p38β, A Novel Regulatory Target of Pokemon in Hepatic Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
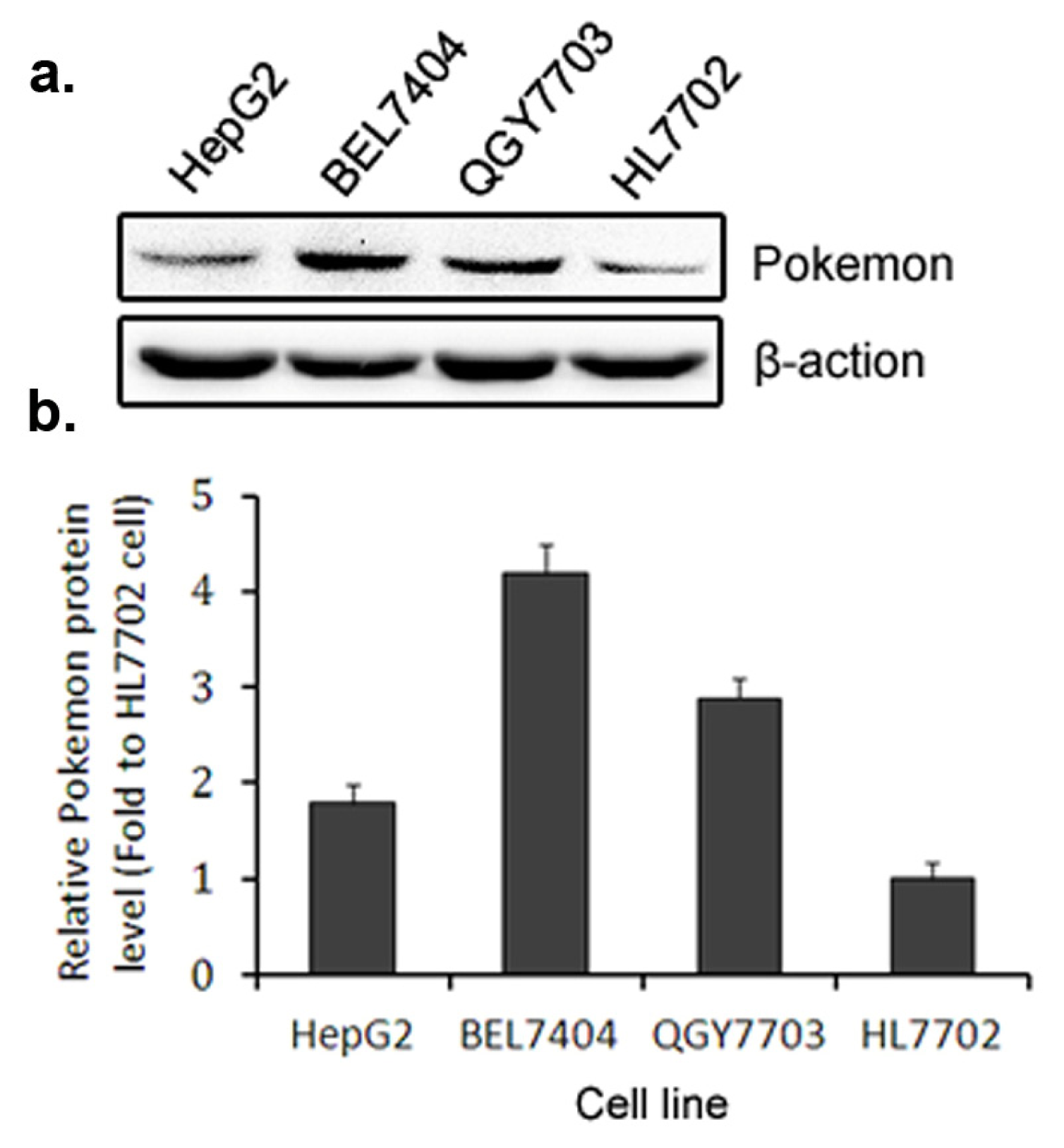
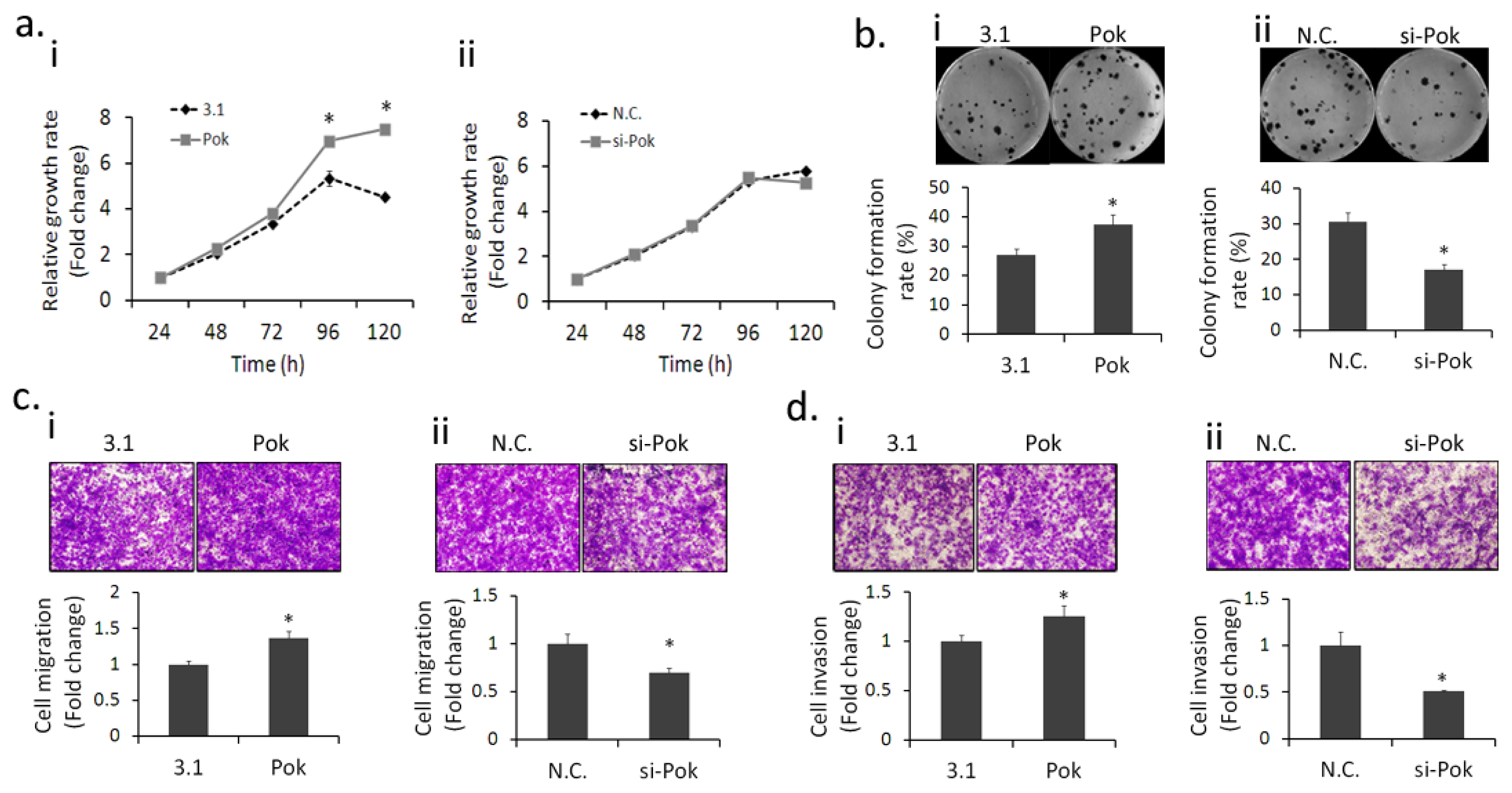
2.1. Pokemon Promotes HepG2 Cell Growth, Migration and Invasion
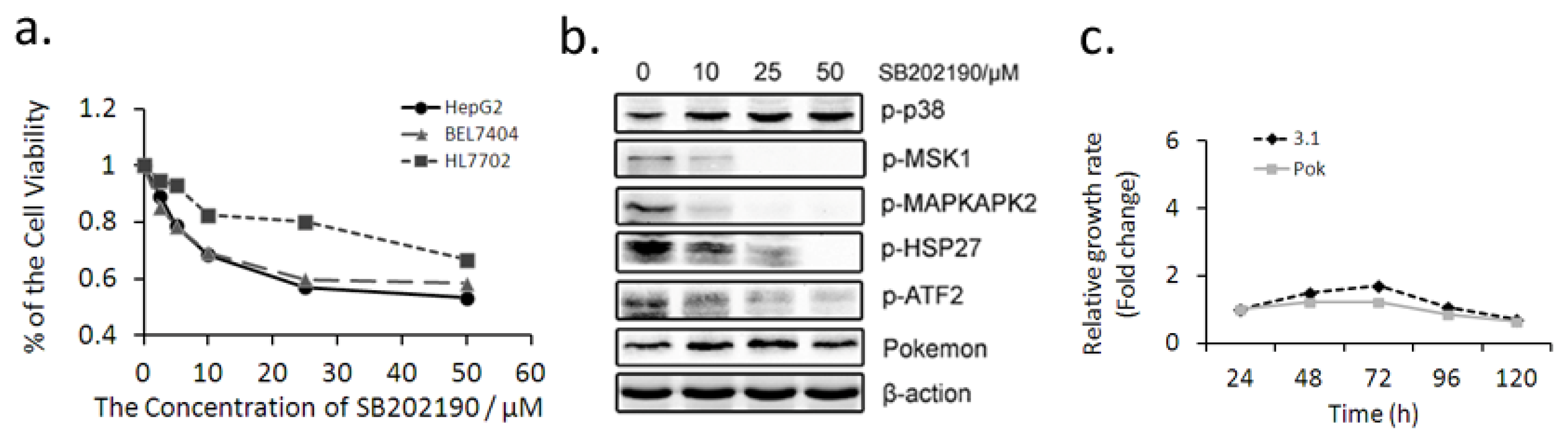
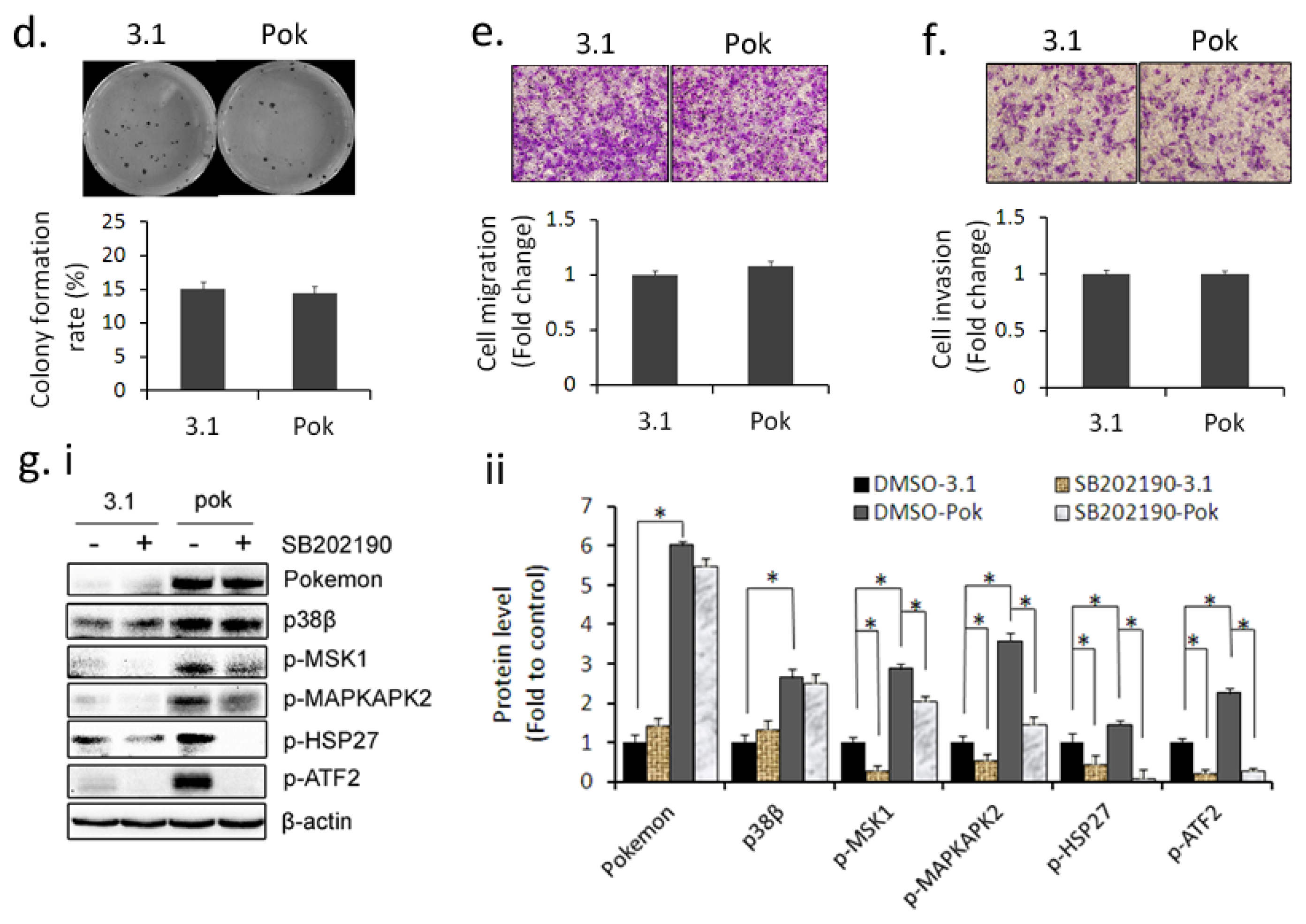
2.2. p38 Inhibitor SB202190 Attenuates the Promotion of Pokemon on HepG2 Growth, Migration and Invasion
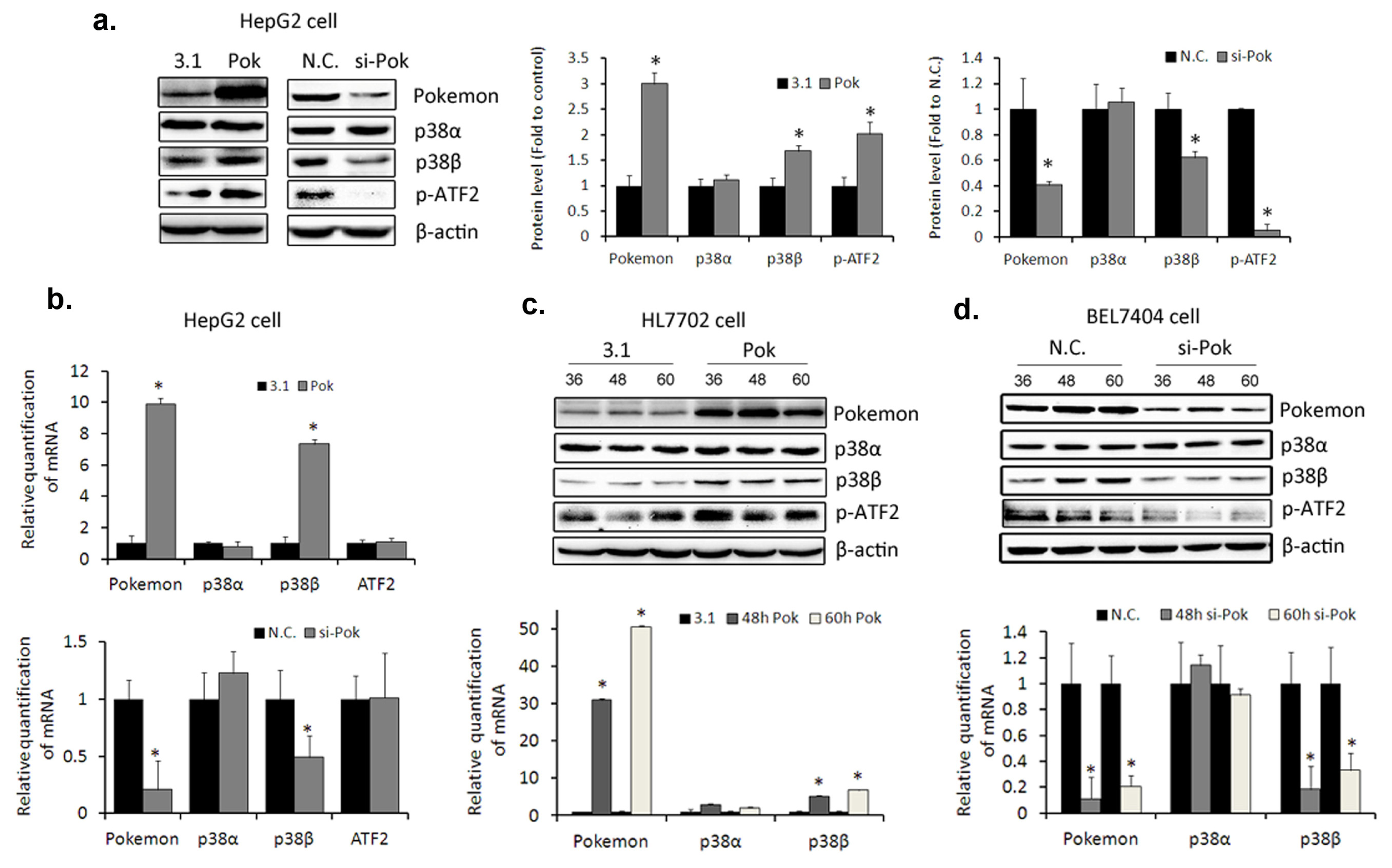
2.3. Pokemon Up-Regulates p38β Expression in Hepatic Cells
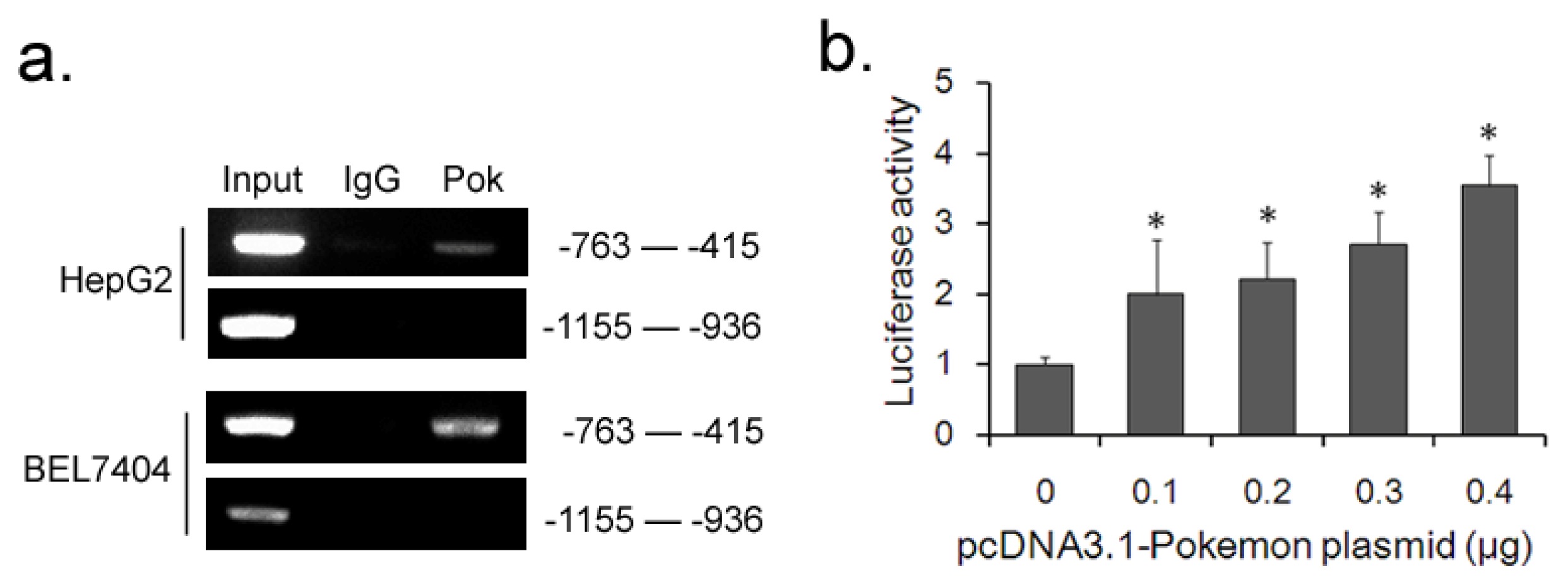
2.4. Pokemon Promotes p38β Expression by Up-Regulating Its Transcription
2.5. Discussion
3. Experimental Section
3.1. Cell Culture
3.2. Real-Time Quantitative PCR
3.3. Western Blot
3.4. Dual Luciferase Reporter Assay
3.5. Chromatin Immunoprepcipitation (ChIP)
3.6. MTT Assay
3.7. Colony Formation Assay
3.8. Invasion and Migration Assays
3.9. Statistical Analysis
4. Conclusions
Supplementary Information
ijms-14-13511-s001.pdfAcknowledgments
Conflict of Interest
References
- Pessler, F.; Pendergrast, P.S.; Hernandez, N. Purification and characterization of FBI-1, a cellular factor that binds to the human immunodeficiency virus type 1 inducer of short transcripts. Mol. Cell Biol 1997, 17, 3786–3798. [Google Scholar]
- Maeda, T.; Ito, K.; Merghoub, T.; Poliseno, L.; Hobbs, R.M.; Wang, G.; Dong, L.; Maeda, M.; Dore, L.C.; Zelent, A.; et al. LRF is an essential downstream target of GATA1 in erythroid development and regulates BIM-dependent apoptosis. Dev. Cell 2009, 17, 527–540. [Google Scholar]
- Maeda, T.; Merghoub, T.; Hobbs, R.M.; Dong, L.; Maeda, M.; Zakrzewski, J.; van den Brink, M.R.; Zelent, A.; Shigematsu, H.; Akashi, K.; et al. Regulation of B versus T lymphoid lineage fate decision by the proto-oncogene LRF. Science 2007, 316, 860–866. [Google Scholar]
- Collins, T.; Stone, J.R.; Williams, A.J. All in the family: The BTB/POZ, KRAB, and SCAN domains. Mol. Cell Biol 2001, 21, 3609–3615. [Google Scholar]
- Maeda, T.; Hobbs, R.M.; Merghoub, T.; Guernah, I.; Zelent, A.; Cordon-Cardo, C.; Teruya-Feldstein, J.; Pandolfi, P.P. Role of the proto-oncogene Pokemon in cellular transformation and ARF repression. Nature 2005, 433, 278–285. [Google Scholar]
- Weinberg, R.A. The cat and mouse games that genes, viruses, and cells play. Cell 1997, 88, 573–575. [Google Scholar]
- Aggarwal, H.; Aggarwal, A.; Hunter, W.J.; Yohannes, P.; Khan, A.U.; Agrawal, D.K. Expression of leukemia/lymphoma related factor (LRF/Pokemon) in human benign prostate hyperplasia and prostate cancer. Exp. Mol. Pathol 2011, 90, 226–230. [Google Scholar]
- Aggarwal, A.; Hunter, W.J.; Aggarwal, H.; Silva, E.D.; Davey, M.S.; Murphy, R.F.; Agrawal, D.K. Expression of leukemia/lymphoma-related factor (LRF/POKEMON) in human breast carcinoma and other cancers. Exp. Mol. Pathol 2010, 89, 140–148. [Google Scholar]
- Zu, X.; Ma, J.; Liu, H.; Liu, F.; Tan, C.; Yu, L.; Wang, J.; Xie, Z.; Cao, D.; Jiang, Y. Pro-oncogene Pokemon promotes breast cancer progression by upregulating survivin expression. Breast Cancer Res. 2011, 13. [Google Scholar] [CrossRef]
- Rovin, R.A.; Winn, R. Pokemon expression in malignant glioma: An application of bioinformatics methods. Neurosurg. Focus 2005, 19, E8. [Google Scholar]
- Nakayama, K.; Nakayama, N.; Wang, T.L.; Shih, I.M. NAC-1 controls cell growth and survival by repressing transcription of Gadd45GIP1, a candidate tumor suppressor. Cancer Res 2007, 67, 8058–8064. [Google Scholar]
- Choi, W.I.; Jeon, B.N.; Park, H.; Yoo, J.Y.; Kim, Y.S.; Koh, D.I.; Kim, M.H.; Kim, Y.R.; Lee, C.E.; Kim, K.S.; et al. Proto-oncogene FBI-1 (Pokemon) and SREBP-1 synergistically activate transcription of fatty-acid synthase gene (FASN). J. Biol. Chem 2008, 283, 29341–29354. [Google Scholar]
- Choi, W.I.; Jeon, B.N.; Yun, C.O.; Kim, P.H.; Kim, S.E.; Choi, K.Y.; Kim, S.H.; Hur, M.W. Proto-oncogene FBI-1 represses transcription of p21CIP1 by inhibition of transcription activation by p53 and Sp1. J. Biol. Chem 2009, 284, 12633–12644. [Google Scholar]
- Jeon, B.N.; Yoo, J.Y.; Choi, W.I.; Lee, C.E.; Yoon, H.G.; Hur, M.W. Proto-oncogene FBI-1 (Pokemon/ZBTB7A) represses transcription of the tumor suppressor Rb gene via binding competition with Sp1 and recruitment of co-repressors. J. Biol. Chem 2008, 283, 33199–33210. [Google Scholar]
- Zhang, N.N.; