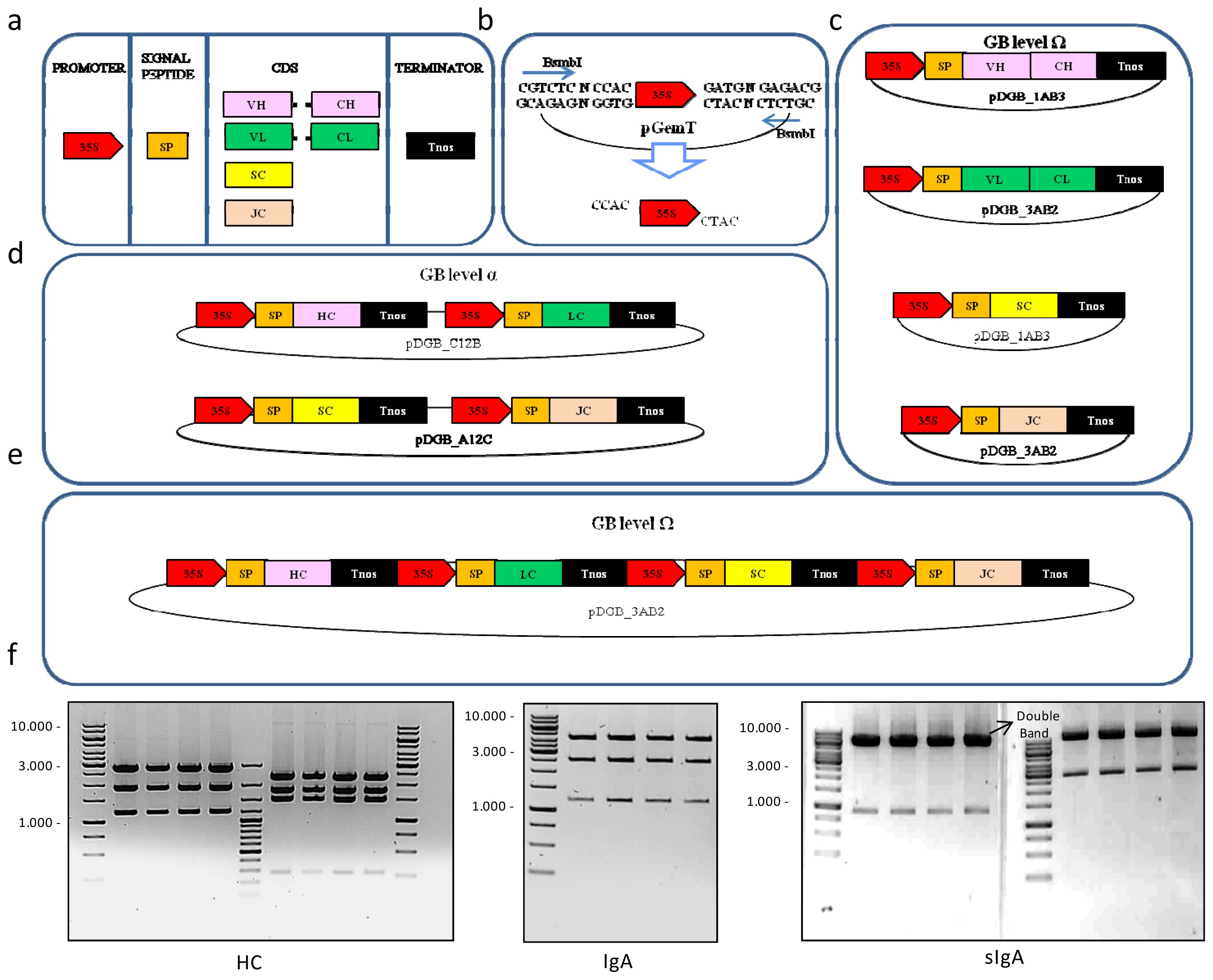
2.1. GoldenBraid-Assisted Multigene Assembly of 16 Versions of Secretory IgA
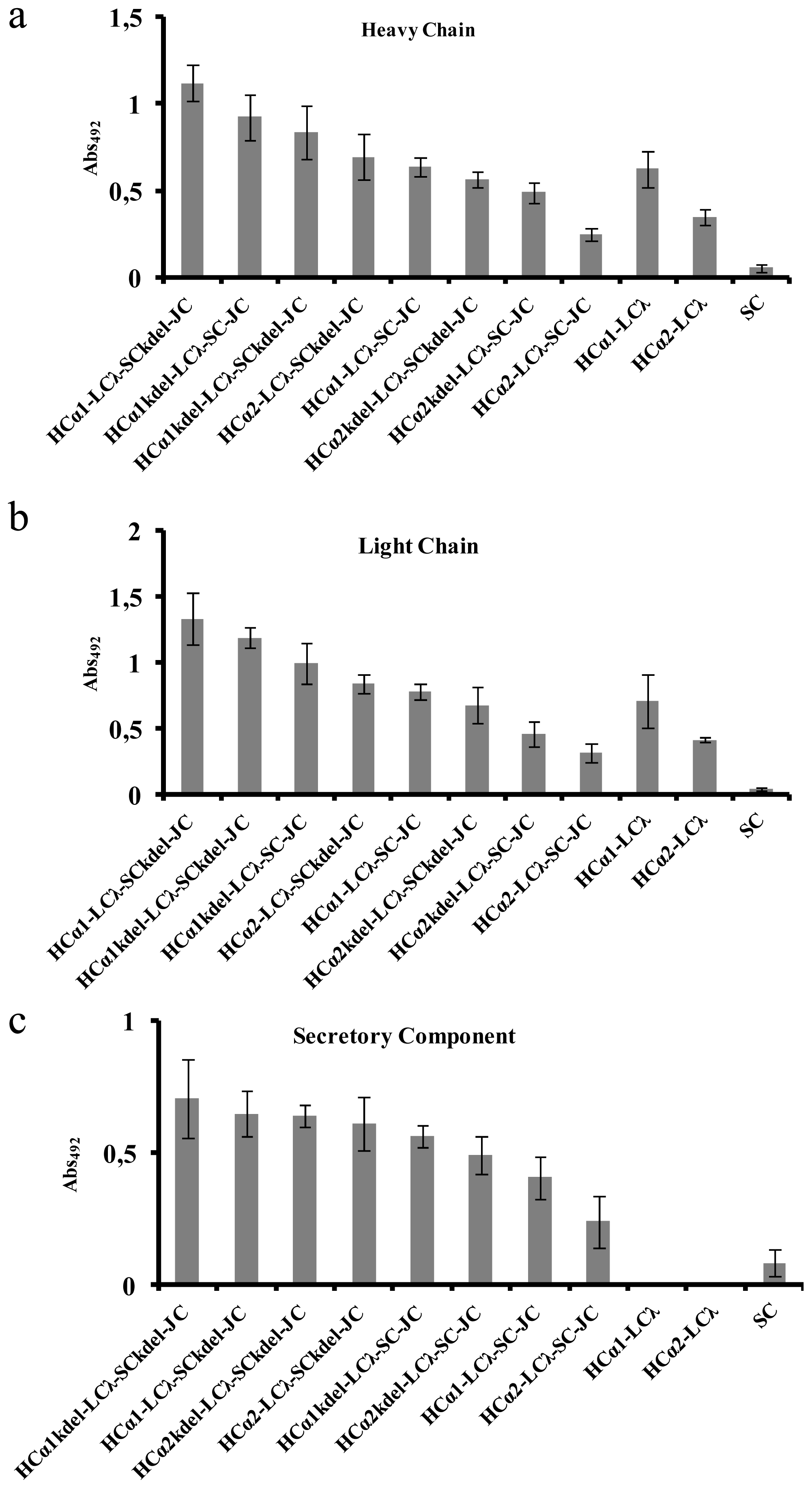
2.2. Transient Expression in Nicotiana benthamiana of 16 Versions of sIgA against Rotavirus
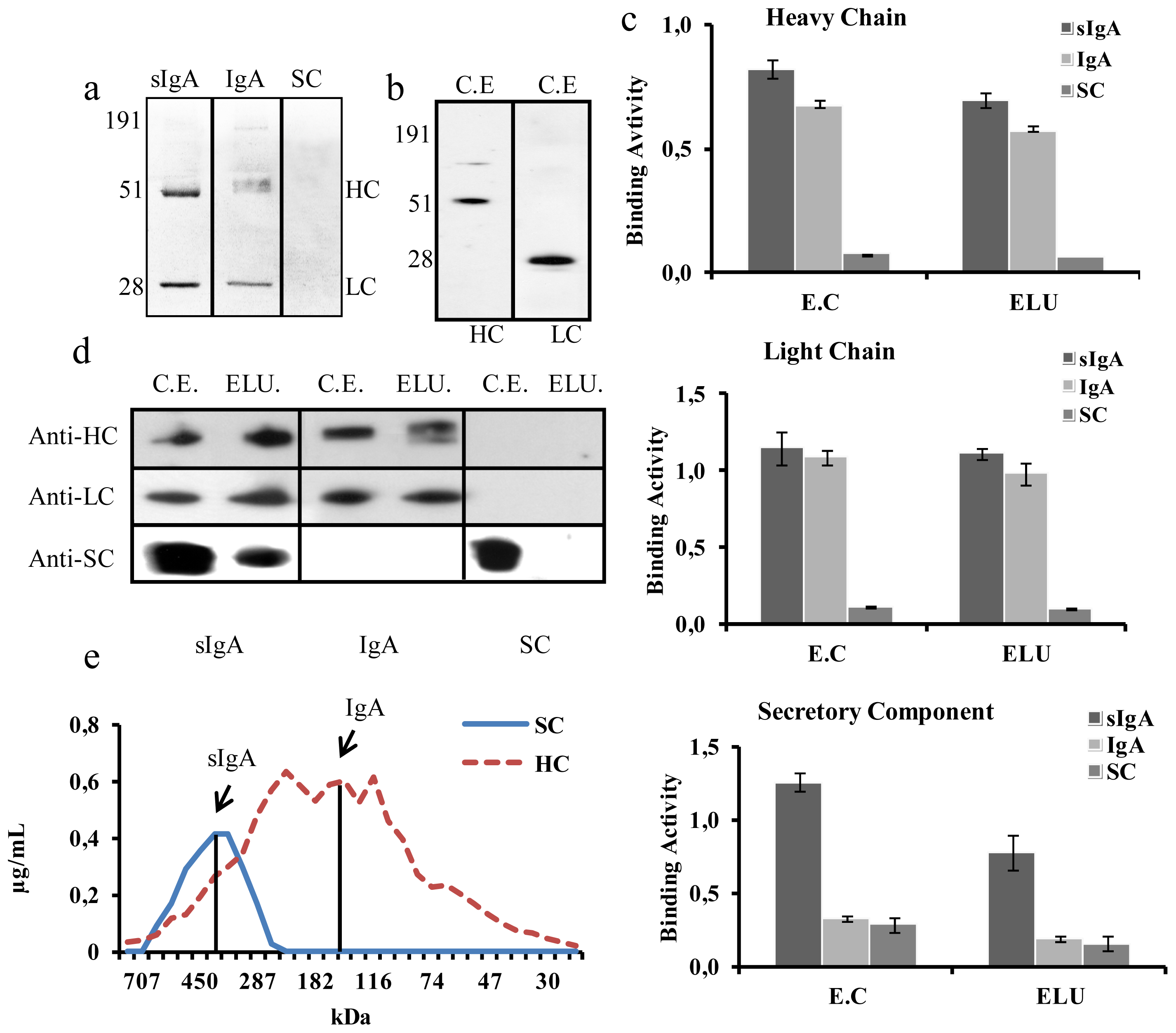
2.3. Detailed Characterization and Purification of the HCα1-LCλ-JC-SCkdel Combination
3. Discussion
4. Experimental Section
4.1. Cloning and Assembly of Modular Pieces
4.2. Strains and Growth Conditions
4.3. Plant Transient Transformation
4.4. Plant Material and Sample Preparation
4.5. VP8* Rotavirus Surface Protein Production
4.6. ELISAs for the Quantification and Detection of VP8* Binding Activity of IgA and sIgA
4.7. SDS-PAGE and Western Blot Analysis
4.8. SSL7 Affinity Purification
5. Conclusions
Supplementary Information
ijms-14-06205-s001.docxAcknowledgments
References
- Reichert, J.M.; Valge-Archer, V.E. Development trends for monoclonal antibody cancer therapeutics. Nat. Rev. Drug Discov 2007, 6, 349–356. [Google Scholar]
- Corthesy, B. Role of secretory immunoglobulin a and secretory component in the protection of mucosal surfaces. Future Microbiol 2010, 5, 817–829. [Google Scholar]
- Corthesy, B. Recombinant secretory immunoglobulin a in passive immunotherapy: Linking immunology and biotechnology. Curr. Pharm. Biotechnol 2003, 4, 51–67. [Google Scholar]
- Corthesy, B. Secretory immunoglobulin a: Well beyond immune exclusion at mucosal surfaces. Immunopharmacol. Immunotoxicol 2009, 31, 174–179. [Google Scholar]
- Sarrion-Perdigones, A.; Juarez, P.; Granell, A.; Orzaez, D. Production of antibodies in plants. In Cell Engineering; Al-Rubeai, M., Ed.; Springer Science Business Media: Dublin, Ireland, 2011; Volume 7, pp. 143–164. [Google Scholar]
- Paul, M.; van Dolleweerd, C.; Drake, P.M.; Reljic, R.; Thangaraj, H.; Barbi, T.; Stylianou, E.; Pepponi, I.; Both, L.; Hehle, V.; et al. Molecular pharming: Future targets and aspirations. Hum. Vaccin. 2011, 7, 375–382. [Google Scholar]
