Oxidative and Molecular Responses in Capsicum annuum L. after Hydrogen Peroxide, Salicylic Acid and Chitosan Foliar Applications



Abstract
:1. Introduction
2. Results and Discussion
2.1. H2O2 Detection with DAB in C. annuum L. due to Elicitors’ Application
2.2. Quantitative Analysis of the H2O2 Endogenous Content in Capsicum annuum L
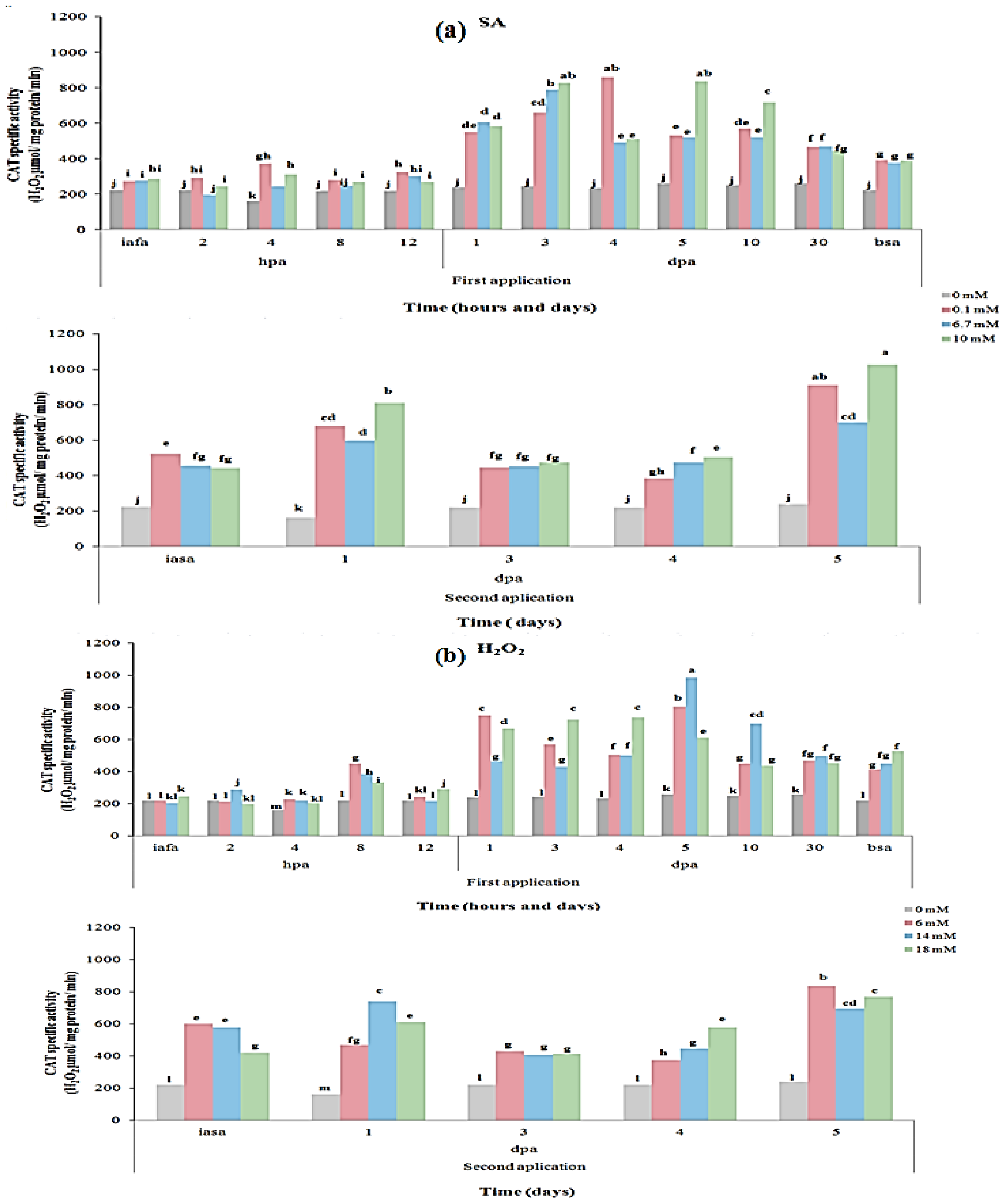
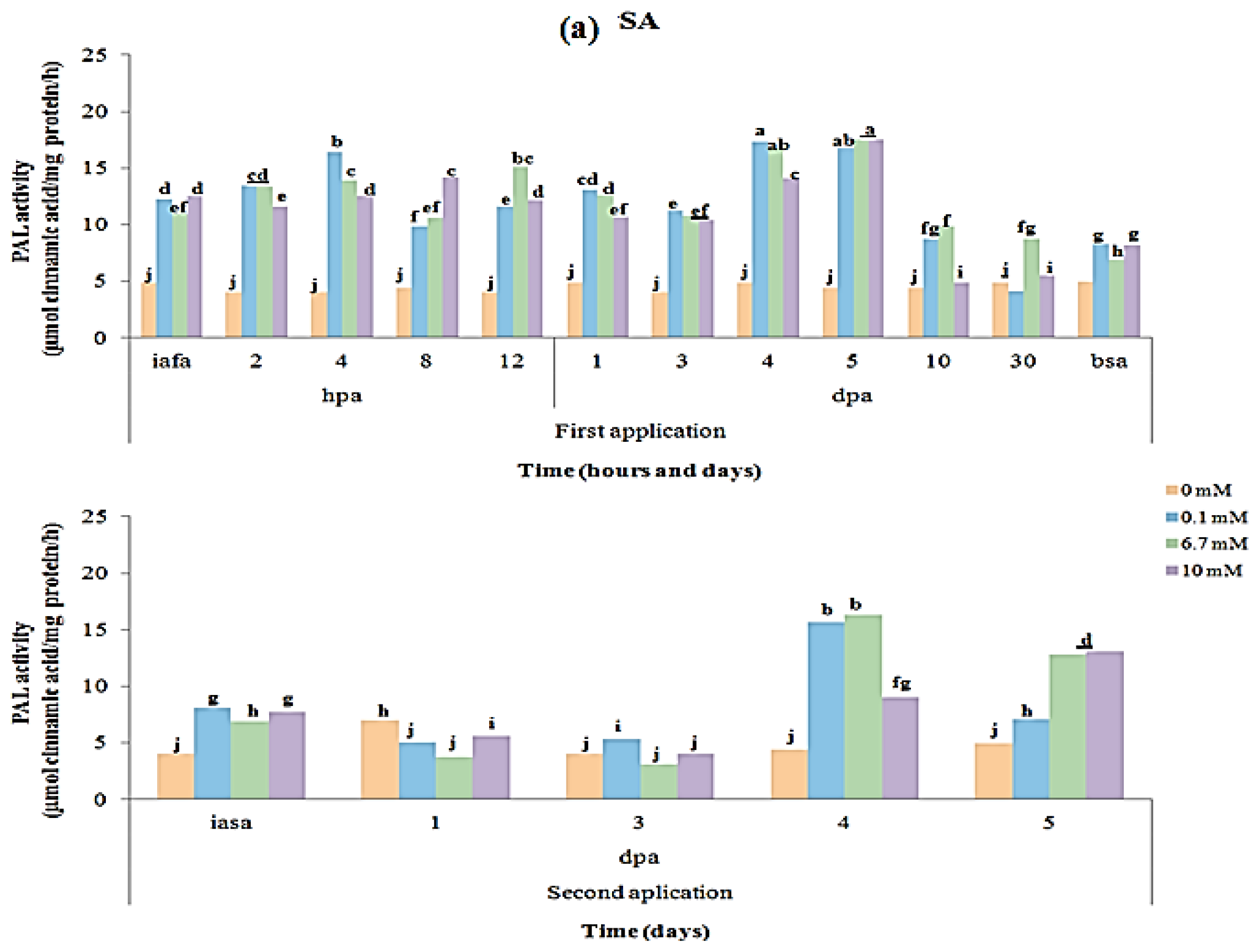
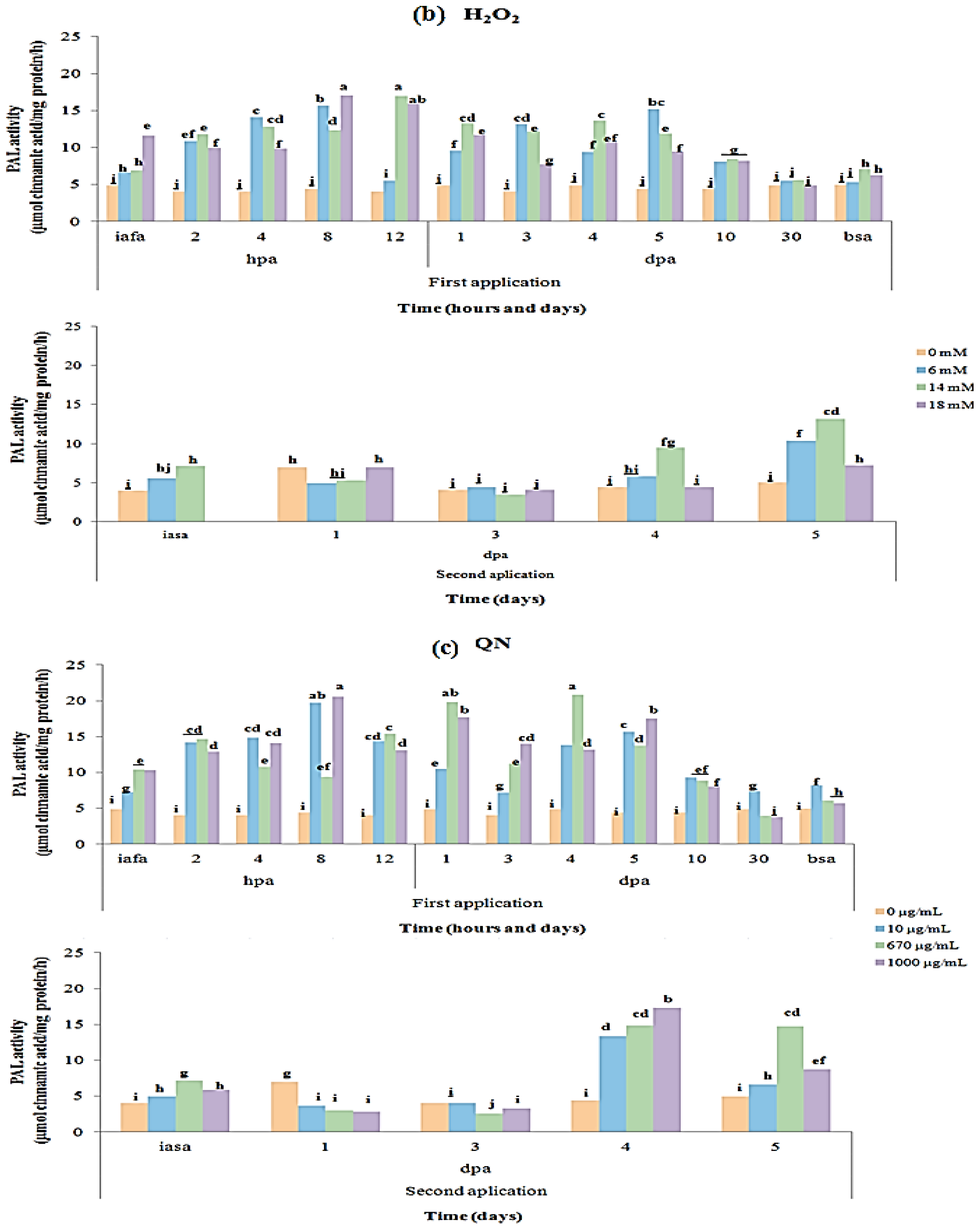
2.3. CAT and PAL Enzyme Activity
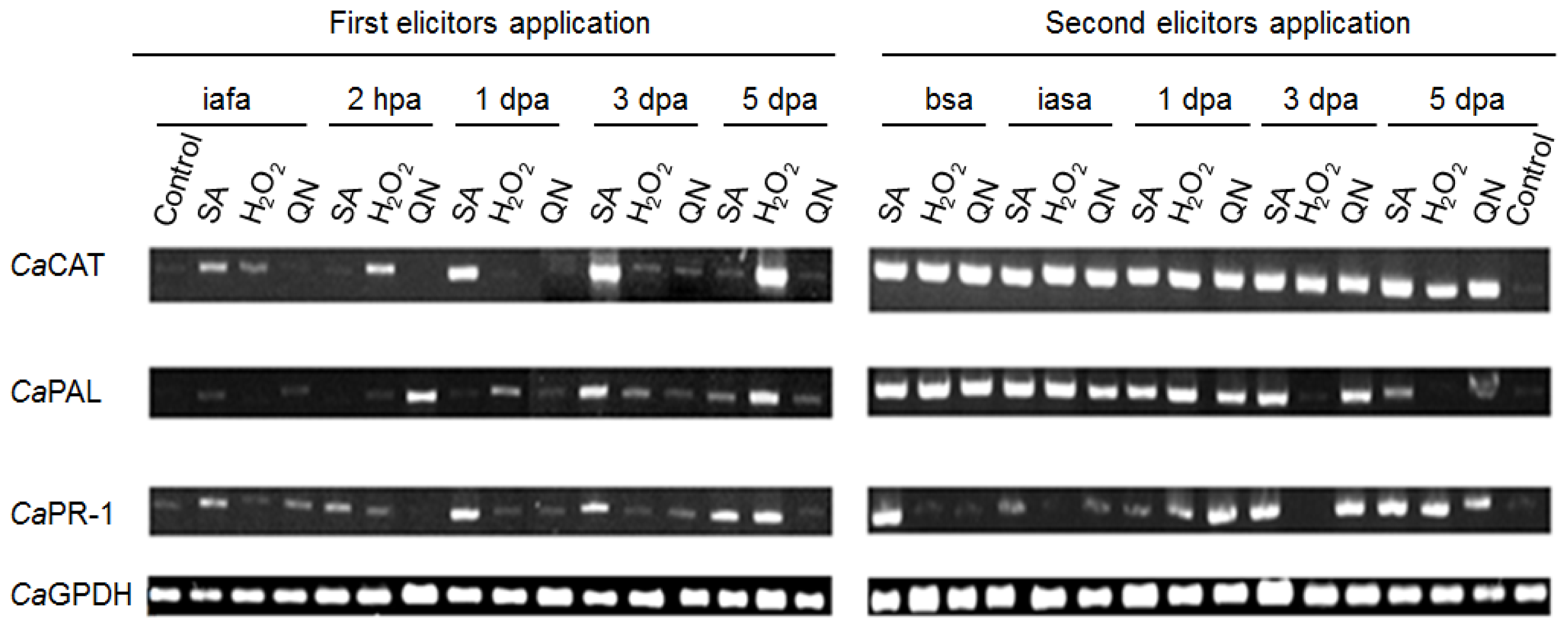
2.4. Gene Expression Pattern of cat1, pal, and pr1 in C. annuum L. var. Don Benito
3. Experimental Section
3.1. Plant Growth
3.2. Stress Treatments with Elicitors’ Application
3.3. H2O2 Detection with DAB Staining
3.4. Plant Extracts Preparation
3.5. H2O2 Content Assay
3.6. Enzyme Activity Assays
3.7. Gene Expression Analysis of cat1, pal, and pr1 in Capsicum annuum L
3.8. Statistical Analyses
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signalling molecules in plants. J. Exp. Bot 2002, 53, 1237–1247. [Google Scholar]
- Gechev, T.S.; Hille, J. Hydrogen peroxide as a signal controlling plant programmed cell death. J. Cell Biol 2005, 168, 17–20. [Google Scholar]
- Pitzschke, A.; Hirt, H. Mitogen-Activated Protein Kinases and Reactive Oxygen Species Signaling in Plants. Plant Physiol 2006, 141, 351–356. [Google Scholar]
