DNA Self-Assembly: From Chirality to Evolution

Abstract
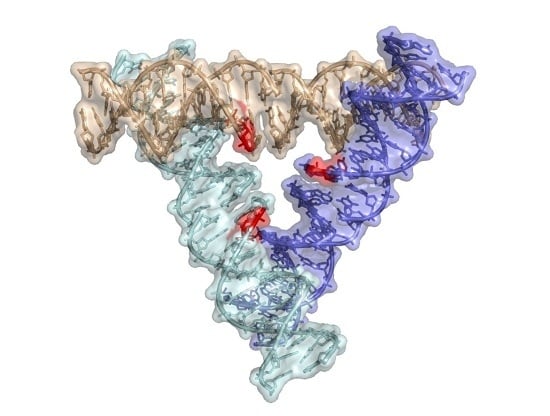
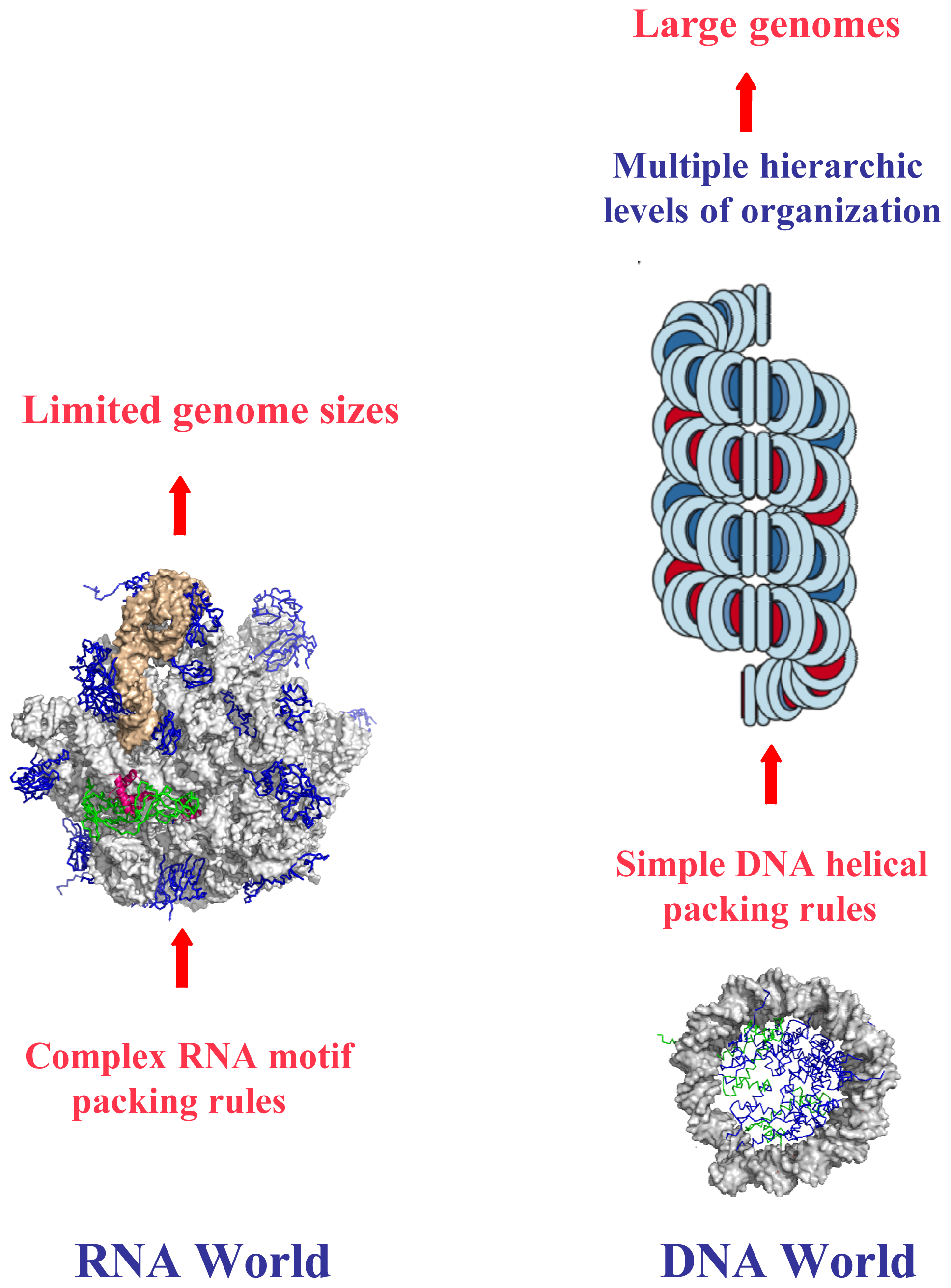
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. From Double-Helix Chirality to Stable DNA Self-Fitting
2.1. Ubiquitous Crossovers
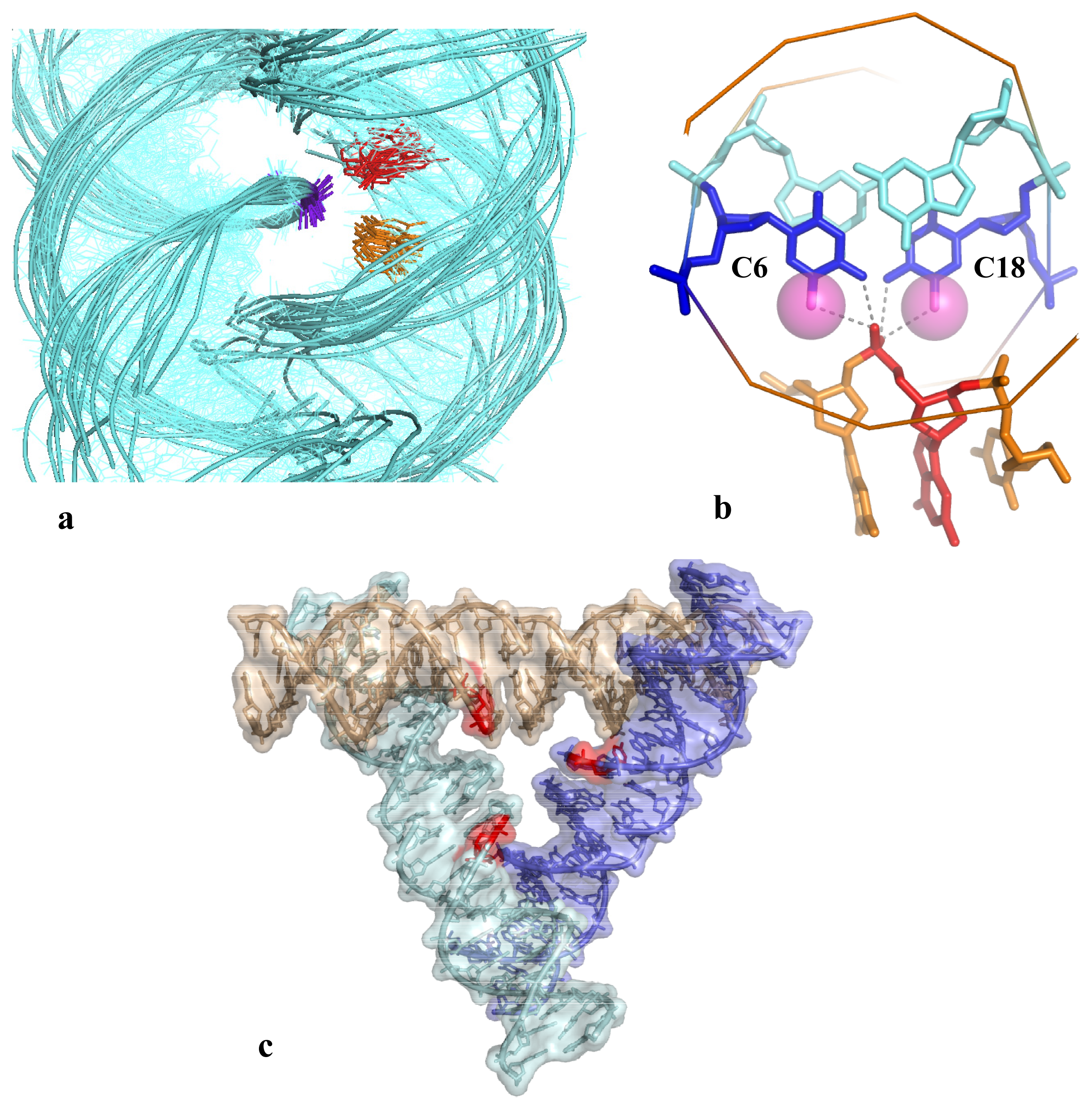
2.2. Cytosine and DNA Self-Assembly
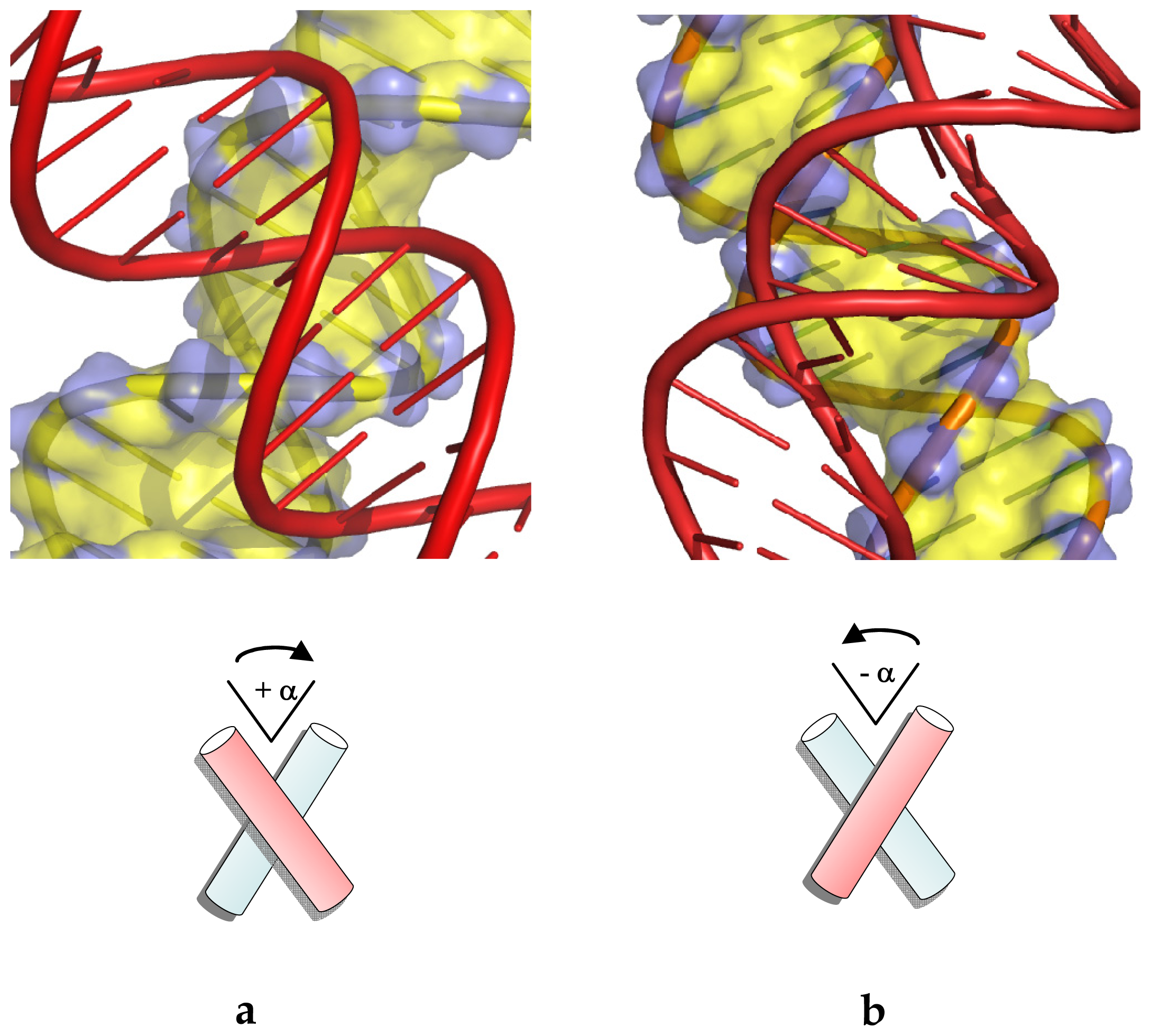
2.3. Differential Stability of Chiral Crossovers
2.4. Role of Divalent Cations in DNA Assembly
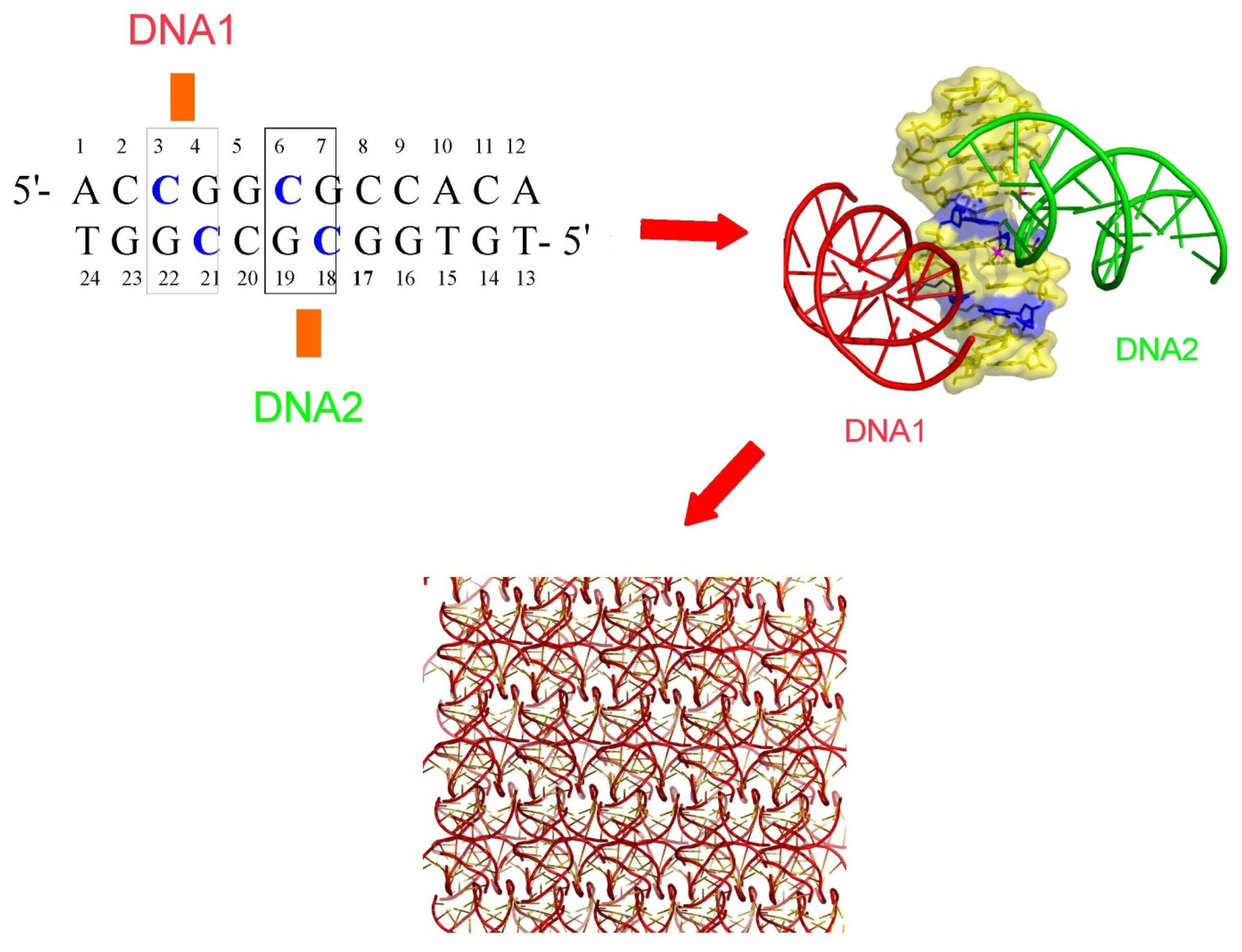
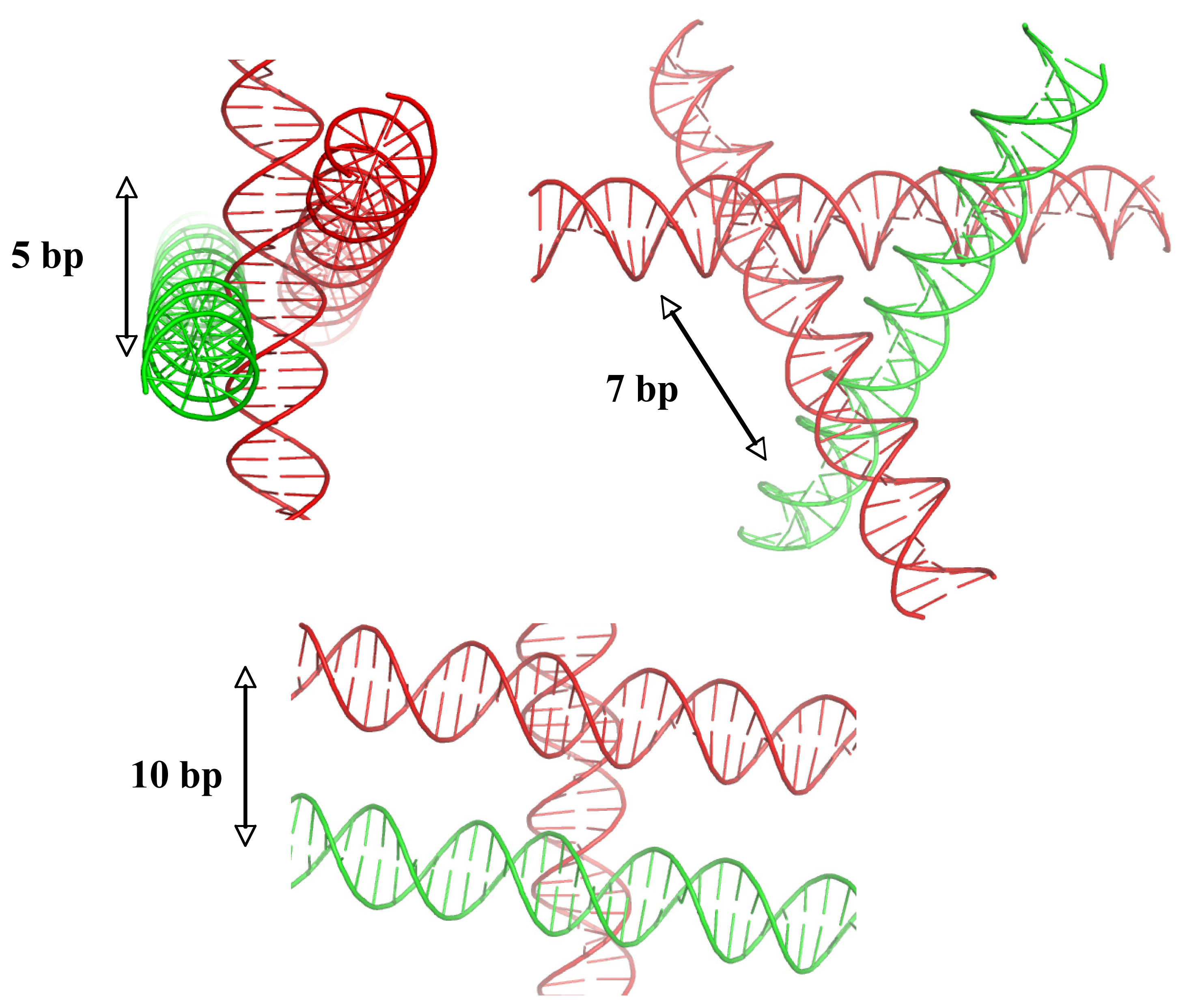
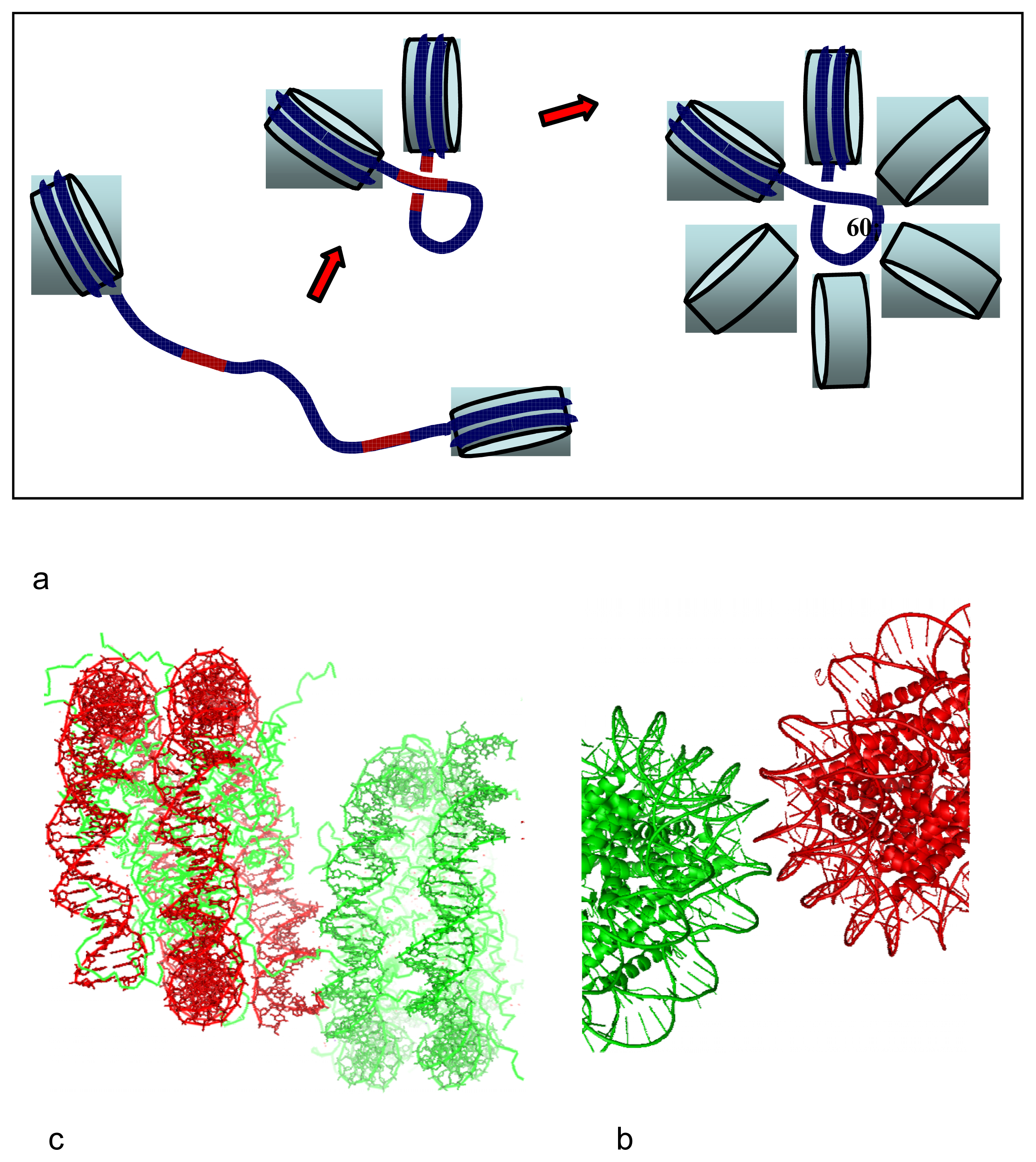
2.5. DNA Chirality and Sequence Controls the Architecture of Higher-Order DNA Structures
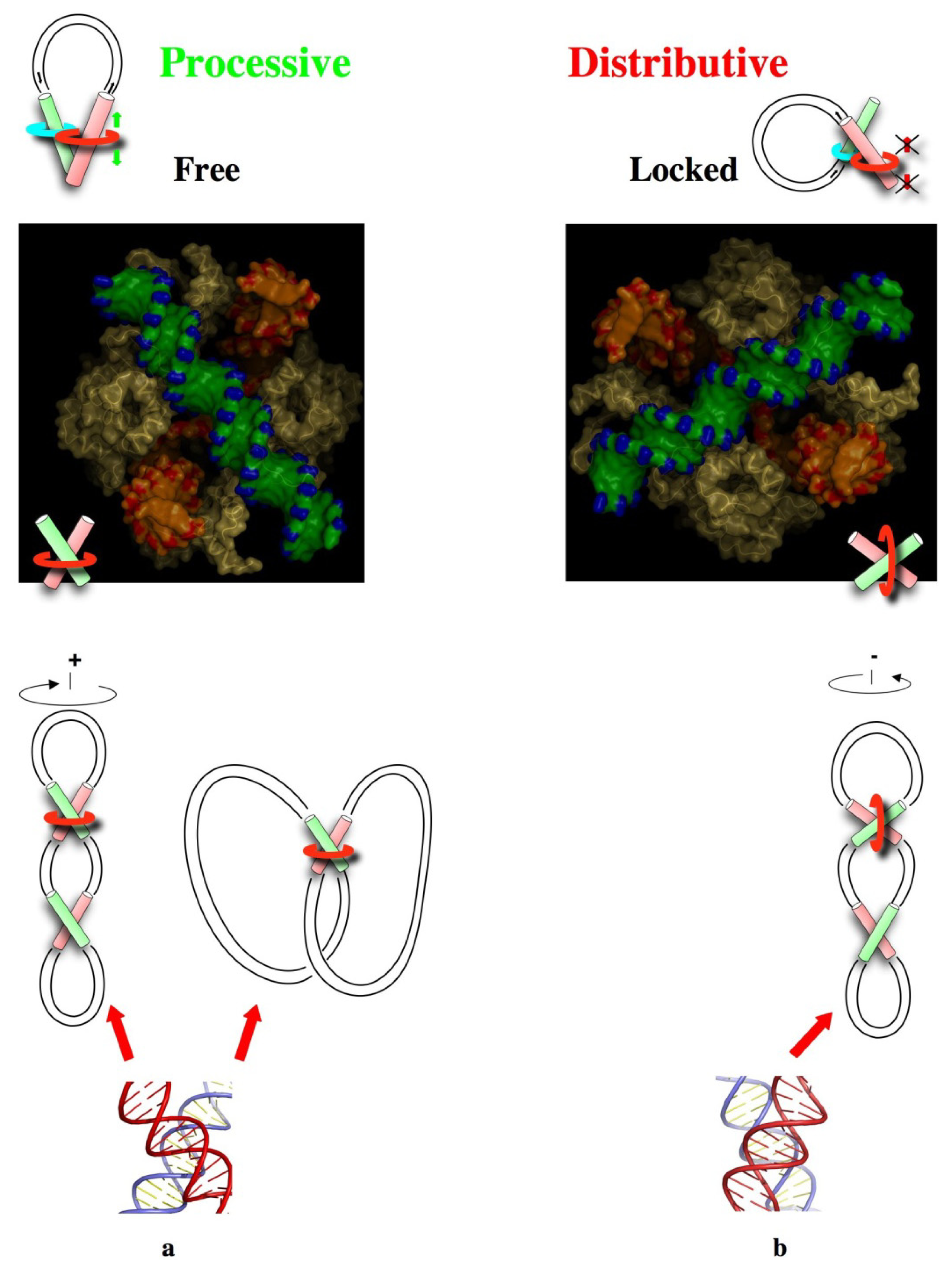
3. From Local DNA-DNA Interactions to Global DNA Topology
4. DNA Self-Assembly and Evolution
5. Conclusions and Future Prospects
References
- Echols, H. Nucleoprotein structures initiating DNA-replication, transcription, and site-specific recombination. J. Biol. Chem 1990, 265, 14697–14700. [Google Scholar]
- Grosschedl, R. Higher-order nucleoprotein complexes in transcription—Analogies with site-specific recombination. Curr. Opin. Cell Biol 1995, 7, 362–370. [Google Scholar]
- Segal, E.; Widom, J. What controls nucleosome positions? Trends Genet 2009, 25, 335–343. [Google Scholar]
- Bloomfield, V.A. DNA condensation. Curr. Opin. Struct. Biol 1996, 6, 334–341. [Google Scholar]
- Strey, H.H.; Podgornik, R.; Rau, D.C.; Parsegian, V.A. DNA-DNA interactions. Curr. Opin. Struct. Biol 1998, 8, 309–313. [Google Scholar]
- Schellman, J.A.; Parthasarathy, N. X-Ray-diffraction studies on cation-collapsed DNA. J. Mol. Biol 1984, 175, 313–329. [Google Scholar]
- Raspaud, E.; Durand, D.; Livolant, F. Interhelical spacing in liquid crystalline spermine and spermidine-DNA precipitates. Biophys. J 2005, 88, 392–403. [Google Scholar]
- Kornyshev, A.A.; Leikin, S. Electrostatic interaction between helical macromolecules in dense aggregates: An impetus for DNA poly- and meso-morphism. Proc. Natl. Acad. Sci. USA 1998, 95, 13579–13584. [Google Scholar]