- Fischer, R.; Stoger, E.; Schillberg, S.; Christou, P.; Twyman, R.M. Plant-Based production of biopharmaceuticals. Curr. Opin. Plant Biol 2004, 7, 152–158. [Google Scholar]
- Twyman, R.M.; Stoger, E.; Schillberg, S.; Christou, P.; Fischer, R. Molecular farming in plants: Host systems and expression technology. Trends Biotechnol 2003, 21, 570–578. [Google Scholar]
- Saint-Jore-Dupas, C.; Faye, L.; Gomord, V. From planta to pharma with glycosylation in the toolbox. Trends Biotechnol 2007, 25, 317–323. [Google Scholar]
- Ma, J.K.; Hiatt, A.; Hein, M.; Vine, N.D.; Wang, F.; Stabila, P.; van Dolleweerd, C.; Mostov, K.; Lehner, T. Generation and assembly of secretory antibodies in plants. Science 1995, 268, 716–719. [Google Scholar]
- Ma, J.K.; Hikmat, B.Y.; Wycoff, K.; Vine, N.D.; Chargelegue, D.; Yu, L.; Hein, M.B.; Lehner, T. Characterization of a recombinant plant monoclonal secretory antibody and preventive immunotherapy in humans. Nat. Med 1998, 4, 601–606. [Google Scholar]
- Weintraub, J.A.; Hilton, J.F.; White, J.M.; Hoover, C.I.; Wycoff, K.L.; Yu, L.; Larrick, J.W.; Featherstone, J.D. Clinical trial of a plant-derived antibody on recolonization of mutans streptococci. Caries Res 2005, 39, 241–250. [Google Scholar]
- Wycoff, K.L. Secretory IgA antibodies from plants. Curr. Pharm. Des 2004, 11, 2429–2437. [Google Scholar]
- Nicholson, L.; Gonzalez-Melendi, P.; van Dolleweerd, C.; Tuck, H.; Perrin, Y.; Ma, J.K.; Fischer, R.; Christou, P.; Stoger, E. A recombinant multimeric immunoglobulin expressed in rice shows assembly-dependent subcellular localization in endosperm cells. Plant Biotechnol. J 2005, 3, 115–127. [Google Scholar]
- Wieland, W.H.; Lammers, A.; Schots, A.; Orzaez, D.V. Plant expression of chicken secretory antibodies derived from combinatorial libraries. J. Biotechnol 2006, 122, 382–391. [Google Scholar]