- Fones, H.; Preston, G.M. Reactive oxygen and oxidative stress tolerance in plant pathogenic Pseudomonas. FEMS Microbiol. Lett 2011, 327, 1–8. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol 2004, 55, 373–399. [Google Scholar]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; van Breusegem, F. Reactive oxygen gene network of plants. Trends. Plant Sci 2004, 9, 490–498. [Google Scholar]
- Inzé, A.; Vanderauwera, S.; Hoeberichts, F.A.; Vandorpe, M.; Van Gaever, T.; van Breusegem, F. A subcellular localization compendium of hydrogen peroxide-induced proteins. Plant Cell. Environ 2012, 35, 308–320. [Google Scholar]
- Holopainen, J.K.; Heijari, J.; Nerg, A.M.; Vuorinen, M.; Kainulainen, P. Potential for the use of exogenous chemical elicitors in disease and insect pest management of conifer seedling production. Open For. Sci. J 2009, 2, 17–24. [Google Scholar]
- Mejía-Teniente, L.; Torres-Pacheco, I.; González-Chavira, M.M.; Ocampo-Velázquez, R.V.; Herrera-Ruiz, G.; Chapa-Oliver, A.M.; Guevara-González, R.G. Use of elicitors as an approach for sustainable agriculture. Afr. J. Biotechnol 2010, 9, 9155–9162. [Google Scholar]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol 2012, 12, 89–100. [Google Scholar]
- Jones, D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar]
- Terry, L.A.; Joyce, D.C. Elicitors of induced disease resistance in postharvest horticultural crops: A brief review. Postharvest Biol. Technol 2004, 32, 1–13. [Google Scholar]
- García-Mier, L.; Guevara-González, R.G.; Mondragón-Olguín, V.M.; del Verduzco-Cuellar, B.R.; Torres-Pacheco, I. Agriculture and Bioactives: Achieving Both Crop Yield and Phytochemicals. Int. J. Mol. Sci 2013, 14, 4203–4222. [Google Scholar]