- Minsky, A. Information content and complexity in the high-order organization of DNA. Annu. Rev. Biophys. Biomol. Struct 2004, 33, 317–342. [Google Scholar]
- Zanchetta, G.; Giavazzi, F.; Nakata, M.; Buscaglia, M.; Cerbino, R.; Clark, N.; Bellini, T. Right-handed double helix ultrashort DNA yields chiral nematic phases with both right and left-handed director twist. Proc. Natl. Acad. Sci. USA 2010, 107, 17797, –17502.. [Google Scholar]
- Srinivasan, A.R.; Olson, W.K. DNA associations: packing calculation in A, B- and Z-DNA structures. Biophys. Chem 1992, 43, 279–310. [Google Scholar]
- Timsit, Y.; Moras, D. Crystallization of DNA. Meth. Enzymol 1992, 211, 409–429. [Google Scholar]
- Murthy, V.L.; Rose, G.D. Is counterion delocalization responsible for collapse in RNA folding? Biochemistry 2000, 39, 14365–14370. [Google Scholar]
- Qiu, X.Y.; Kwok, L.W.; Park, H.Y.; Lamb, J.S.; Andresen, K.; Pollack, L. Measuring inter-DNA potentials in solution. Phys. Rev. Lett 2006, 96, 138101–138104. [Google Scholar]
- Tan, Z.J.; Chen, S.J. Electrostatic free energy landscapes for nucleic acid helix assembly. Nucleic Acids Res 2006, 34, 6629–6639. [Google Scholar]
- Inoue, S.; Sugiyama, S.; Travers, A.A.; Ohyama, T. Self-assembly of double-stranded DNA molecules at nanomolar concentrations. Biochemistry 2007, 46, 164–171. [Google Scholar]
- Qiu, X.; Andresen, K.; Kwok, L.W.; Lamb, J.S.; Park, H.Y.; Pollack, L. Inter-DNA attraction mediated by divalent counterions. Phys. Rev. Lett 2007, 99, 038104–038107. [Google Scholar]
- Varnai, P.; Timsit, Y. Differential stability of chiral DNA crossovers mediated by divalent cations. Nucl. Acid Res 2010, 38, 4163–4172. [Google Scholar]
- Timsit, Y.; Varnai, P. Helical chirality: A link between local interactions and global topology in DNA. PLoS One 2010, 5, e9326. [Google Scholar]
- Timsit, Y.; Varnai, P. Cytosine, the double helix and DNA self-assembly. J. Mol. Recognit 2011, 24, 137–138. [Google Scholar]