- Larrick, J.W.; Yu, L.; Naftzger, C.; Jaiswal, S.; Wycoff, K. Production of secretory IgA antibodies in plants. Biomol. Eng 2001, 18, 87–94. [Google Scholar]
- Foley, R.C.; Raison, R.L.; Beh, K.J. Monoclonal antibody against sheep kappa light chain. Hybridoma 1991, 10, 507–515. [Google Scholar]
- Furtado, P.B.; Whitty, P.W.; Robertson, A.; Eaton, J.T.; Almogren, A.; Kerr, M.A.; Woof, J.M.; Perkins, S.J. Solution structure determination of monomeric human IgA2 by X-ray and neutron scattering, analytical ultracentrifugation and constrained modelling: A comparison with monomeric human IgA1. J. Mol. Biol 2004, 338, 921–941. [Google Scholar]
- De Muynck, B.; Navarre, C.; Boutry, M. Production of antibodies in plants: Status after twenty years. Plant Biotechnol. J 2010, 8, 529–563. [Google Scholar]
- Ma, J.K.C.; Drake, P.M.W.; Christou, P. The production of recombinant pharmaceutical proteins in plants. Nat. Rev. Genet 2003, 4, 794–805. [Google Scholar]
- Petruccelli, S.; Otegui, M.S.; Lareu, F.; Tran Dinh, O.; Fitchette, A.C.; Circosta, A.; Rumbo, M.; Bardor, M.; Carcamo, R.; Gomord, V.; et al. A KDEL-tagged monoclonal antibody is efficiently retained in the endoplasmic reticulum in leaves, but is both partially secreted and sorted to protein storage vacuoles in seeds. Plant Biotechnol. J. 2006, 4, 511–527. [Google Scholar]
- De Muynck, B.; Navarre, C.; Nizet, Y.; Stadlmann, J.; Boutry, M. Different subcellular localization and glycosylation for a functional antibody expressed in Nicotiana tabacum plants and suspension cells. Transgenic Res 2009, 18, 467–482. [Google Scholar]
- Düring, K.; Hippe, S.; Kreuzaler, F.; Schell, J. Synthesis and self-assembly of a functional monoclonal antibody in transgenic nicotiana tabacum. Plant Mol. Biol 1990, 15, 281–293. [Google Scholar]
- De Wilde, C.; de Rycke, R.; Beeckman, T.; de Neve, M.; Van Montagu, M.; Engler, G.; Depicker, A. Accumulation pattern of IgG antibodies and Fab fragments in transgenic Arabidopsis thaliana plants. Plant Cell Physiol 1998, 39, 639–646. [Google Scholar]
