NS5ATP9 Contributes to Inhibition of Cell Proliferation by Hepatitis C Virus (HCV) Nonstructural Protein 5A (NS5A) via MEK/Extracellular Signal Regulated Kinase (ERK) Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. HCV NS5A Inhibited Proliferation of HCC Cell Lines
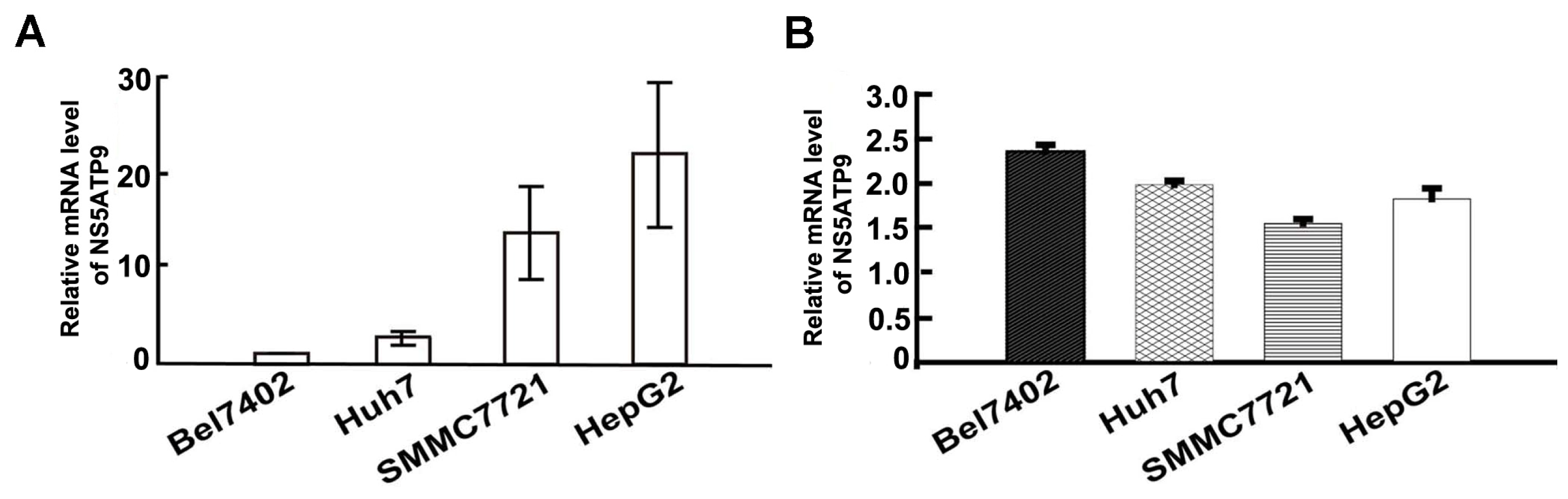
2.2. Four HCC Cell Lines Showed the Differential mRNA Levels of NS5ATP9
2.3. HCV NS5A Up-Regulated NS5ATP9 mRNA Levels in HCC Cell Lines
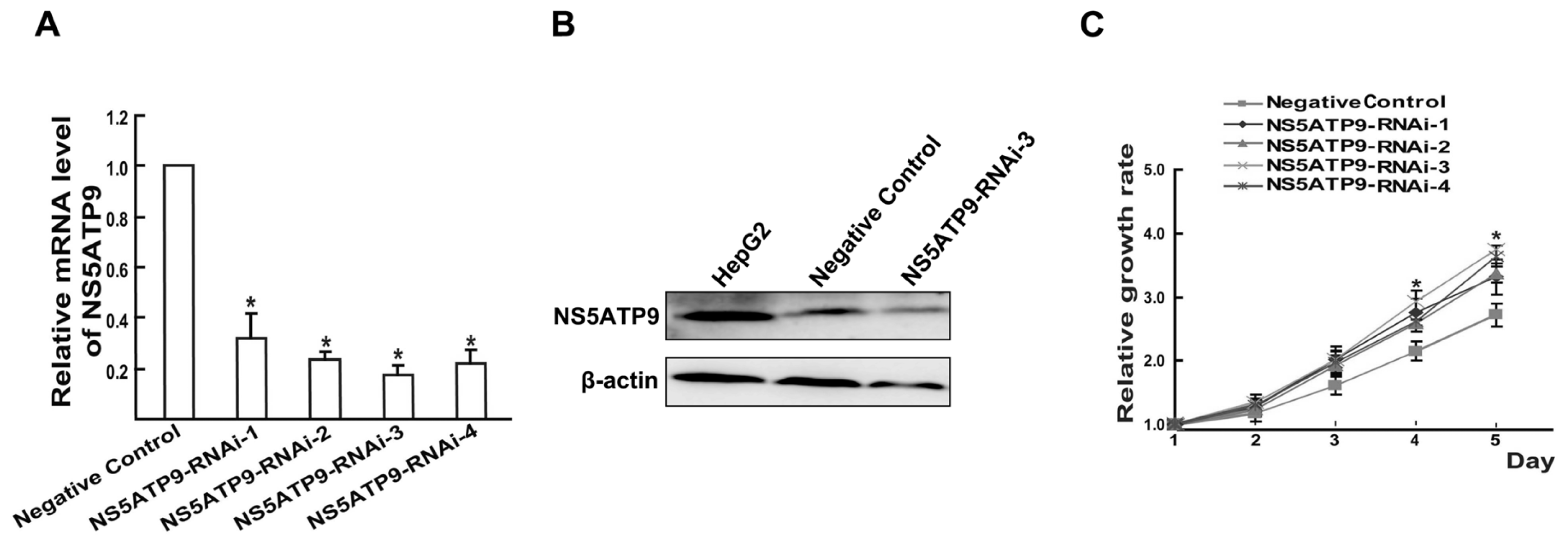
2.4. Overexpression and Knockdown of NS5ATP9 Showed the Reverse Effect on Cell Proliferation
2.5. NS5ATP9 Mediated the Inhibition of Proliferation under HCV NS5A Expression
2.6. NS5ATP9 Knockdown Activated MEK/ERK Signaling Pathway under HCV NS5A Expression
3. Experimental Section
3.1. Cell Culture and Transient Transfection
3.2. Expression and RNAi Plasmids Construction
- NS5ATP9-RNAi-1(5′-TGCTGAACACTGTCTGCTTTAGTCCGGTTTTGGCCACTGACTGACCG GACTAACAGACAGTGTT-3′);
- NS5ATP9-RNAi-2(5′-TGCTGTTCTGTAAGTGCCTGGAACACGTTTTGGCCACTGACTGACGT GTTCCACACTTACAGAA-3′);
- NS5ATP9-RNAi-3(5′-TGCTGACTGATGTCGAATTAGTGGCAGTTTTGGCCACTGACTGACTG CCACTATCGACATCAGT-3′);
- NS5ATP9-RNAi-4(5′-TGCTGTTTCCTAAGCCACTGCTTCCTGTTTTGGCCACTGACTGACAGG AAGC AGG CTTAG GAAA-3′).
3.3. Real Time PCR Analysis
3.4. Cell Viability Assay
3.5. Western Blot
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
Abbreviations
| HCV NS5A | Hepatitis C virus nonstructural protein 5A |
| HCC | hepatocellular carcinoma |
| RNAi | RNA interference |
| β-actin | beta-actin |
| MAPK | mitogen-activated protein kinase |
| MEK | mitogen-activated protein kinase kinase |
| ERK | extracellular signal regulated kinase. |
References
- Krieger, N.; Lohmann, V.; Bartenschlager, R. Enhancement of hepatitis C virus RNA replication by cell culture-adaptive mutations. J. Virol 2001, 75, 4614–4624. [Google Scholar]
- Kiyosawa, K.; Sodeyama, T.; Tanaka, E.; Gibo, Y.; Yoshizawa, K.; Nakano, Y.; Furuta, S.; Akahane, Y.; Nishioka, K.; Purcell, R.H.; et al. Interrelationship of blood transfusion, non-A, non-B hepatitis and hepatocellular carcinoma: Analysis by detection of antibody to hepatitis C virus. Hepatology 1990, 12, 671–675. [Google Scholar]
- Takamizawa, A.; Mori, C.; Fuke, I.; Manabe, S.; Murakami, S.; Fujita, J.; Onishi, E.; Andoh, T.; Yoshida, I.; Okayama, H. Structure and organization of the hepatitis C virus genome isolated from human carriers. J. Virol 1991, 65, 1105–1113. [Google Scholar]
- Lim, Y.S.; Hwang, S.B. Hepatitis C virus NS5A protein interacts with phosphatidylinositol 4-kinase type IIIalpha and regulates viral propagation. J. Biol. Chem 2011, 286, 11290–11298. [Google Scholar]
- Reed, K.E.; Xu, J.; Rice, C.M. Phosphorylation of the hepatitis C virus NS5A protein in vitro and in vivo: Properties of the NS5A-associated kinase. J. Virol 1997, 71, 7187–7197. [Google Scholar]
- Barber, G.N.; Blight, K.J.; Carney, D.S.; Chevaliez, S.; Couto, L.B., Jr.; Gale, M.; Dubuisson, J.; Frick, D.N.; Glenn, J.S.; Goffard, A.; et al. Hepatitis C Viruses: Genomes and Molecular Biology; Horizon Bioscience: Norfolk, UK, 2006; Volume Chapter 9, pp. 267–292. [Google Scholar]
- Gimenez-Barcons, M.; Wang, C.; Chen, M.; Sanchez-Tapias, J.M.; Saiz, J.C.; Gale, M., Jr. The oncogenic potential of hepatitis C virus NS5A sequence variants is associated with PKR regulation. J. Interferon Cytokine Res 2005, 25, 152–164. [Google Scholar]
- Ghosh, A.K.; Majumder, M.; Steele, R.; Yaciuk, P.; Chrivia, J.; Ray, R.; Ray, R.B. Hepatitis C virus NS5A protein modulates transcription through a novel cellular transcription factor SRCAP. J. Biol. Chem 2000, 275, 7184–7188. [Google Scholar]
- Gong, G.Z.; Jiang, Y.F.; He, Y.; Lai, L.Y.; Zhu, Y.H.; Su, X.S. HCV NS5A abrogates p53 protein function by interfering with p53-DNA binding. World J. Gastroenterol 2004, 10, 2223–2227. [Google Scholar]
- Lan, K.H.; Sheu, M.L.; Hwang, S.J.; Yen, S.H.; Chen, S.Y.; Wu, J.C.; Wang, Y.J.; Kato, N.; Omata, M.; Chang, F.Y.; et al. HCV NS5A interacts with p53 and inhibits p53-mediated apoptosis. Oncogene 2002, 21, 4801–4811. [Google Scholar]
- Qadri, I.; Iwahashi, M.; Simon, F. Hepatitis C virus NS5A protein binds TBP and p53, inhibiting their DNA binding and p53 interactions with TBP and ERCC3. Biochim. Biophys. Acta 2002, 1592, 193–204. [Google Scholar]
- Siavoshian, S.; Abraham, J.D.; Kieny, M.P.; Schuster, C. HCV core, NS3, NS5A and NS5B proteins modulate cell proliferation independently from p53 expression in hepatocarcinoma cell lines. Arch. Virol 2004, 149, 323–336. [Google Scholar]
- Arima, N.; Kao, C.Y.; Licht, T.; Padmanabhan, R.; Sasaguri, Y.; Padmanabhan, R. Modulation of cell growth by the hepatitis C virus nonstructural protein NS5A. J. Biol. Chem 2001, 276, 12675–12684. [Google Scholar]
- Yu, P.; Huang, B.; Shen, M.; Lau, C.; Chan, E.; Michel, J.; Xiong, Y.; Payan, D.G.; Luo, Y. p15(PAF), a novel PCNA associated factor with increased expression in tumor tissues. Oncogene 2001, 20, 484–489. [Google Scholar]
- Yuan, R.H.; Jeng, Y.M.; Pan, H.W.; Hu, F.C.; Lai, P.L.; Lee, P.H.; Hsu, H.C. Overexpression of KIAA0101 predicts high stage, early tumor recurrence, and poor prognosis of hepatocellular carcinoma. Clin. Cancer Res 2007, 13, 5368–5376. [Google Scholar]
- Mizutani, K.; Onda, M.; Asaka, S.; Akaishi, J.; Miyamoto, S.; Yoshida, A.; Nagahama, M.; Ito, K.; Emi, M. Overexpressed in anaplastic thyroid carcinoma-1 (OEATC-1) as a novel gene responsible for anaplastic thyroid carcinoma. Cancer 2005, 103, 1785–1790. [Google Scholar]
- Hosokawa, M.; Takehara, A.; Matsuda, K.; Eguchi, H.; Ohigashi, H.; Ishikawa, O.; Shinomura, Y.; Imai, K.; Nakamura, Y.; Nakagawa, H. Oncogenic role of KIAA0101 interacting with proliferating cell nuclear antigen in pancreatic cancer. Cancer Res 2007, 67, 2568–2576. [Google Scholar]
- Guo, M.; Li, J.; Wan, D.; Gu, J. KIAA0101 (OEACT-1), an expressionally down-regulated and growth-inhibitory gene in human hepatocellular carcinoma. BMC Cancer 2006, 6, 109. [Google Scholar]
- Kato, T.; Furusaka, A.; Miyamoto, M.; Date, T.; Yasui, K.; Hiramoto, J.; Nagayama, K.; Tanaka, T.; Wakita, T. Sequence analysis of hepatitis C virus isolated from a fulminant hepatitis patient. J. Med. Virol 2001, 64, 334–339. [Google Scholar]
- Liu, S.; Xiao, L.; Nelson, C.; Hagedorn, C. A cell culture adapted HCV JFH1 variant that increases viral titers and permits the production of high titer infectious chimeric reporter viruses. PLoS One 2012, 7, e44965. [Google Scholar]
- Liu, L.; Chen, X.; Xie, S.; Zhang, C.; Qiu, Z.; Zhu, F. Variant 1 of KIAA0101, overexpressed in hepatocellular carcinoma, prevents doxorubicin-induced apoptosis by inhibiting p53 activation. Hepatology 2012, 56, 1760–1769. [Google Scholar]
- Petroziello, J.; Yamane, A.; Westendorf, L.; Thompson, M.; McDonagh, C.; Cerveny, C.; Law, C.L.; Wahl, A.; Carter, P. Suppression subtractive hybridization and expression profiling identifies a unique set of genes overexpressed in non-small-cell lung cancer. Oncogene 2004, 23, 7734–7745. [Google Scholar]
- Simpson, F.; van Lammerts, B.K.; Butterfield, N.; Bennetts, J.S.; Bowles, J.; Adolphe, C.; Simms, L.A.; Young, J.; Walsh, M.D.; Leggett, B.; et al. The PCNA-associated factor KIAA0101/p15(PAF) binds the potential tumor suppressor product p33ING1b. Exp. Cell Res 2006, 312, 73–85. [Google Scholar]
- Jain, M.; Zhang, L.; Patterson, E.E.; Kebebew, E. KIAA0101 is overexpressed, and promotes growth and invasion in adrenal cancer. PLoS One 2011, 6, e26866. [Google Scholar]
- Emanuele, M.J.; Ciccia, A.; Elia, A.E.; Elledge, S.J. Proliferating cell nuclear antigen (PCNA)-associated KIAA0101/PAF15 protein is a cell cycle-regulated anaphase-promoting complex/cyclosome substrate. Proc. Natl. Acad. Sci. USA 2011, 108, 9845–9850. [Google Scholar]
- Kato, T.; Daigo, Y.; Aragaki, M.; Ishikawa, K.; Sato, M.; Kaji, M. Overexpression of KIAA0101 predicts poor prognosis in primary lung cancer patients. Lung Cancer 2012, 75, 110–118. [Google Scholar]
- Shi, L.; Zhang, S.L.; Li, K.; Hong, Y.; Wang, Q.; Li, Y.; Guo, J.; Fan, W.H.; Zhang, L.; Cheng, J. NS5ATP9, a gene up-regulated by HCV NS5A protein. Cancer Lett 2008, 259, 192–197. [Google Scholar]
- Li, K.; Ma, Q.; Shi, L.; Dang, C.; Hong, Y.; Wang, Q.; Li, Y.; Fan, W.; Zhang, L.; Cheng, J. NS5ATP9 gene regulated by NF-kappaB signal pathway. Arch. Biochem. Biophys 2008, 479, 15–19. [Google Scholar]
- Roberts, P.J.; Der, C.J. Targeting the Raf-MEK-ERK mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene 2007, 26, 3291–3310. [Google Scholar]
- Georgopoulou, U.; Caravokiri, K.; Mavromara, P. Suppression of the ERK1/2 signaling pathway from HCV NS5A protein expressed by herpes simplex recombinant viruses. Arch. Virol 2003, 148, 237–251. [Google Scholar]
- Nakabayashi, H.; Taketa, K.; Miyano, K.; Yamane, T.; Sato, J. Growth of human hepatoma cells lines with differentiated functions in chemically defined medium. Cancer Res 1982, 42, 3858–3863. [Google Scholar]
- Shen, D.W.; Lu, Y.G.; Chin, K.V.; Pastan, I.; Gottesman, M.M. Human hepatocellular carcinoma cell lines exhibit multidrug resistance unrelated to MRD1 gene expression. J. Cell. Sci 1991, 98, 317–322. [Google Scholar]
- Morris, K.M.; Aden, D.P.; Knowles, B.B.; Colten, H.R. Complement biosynthesis by the human hepatoma-derived cell line HepG2. J. Clin. Invest 1982, 70, 906–913. [Google Scholar]
- Invitrogen’s online RNAi Designer. Available online: http://www.invitrogen.com/rnai (accessed on 26 April 2013).






© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Q.; Wang, Y.; Li, Y.; Gao, X.; Liu, S.; Cheng, J. NS5ATP9 Contributes to Inhibition of Cell Proliferation by Hepatitis C Virus (HCV) Nonstructural Protein 5A (NS5A) via MEK/Extracellular Signal Regulated Kinase (ERK) Pathway. Int. J. Mol. Sci. 2013, 14, 10539-10551. https://doi.org/10.3390/ijms140510539
Wang Q, Wang Y, Li Y, Gao X, Liu S, Cheng J. NS5ATP9 Contributes to Inhibition of Cell Proliferation by Hepatitis C Virus (HCV) Nonstructural Protein 5A (NS5A) via MEK/Extracellular Signal Regulated Kinase (ERK) Pathway. International Journal of Molecular Sciences. 2013; 14(5):10539-10551. https://doi.org/10.3390/ijms140510539
Chicago/Turabian StyleWang, Qi, Yongsheng Wang, Yue Li, Xuesong Gao, Shunai Liu, and Jun Cheng. 2013. "NS5ATP9 Contributes to Inhibition of Cell Proliferation by Hepatitis C Virus (HCV) Nonstructural Protein 5A (NS5A) via MEK/Extracellular Signal Regulated Kinase (ERK) Pathway" International Journal of Molecular Sciences 14, no. 5: 10539-10551. https://doi.org/10.3390/ijms140510539
APA StyleWang, Q., Wang, Y., Li, Y., Gao, X., Liu, S., & Cheng, J. (2013). NS5ATP9 Contributes to Inhibition of Cell Proliferation by Hepatitis C Virus (HCV) Nonstructural Protein 5A (NS5A) via MEK/Extracellular Signal Regulated Kinase (ERK) Pathway. International Journal of Molecular Sciences, 14(5), 10539-10551. https://doi.org/10.3390/ijms140510539