- Timsit, Y. Local sensing of global DNA topology: From crossover geometry to type II topoisomerase processivity. Nuceic Acids Res 2011, 39, 8665–8676. [Google Scholar]
- Timsit, Y.; Vilbois, E.; Moras, D. Base pairing shift in the major groove of CAn tracts by B-DNA crystal structures. Nature 1991, 354, 167–170. [Google Scholar]
- Timsit, Y.; Moras, D. Groove-Backbone Interaction in B-DNA—Implication for DNA condensation and recombination. J. Mol. Biol 1991, 221, 919–940. [Google Scholar]
- Timsit, Y.; Moras, D. DNA self-fitting: The double helix directs the geometry of its supramolecular assemblies. EMBO J 1994, 13, 2737–2746. [Google Scholar]
- Lu, X.-J.; Shakked, Z.; Olson, W. A-form conformational motifs in ligand-bound DNA structures. J. Mol. Biol 2000, 300, 819–840. [Google Scholar]
- Nissen, P.; Ippolito, J.A.; Ban, N.; Moore, P.; Steitz, T. RNA tertiary interactions in the large ribosomal subunit: The A-minor motif. Proc. Natl. Acad. Sci. USA 2001, 98, 4899–4903. [Google Scholar]
- Schindelin, H.; Zhang, M.; Bald, R.; Fürste, J.-P.; Erdmann, V.A.; Heinemann, U. Crystal structure of an RNA dodecamer containing the Escherichia coli Shine-Dalgarno sequence. J. Mol. Biol 1995, 249, 595–603. [Google Scholar]
- Baeyens, K.; de Bondt, H.L.; Holbrook, S.R. Structure of an RNA double helix including uracil-uracil base pairs in an internal loop. Nat. Struct. Biol 1996, 2, 56–62. [Google Scholar]
- Gagnon, M.; Steinberg, S. GU receptors of double helices mediate tRNA movement in the ribosome. RNA 2002, 8, 873–877. [Google Scholar]
- Gagnon, M.G.; Mukhopadhyay, A.; Steinberg, S.V. Close packing of helices 3 and 12 of 16S rRNA is required for the normal ribosome function. J. Biol. Chem 2006, 281, 39349–39357. [Google Scholar]
- Timsit, Y.; Shatzky-Schwartz, M.; Shakked, Z. Left-handed DNA crossovers. Implications for DNA-DNA recognition and structural alterations. J. Biomol. Struct. Dyn 1999, 16, 775–785. [Google Scholar]