- Schouten, A.; Roosien, J.; van Engelen, F.A.; de Jong, G.A.; Borst-Vrenssen, A.W.; Zilverentant, J.F.; Bosch, D.; Stiekema, W.J.; Gommers, F.J.; Schots, A.; et al. The C-terminal KDEL sequence increases the expression level of a single-chain antibody designed to be targeted to both the cytosol and the secretory pathway in transgenic tobacco. Plant Mol. Biol. 1996, 30, 781–793. [Google Scholar]
- Bencurova, M.; Hemmer, W.; Focke-Tejkl, M.; Wilson, I.B.; Altmann, F. Specificity of IgG and IgE antibodies against plant and insect glycoprotein glycans determined with artificial glycoforms of human transferrin. Glycobiology 2004, 14, 457–466. [Google Scholar]
- Gomord, V.; Fitchette, A.C.; Menu-Bouaouiche, L.; Saint-Jore-Dupas, C.; Plasson, C.; Michaud, D.; Faye, L. Plant-specific glycosylation patterns in the context of therapeutic protein production. Plant Biotechnol. J 2010, 8, 564–587. [Google Scholar]
- Sarrion-Perdigones, A.; Falconi, E.E.; Zandalinas, S.I.; Juarez, P.; Fernandez-del-Carmen, A.; Granell, A.; Orzaez, D. Goldenbraid: An iterative cloning system for standardized assembly of reusable genetic modules. PLoS One 2011, 6, e21622. [Google Scholar]
- Weber, E.; Engler, C.; Gruetzner, R.; Werner, S.; Marillonnet, S. A modular cloning system for standardized assembly of multigene constructs. PLoS One 2011, 6, e16765. [Google Scholar]
- Monedero, V.; Rodriguez-Diaz, J.; Viana, R.; Buesa, J.; Perez-Martinez, G. Selection of single-chain antibodies against the VP8* subunit of rotavirus VP4 outer capsid protein and their expression in lactobacillus casei. Appl. Environ. Microbiol 2004, 70, 6936–6939. [Google Scholar]
- Juarez, P.; Presa, S.; Espi, J.; Pineda, B.; Anton, M.T.; Moreno, V.; Buesa, J.; Granell, A.; Orzaez, D. Neutralizing antibodies against rotavirus produced in transgenically labelled purple tomatoes. Plant Biotechnol. J 2012, 10, 341–352. [Google Scholar]