- Zinovieva, S.V.; Vasyukova, N.I.; Udalova, Z.V.; Gerasimova, N.G.; Ozeretskovskaya, O.L. Involvement of salicylic acid in induction of nematode resistance in plants. Biol. Bull 2011, 38, 453–458. [Google Scholar]
- Montesano, M.; Brader, G.; Palva, E.T. Pathogen derived elicitors: Searching for receptors in plants. Mol. Plant Pathol 2003, 4, 73–79. [Google Scholar]
- Jeong, G.-T.; Park, D.-H. Enhancement of growth and secondary metabolite biosynthesis: Effect of elicitors derived from plants and insects. Biotechnol. Bioprocess Eng 2005, 10, 73–77. [Google Scholar]
- Heredia, J.B.; Cisneros-Zevallos, L. The effects of exogenous ethylene and methyl jasmonate on the accumulation of phenolic antioxidants in selected whole and wounded fresh produce. Food Chem 2009, 115, 1500–1508. [Google Scholar]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J 1997, 11, 1187–1194. [Google Scholar]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat isease. Annu. Rev. Phytopathol 2009, 47, 177–206. [Google Scholar]
- Guevara-Olvera, L.; Ruíz-Nito, M.L.; Rangel-Cano, R.M.; Torres-Pacheco, I.; Rivera-Bustamante, R.F.; Muñoz-Sánchez, C.I.; González-Chavira, M.M.; Cruz-Hernandez, A.; Guevara-González, R.G. Expression of a germin-like protein gene (CchGLP) from a geminivirus-resistant pepper (Capsicum chinense Jacq.) enhances tolerance to geminivirus infection in transgenic tobacco. Physiol. Mol. Plant Pathol 2012, 78, 45–50. [Google Scholar]