- Srinivasan, A.R.; Sauers, R.R.; Fenley, M.O.; Boschitsch, A.H.; Matsumoto, A.; Colasanti, A.V.; Olson, W.K. Properties of the nucleic-acid bases in free and Watson-Crick hydrogen-bonded states: Computational insights into the sequence-dependent features of double-helical DNA. Biophys. Rev 2009, 1, 13–20. [Google Scholar]
- Auffinger, P.; Bielecki, L.; Westhof, E. Anion binding to nucleic acids. Structure 2004, 12, 379–388. [Google Scholar]
- Mayer-Jung, C.; Moras, D.; Timsit, Y. Effect of cytosine methylation on DNA-DNA recognition at CpG steps. J. Mol. Biol 1997, 270, 328–335. [Google Scholar]
- Zheng, J.; Birktoft, J.J.; Chen, Y.; Wang, T.; Sha, R.; Constantinou, P.E.; Ginell, S.L.; Mao, C.; Seeman, N.C. From molecular to macroscopic via the rational design of self-assembled 3D DNA crystal. Nature 2009, 461, 74–77. [Google Scholar]
- Ortiz-Lombardia, M.; Gonzalez, A.; Eritja, R.; Aymami, J.; Azorin, F.; Coll, M. Crystal structure of a DNA Holliday junction. Nat. Struct. Biol 1999, 6, 913–917. [Google Scholar]
- Khuu, P.; Regier Voth, A.; Hays, F.; Ho, P.S. The stacked-X DNA Holliday junction and protein recognition. J. Mol. Recogn 2006, 19, 234–242. [Google Scholar]
- Shlyakhtenko, L.S.; Miloseska, L.; Potaman, V.N.; Sinden, R.R.; Lyubchenko, Y.L. Intersegmental interactions in supercoiled DNA: Atomic force microscope study. Ultramicroscopy 2003, 97, 263–270. [Google Scholar]
- Lilley, D.M.J. Structures of helical junctions in nucleic acids. Q. Rev. Biophys 2000, 33, 109–159. [Google Scholar]
- Van Buuren, B.N.M.; Hermann, T.; Wijmenga, S.S.; Westhof, E. Brownian-dynamics simulations of metal-ion binding to four-way junctions. Nucleic Acids Res 2002, 30, 507–514. [Google Scholar]
- Tinoco, I., Jr; Kieft, JS. The ion core in RNA folding. Nat. Struct. Biol 1997, 4, 509–512. [Google Scholar]
- Klein, D.J.; Moore, P.B.; Steitz, T.A. The contribution of metal ions to the structural stability of the large ribosomal subunit. RNA 2004, 10, 1366–1379. [Google Scholar]
- Woodson, S.A. Metal ions and RNA folding: A highly charged topic with a dynamic future. Curr. Opin. Chem. Biol 2005, 9, 104–109. [Google Scholar]
- Robinson, P.J.J.; Rhodes, D. Structure of the ‘30 nm’ chromatin fibre: A key role for the linker histone. Curr. Opin. Struct. Biol 2006, 16, 336–343. [Google Scholar]
- Wu, C.Y.; Bassett, A.; Travers, A. A variable topology for the 30-nm chromatin fibre. EMBO Rep 2007, 8, 1129–1134. [Google Scholar]
- Robinson, P.J.J.; Fairall, L.; Huynh, V.A.T.; Rhodes, D. EM measurements define the dimensions of the “30-nm” chromatin fiber: Evidence for a compact, interdigitated structure. Proc. Natl. Acad. Sci. USA 2006, 103, 6506–6511. [Google Scholar]
- Schalch, T.; Duda, S.; Sargent, D.F.; Richmond, T.J. X-ray structure of a tetranucleosome and its implications for the chromatin fibre. Nature 2005, 436, 138–141. [Google Scholar]
- Davey, C.A.; Richmond, T.J. DNA-dependent divalent cation binding in the nucleosome core particle. Proc. Natl. Acad. Sci. USA 2002, 99, 11169–11174. [Google Scholar]
- Wong, H.; Victor, J.M.; Mozziconacci, J. An all-atom model of the chromatin fiber containing linker histones reveals a versatile structure tuned by the nucleosomal repeat length. PLoS One 2007, 2, e877. [Google Scholar]
- Vologodskii, A.V.; Cozzarelli, N.R. Conformational and thermodynamic properties of supercoiled DNA. Annu. Rev. Biophys. Biomol. Struc 1994, 23, 609–643. [Google Scholar]
- Schlick, T.; Olson, W.K. Trefoil knotting revealed by molecular-dynamics simulations of supercoiled DNA. Science 1992, 257, 1110–1115. [Google Scholar]
- Wang, J.C. Cellular roles of DNA topoisomerases: A molecular perspective. Nat. Rev 2002, 3, 430–440. [Google Scholar]
- Travers, A.; Muskhelishvili, G. A common topology for bacterial and eukaryotic transcription initiation? EMBO Rep 2007, 8, 147–151. [Google Scholar]
- Stupina, V.A.; Wang, J.C. DNA axial rotation and the merge of oppositely supercoiled DNA domains in Escherichia coli: Effects of DNA bends. Proc. Natl. Acad. Sci. USA 2004, 101, 8608–8613. [Google Scholar]
- Postow, L.; Crisona, N.J.; Peter, B.J.; Hardy, C.D.; Cozzarelli, N.R. Topological challenges to DNA replication: Conformations at the fork. Proc. Natl. Acad. Sci. USA 2001, 98, 8219–8226. [Google Scholar]
- Liu, L.F.; Wang, J.C. Supercoiling of the DNA template during transcription. Proc. Natl. Acad. Sci. USA 1987, 84, 7024–7027. [Google Scholar]
- Roca, J.; Wang, J.C. The probabilities of supercoil removal and decatenation by yeast DNA topoisomerase II. Genes Cells 1996, 1, 17–27. [Google Scholar]
- Rybenkov, V.V.; Ullsperger, C.; Vologodskii, A.V.; Cozzarelli, N.R. Simplification of DNA topology below equilibrium values by type II topoisomerases. Science 1997, 277, 690–693. [Google Scholar]
- Shaw, S.; Wang, J.C. Chirality of DNA trefoils: Implications in intramolecular synapsis of distant DNA segments. Proc. Natl. Acad. Sci. USA 1997, 94, 1692–1697. [Google Scholar]
- Crisona, N.J.; Strick, T.R.; Bensimon, D.; Croquette, V.; Cozzarelli, N.R. Preferential relaxation of positively supercoiled DNA by E. coli topoisomerase IV in single-molecule and ensemble measurements. Gene Dev 2000, 14, 2881–2892. [Google Scholar]
- Charvin, G.; Bensimon, D.; Croquette, V. Single-molecule study of DNA unlinking by eukaryotic and prokaryotic type-II topoisomerases. Proc. Natl. Acad. Sci. USA 2003, 100, 9820–9825. [Google Scholar]
- Stone, M.D.; Bryant, Z.; Crisona, N.J.; Smith, S.B.; Vologodskii, A.; Bustamente, C.; Cozzarelli, N.R. Chirality sensing by Escherichia coli topoisomerase IV and the mechanism of type II topoisomerases. Proc. Natl. Acad. Sci. USA 2003, 100, 8654–8659. [Google Scholar]
- Nöllmann, N.; Stone, M.D.; Bryant, Z.; Gore, J.; Crisona, N.J.; Hong, S.C.; Mitelheiser, S.; Maxwell, A.; Bustamante, C.; Cozzarelli, N.R. Multiple modes of Escherichia coli DNA gyrase activity revealed by force and torque. Nat. Struct. Mol. Biol 2007, 14, 264–271. [Google Scholar]
- McClendon, A.K.; Rodriguez, A.C.; Osheroff, N. Human topoisomerase IIα rapidly relaxes positively supercoiled DNA. Implications for enzyme action ahead of replication forks. J. Biol. Chem 2005, 280, 39337–39345. [Google Scholar]
- Pulleyblank, D.E. Of Topo and Maxwell’s dream. Science 1997, 277, 648–649. [Google Scholar]
- Dong, K.C.; Berger, J.M. Structural basis for gate-DNA recognition and bending by type IIA topoisomerases. Nature 2007, 450, 1201–1206. [Google Scholar]
- Yan, J.; Magnasco, M.O.; Marko, J.F. A kinetic proofreading mechanism for disentanglement of DNA by topoisomerases. Nature 1999, 401, 932–935. [Google Scholar]
- Buck, G.R.; Zechiedrich, E.L. DNA disentangling by type-2 topoisomerases. J. Mol. Biol 2004, 340, 933–939. [Google Scholar]
- Randall, G.L.; Pettitt, B.M.; Buck, G.R.; Zechiedriech, E.L. Electrostatics of DNA–DNA juxtapositions: Consequences for type II topoisomerase function. J. Phys. Condens. Matter 2006, 18, S173–S185. [Google Scholar]
- Corbett, K.D.; Schoeffler, A.J.; Thomsen, N.D.; Berger, J.B. The structural basis for substrate specificity in DNA topoisomerase IV. J. Mol. Biol 2005, 351, 545–561. [Google Scholar]
- Neuman, K.C.; Charvin, G.; Bensimon, D.; Croquette, V. Mechanisms of chiral discrimination by topoisomerase IV. Proc. Natl. Acad. Sci. USA 2009, 106, 6986–6991. [Google Scholar]
- Zechiedrich, E.L.; Osheroff, N. Eukaryotic topoisomerases recognize nucleic acid topology by preferentially interacting with DNA crossovers. EMBO J 1990, 9, 4555–4562. [Google Scholar]
- Forterre, P.; Gadelle, D. Phylogenomics of DNA topoisomerases: Their origin and putative roles in the emergence of modern organisms. Nucleic Acids Res 2009, 37, 679–692. [Google Scholar]
- Panyutin, I.G.; Hsieh, P. The kinetics of spontaneous DNA branch migration. Proc. Natl. Acad. Sci. USA 1994, 91, 2021–2025. [Google Scholar]
- Olson, W.K.; Zhurkin, V.B. Modeling DNA deformations. Curr. Opin. Struc. Biol 2000, 10, 286–297. [Google Scholar]
- Travers, A.A. The structural basis of DNA flexibility. Philos. Trans. Roy. Soc. A 2004, 362, 1423–1438. [Google Scholar]
- Brochier-Armanet, C.; Forterre, P. Widespread archael reverse gyrase in thermophilic bacteria suggest a complex history of vertical inheritance and lateral gene transfers. Archaea 2006, 2, 83–93. [Google Scholar]
- Charbonnier, F.; Forterre, P. Comparison of plasmid DNA topology among mesophilic and thermophilic eubacteria and archaebacteria. J. Bacteriol 1994, 176, 1251–1259. [Google Scholar]
- Guipaud, O.; Marguet, E.; Noll, K.; Bouthier de la Tour, C.; Forterre, P. Both gyrase and reverse gyrase are present in the hyperthermophilic bacterium Thermogata maritima. Proc. Natl. Acad. Sci. USA 1997, 94, 10606–10611. [Google Scholar]
- Lopez-Garcia, P.; Forterre, P.; van der Oost, J.; Erauso, G. Plasmid pGS5 from hyperthermophilic archaeon archaeoglobus profundus is negatively supercoiled. J. Bacteriol 2000, 182, 4998–5000. [Google Scholar]
- Marguet, E.; Forterre, P. DNA stability at temperatures typical for hyperthermophiles. Nucleic Acids Res 1994, 22, 1681–1686. [Google Scholar]
- Leontis, N.B.; Westhof, E. Geometric nomenclature and classification of RNA base pairs. RNA 2001, 7, 499–512. [Google Scholar]
- Leontis, N.B.; Lescoute, A.; Westhof, E. The buidling blocks and motifs of RNA architecture. Curr. Opin. Struct. Biol 2006, 16, 279–287. [Google Scholar]
- Batey, R.T.; Rambo, R.P.; Doudna, J.A. Tertiary motifs in RNA structure and folding. Angew. Chem. Int. Ed 1999, 38, 2326–2343. [Google Scholar]
- Timsit, Y. DNA structure and polymerase fidelity. J. Mol. Biol 1999, 293, 835–853. [Google Scholar]
- Timsit, Y. DNA-directed base pair opening. Molecules 2012, 17, 11047–11064. [Google Scholar]







© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Timsit, Y. DNA Self-Assembly: From Chirality to Evolution. Int. J. Mol. Sci. 2013, 14, 8252-8270. https://doi.org/10.3390/ijms14048252
Timsit Y. DNA Self-Assembly: From Chirality to Evolution. International Journal of Molecular Sciences. 2013; 14(4):8252-8270. https://doi.org/10.3390/ijms14048252
Chicago/Turabian StyleTimsit, Youri. 2013. "DNA Self-Assembly: From Chirality to Evolution" International Journal of Molecular Sciences 14, no. 4: 8252-8270. https://doi.org/10.3390/ijms14048252
APA StyleTimsit, Y. (2013). DNA Self-Assembly: From Chirality to Evolution. International Journal of Molecular Sciences, 14(4), 8252-8270. https://doi.org/10.3390/ijms14048252