- Langley, R.; Wines, B.; Willoughby, N.; Basu, I.; Proft, T.; Fraser, J.D. The staphylococcal superantigen-like protein 7 binds IgA and complement C5 and inhibits IgA-Fc alpha Ri binding and serum killing of bacteria. J. Immunol 2005, 174, 2926–2933. [Google Scholar]
- Sack, M.; Paetz, A.; Kunert, R.; Bomble, M.; Hesse, F.; Stiegler, G.; Fischer, R.; Katinger, H.; Stoeger, E.; Rademacher, T. Functional analysis of the broadly neutralizing human anti-HIV-1 antibody 2F5 produced in transgenic BY-2 suspension cultures. FASEB J 2007, 21, 1655–1664. [Google Scholar]
- Ramessar, K.; Rademacher, T.; Sack, M.; Stadlmann, J.; Platis, D.; Stiegler, G.; Labrou, N.; Altmann, F.; Ma, J.; Stoger, E.; et al. Cost-effective production of a vaginal protein microbicide to prevent HIV transmission. Proc. Natl. Acad. Sci. USA 2008, 105, 3727–3732. [Google Scholar]
- Brandtzaeg, P.; Prydz, H. Direct evidence for an integrated function of J chain and secretory component in epithelial transport of immunoglobulins. Nature 1984, 311, 71–73. [Google Scholar]
- Johansen, F.E.; Braathen, R.; Brandtzaeg, P. Role of J chain in secretory immunoglobulin formation. Scand. J. Immunol 2000, 52, 240–248. [Google Scholar]
- Braathen, R.; Hohman, V.S.; Brandtzaeg, P.; Johansen, F.E. Secretory antibody formation: Conserved binding interactions between J chain and polymeric Ig receptor from humans and amphibians. J. Immunol 2007, 178, 1589–1597. [Google Scholar]
- Murthy, A.K.; Chaganty, B.K.; Troutman, T.; Guentzel, M.N.; Yu, J.J.; Ali, S.K.; Lauriano, C.M.; Chambers, J.P.; Klose, K.E.; Arulanandam, B.P. Mannose-containing oligosaccharides of non-specific human secretory immunoglobulin a mediate inhibition of Vibrio cholerae biofilm formation. PLoS One 2011, 6, e16847. [Google Scholar]
- Mathias, A.; Corthesy, B. N-Glycans on secretory component: Mediators of the interaction between secretory IgA and gram-positive commensals sustaining intestinal homeostasis. Gut Microbes 2011, 2, 287–293. [Google Scholar]
- Hu, L.; Crawford, S.E.; Czako, R.; Cortes-Penfield, N.W.; Smith, D.F.; le Pendu, J.; Estes, M.K.; Prasad, B.V. Cell attachment protein VP8* of a human rotavirus specifically interacts with a-type histo-blood group antigen. Nature 2012, 485, 256–259. [Google Scholar]
- Yu, X.; Dang, V.T.; Fleming, F.E.; von Itzstein, M.; Coulson, B.S.; Blanchard, H. Structural basis of rotavirus strain preference towards N-acetyl- or N-glycolylneuraminic acid-containing receptors. J. Virol 2012, 86, 13456–13466. [Google Scholar]
- Haselhorst, T.; Fleming, F.E.; Dyason, J.C.; Hartnell, R.D.; Yu, X.; Holloway, G.; Santegoets, K.; Kiefel, M.J.; Blanchard, H.; Coulson, B.S.; et al. Sialic acid dependence in rotavirus host cell invasion. Nat. Chem. Biol. 2009, 5, 91–93. [Google Scholar]
- Bonner, A.; Almogren, A.; Furtado, P.B.; Kerr, M.A.; Perkins, S.J. The nonplanar secretory IgA2 and near planar secretory IgA1 solution structures rationalize their different mucosal immune responses. J. Biol. Chem 2009, 284, 5077–5087. [Google Scholar]
- Yoo, E.M.; Morrison, S.L. IgA: An immune glycoprotein. Clin. Immunol 2005, 116, 3–10. [Google Scholar]
- Karnoup, A.S.; Turkelson, V.; Anderson, W.H.K. O-linked glycosylation in maize-expressed human IgA1. Glycobiology 2005, 15, 965–981. [Google Scholar]
- Ramsland, P.A.; Willoughby, N.; Trist, H.M.; Farrugia, W.; Hogarth, P.M.; Fraser, J.D.; Wines, B.D. Structural basis for evasion of IgA immunity by Staphylococcus aureus revealed in the complex of SSL7 with Fc of human IgA1. Proc. Natl. Acad. Sci 2007, 104, 15051–15056. [Google Scholar]
- Almogren, A.; Senior, B.W.; Kerr, M.A. A comparison of the binding of secretory component to immunoglobulin A (IgA) in human colostral S-IgA1 and S-IgA2. Immunology 2007, 120, 273–280. [Google Scholar]