- García-Neria, M.A.; Rivera-Bustamante, R. Characterization of Geminivirus Resistance in an Accession of Capsicum chinense Jacq. Mol. Plant. Microbe Interact 2011, 24, 172–182. [Google Scholar]
- Chong, T.M.; Abdullah, M.A.; Fadzillah, N.M.; Lai, O.M.; Lajis, N.H. Jasmonic acid elicitation of anthraquinones with some associated enzymic and non-enzymic antioxidant responses in Morinda elliptica. Enzyme Microb. Tech 2005, 36, 469–477. [Google Scholar]
- Mayers, C.N.; Lee, K.C.; Moore, C.A.; Wong, S.M.; Carr, J.P. Salicylic acid-induced resistance to Cucumber mosaic virus in squash and Arabidopsis thaliana: Contrasting mechanisms of induction and antiviral action. Mol. Plant Microbe. Interact 2005, 18, 428–434. [Google Scholar]
- Iseri, O.D.; Korpe, D.A.; Sahin, F.I.; Haberal, M. Hydrogen peroxide pretreatment of roots enhanced oxidative stress response of tomato under cold stress. Acta Physiol Plant 2013. [Google Scholar] [CrossRef]
- Desikan, R.; Reynolds, A.; Hancock, J.T.; Neill, S.J. Harpin and hydrogen peroxide both initiate programmed cell death but have differential effects on gene expression in Arabidopsis suspension cultures. Biochem. J 1998, 330, 115–120. [Google Scholar]
- Lin, W.; Hu, X.; Zhang, W.; Rogers, W.J.; Cai, W. Hydrogen peroxide mediates defence reponses induced by chitosans of different molecular weights in rice. J. Plant Physiol 2005, 162, 937–944. [Google Scholar]
- Liu, X.; Chi, H.; Yue, M.; Zhang, X.; Li, W.; Jia, E. The regulation of exogenous jasmonic acid on UV-B stress tolerance in Wheat. J Plant Growth Regul 2012, 31, 436–447. [Google Scholar]
- Airaki, M.; Leterrier, M.; Mateos, R.M.; Valderrama, R.; Chaki, M.; Barroso, J.B.; Del Río, L.A.; Palma, J.M.; Corpas, F.J. Metabolism of reactive oxygen species and reactive nitrogen species in pepper (Capsicum annuum L.) plants under low temperature stresspce. Plant Cell Environ 2012, 35, 281–295. [Google Scholar]
- Mutlu, S.; Karadagoglu, O.; Atici, O.; Nalbantoglu, B. Protective role of salicylic acid applied before cold stress on antioxidative system and protein patterns in barley apoplast. Biol. Plant 2013. [Google Scholar] [CrossRef]
- Vimala, R.; Suriachandraselvan, M. Induced resistance in bhendi against powdery mildew by foliar application of salicylic acid. J. Biopesticides 2009, 2, 111–114. [Google Scholar]
- Bautista-Banos, S.; Hernandez-Lauzardo, A.N.; del Velazquez-Valle, M.G. Chitosan as a potential natural compound to control pre and postharvest diseases of horticultural commodities. Crop Prot 2006, 25, 108–118. [Google Scholar]
- Tanabe, S.; Hayashi, N.; Nishizawa, Y.; Yamane, H.; Shibuya, N.; Minami, E. Elicitor and catalase activity of conidia suspensions of various strains of Magnaporthe grisea in suspensions-cultured cells of rice. Biosci. Biotechnol. Biochem 2008, 72, 889–892. [Google Scholar]
- Sahebani, N.; Hadavi, N. Induction of H2O2 and related enzymes in tomato roots infected with root knot nematode (M. javanica) by several chemical and microbial elicitors. Biocontrol. Sci. Technol 2009, 19, 301–313. [Google Scholar]
- Willekens, H.; Langebartelst, C.; Tiré, C.; Van Montagu, M.; Inzé, D.; van Camp, W. Differential expression of catalase genes in Nicotiana plumbaginifolia (L.). EMBO J 1994, 16, 4806–4816. [Google Scholar]
- Lee, S.H.; An, C.S. Differential expression of three catalase genes in hot pepper (Capsicum annuum L.). Mol. Cells 2005, 20, 247–255. [Google Scholar]
- Xu, M.; Dong, J. O2− from elicitor-induced oxidative burst is necessary for triggering phenylalanine ammonia-lyase activation and catharanthine synthesis in Catharanthus roseus cell cultures. Enzyme Microb. Technol 2005, 36, 280–284. [Google Scholar]
- Heath, M.C. Hypersensitive response-related death. Plant Mol. Biol 2000, 44, 321–334. [Google Scholar]
- Choi, H.W.; Kim, Y.J.; Lee, S.C.; Hong, J.K.; Hwang, B.K. Hydrogen peroxide generation by the pepper extracellular peroxidase CaPO2 activates local and systemic cell death and defense response to bacterial pathogens. Plant Physiol 2007, 145, 890–904. [Google Scholar]
- De Román, M.; Fernández, I.; Wyatt, T.; Sahrawy, M.; Heil, M.; Pozo, M.J. Elicitation of foliar resistance mechanisms transiently impairs root association with arbuscular mycorrhizal fungi. J. Ecol 2011, 99, 36–45. [Google Scholar]
- Huang, C.; Verrillo, F.; Renzone, G.; Arena, S.; Rocco, M.; Scalonic, A.; Marra, M. Response to biotic and oxidative stress in Arabidopsis thaliana: Analysis of variably phosphorylated proteins. J. Proteomics 2011, 74, 1934–1949. [Google Scholar]
- Steiner, A.A. The universal nutrient solution. Proceedings of 6th International Congress on Soilles Culture, Wageningen, The Netherlands, 29 April–5 May 1984; pp. 633–650.
- Iriti, M.; Faoro, F. Chitosan as a MAMP, searching for a PRR. Plant Signal Behav 2009, 4, 66–68. [Google Scholar]
- Tierranegra-García, N.; Salinas-Soto, P.; Torres-Pacheco, I.; Ocampo-Velázquez, R.V.; Rico-García, E.; Mendoza-Diaz, S.O.; Feregrino-Pérez, A.A.; Mercado-Luna, A.; Vargas-Hernandez, M.; Soto-Zarazúa, G.M.; et al. Effect of foliar salicylic acid and methyl jasmonate applications on protection against pill-bugs in lettuce plants (Lactuca sativa). Phytoparasitica 2011, 39, 137–144. [Google Scholar]
- Lizárraga-Paulín, E.G.; Torres-Pacheco, I.; Moreno-Martínez, E.; Miranda-Castro, S.P. Chitosan application in maize (Zea mays) to counteract the effects of abiotic stress at seedling level. Afr. J. Biotechnol 2011, 10, 6439–6446. [Google Scholar]
- Sibanda, T.; Okoh, A.I. In vitro evaluation of interactions between acetone extracts of Garcinia kola seeds and some antibiotics. Afr. J. Biotechnol 2008, 7, 1672–1678. [Google Scholar]
- Chandlee, J.M.; Tsaftaris, A.S.; Scandalios, J.G. Purification and partial characterization of three genetically defined catalases of maize. Plant Sci. Lett 1983, 29, 117–131. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Gerasimova, N.G.; Pridvorova, S.M.; Ozeretskovskaya, O.L. Role of l-phenylalanine ammonia lyase in the induced resistance and susceptibility of potato plants. Appl. Biochem. Microbiol 2005, 41, 103–105. [Google Scholar]
- JMP, version 5.0.1; Software for dynamic data visualisation and deep analytics; SAS Institute: Cary, NC, USA, 2002.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Oligonucleotide sequence (5′-3′) | Product size (bp) | Genbank accession No. |
|---|---|---|---|
| CaCAT-F | GTCCATGAGCGTGGAAGCCCCGAAT | 841 | AF227952 |
| CaCAT-R | CGCGATGCATGAAGTTCATGGCACC | ||
| CaPAL-F | TGGTGGATTTTTCGAGTTGCAGCCG | 831 | EU616575 |
| CaPAL-R | TGGCAAAGCGCCACGAGATAGGTTG | ||
| CaPR1-F | CTTTTGCTATATTTCACTCAACACAAGCCC | 522 | AF053343 |
| CaPR1-R | TGCTGGATTTATTTTCCTTTTAACACATGA | ||
| CaGPDH-F | GGCCTTATGACTACAGTTCACTCC | 255 | AJ246011 |
| CaGPDH-R | GATCAACCACAGAGACATCCACAG |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mejía-Teniente, L.; Durán-Flores, F.d.D.; Chapa-Oliver, A.M.; Torres-Pacheco, I.; Cruz-Hernández, A.; González-Chavira, M.M.; Ocampo-Velázquez, R.V.; Guevara-González, R.G. Oxidative and Molecular Responses in Capsicum annuum L. after Hydrogen Peroxide, Salicylic Acid and Chitosan Foliar Applications. Int. J. Mol. Sci. 2013, 14, 10178-10196. https://doi.org/10.3390/ijms140510178
Mejía-Teniente L, Durán-Flores FdD, Chapa-Oliver AM, Torres-Pacheco I, Cruz-Hernández A, González-Chavira MM, Ocampo-Velázquez RV, Guevara-González RG. Oxidative and Molecular Responses in Capsicum annuum L. after Hydrogen Peroxide, Salicylic Acid and Chitosan Foliar Applications. International Journal of Molecular Sciences. 2013; 14(5):10178-10196. https://doi.org/10.3390/ijms140510178
Chicago/Turabian StyleMejía-Teniente, Laura, Flor de Dalia Durán-Flores, Angela María Chapa-Oliver, Irineo Torres-Pacheco, Andrés Cruz-Hernández, Mario M. González-Chavira, Rosalía V. Ocampo-Velázquez, and Ramón G. Guevara-González. 2013. "Oxidative and Molecular Responses in Capsicum annuum L. after Hydrogen Peroxide, Salicylic Acid and Chitosan Foliar Applications" International Journal of Molecular Sciences 14, no. 5: 10178-10196. https://doi.org/10.3390/ijms140510178
APA StyleMejía-Teniente, L., Durán-Flores, F. d. D., Chapa-Oliver, A. M., Torres-Pacheco, I., Cruz-Hernández, A., González-Chavira, M. M., Ocampo-Velázquez, R. V., & Guevara-González, R. G. (2013). Oxidative and Molecular Responses in Capsicum annuum L. after Hydrogen Peroxide, Salicylic Acid and Chitosan Foliar Applications. International Journal of Molecular Sciences, 14(5), 10178-10196. https://doi.org/10.3390/ijms140510178