- Orzaez, D.; Mirabel, S.; Wieland, W.H.; Granell, A. Agroinjection of tomato fruits. A tool for rapid functional analysis of transgenes directly in fruit. Plant. Physiol 2006, 140, 3–11. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| LCλ/SC | LCλ/SCkdel | LCκ/SC | LCκ/SCkdel | |
|---|---|---|---|---|
| HCα1 | ++ | ++ | − | − |
| HCα1kdel | +++ | ++ | − | − |
| HCα2 | ++ | ++ | − | − |
| HCα2kdel | ++ | ++ | − | − |
| Construct | Best Performing Combination | Total IgA (%TSP) | Total IgA (μg/g FW) | sIgA (%TSP) | sIgA (μg/g FW) |
|---|---|---|---|---|---|
| IgA | HCα1kdel-LCλ | 1.1 ± 0.1 | 31.6 ± 3.7 | 0 | 0 |
| sIgA | HCα1-LCλ-JC-SCkdel | 1.5 ± 0.1 | 32.5 ± 1.1 | 0.5 ± 0.1 | 11.0 ± 0.2 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Juarez, P.; Huet-Trujillo, E.; Sarrion-Perdigones, A.; Falconi, E.E.; Granell, A.; Orzaez, D. Combinatorial Analysis of Secretory Immunoglobulin A (sIgA) Expression in Plants. Int. J. Mol. Sci. 2013, 14, 6205-6222. https://doi.org/10.3390/ijms14036205
Juarez P, Huet-Trujillo E, Sarrion-Perdigones A, Falconi EE, Granell A, Orzaez D. Combinatorial Analysis of Secretory Immunoglobulin A (sIgA) Expression in Plants. International Journal of Molecular Sciences. 2013; 14(3):6205-6222. https://doi.org/10.3390/ijms14036205
Chicago/Turabian StyleJuarez, Paloma, Estefania Huet-Trujillo, Alejandro Sarrion-Perdigones, Erica Elvira Falconi, Antonio Granell, and Diego Orzaez. 2013. "Combinatorial Analysis of Secretory Immunoglobulin A (sIgA) Expression in Plants" International Journal of Molecular Sciences 14, no. 3: 6205-6222. https://doi.org/10.3390/ijms14036205
APA StyleJuarez, P., Huet-Trujillo, E., Sarrion-Perdigones, A., Falconi, E. E., Granell, A., & Orzaez, D. (2013). Combinatorial Analysis of Secretory Immunoglobulin A (sIgA) Expression in Plants. International Journal of Molecular Sciences, 14(3), 6205-6222. https://doi.org/10.3390/ijms14036205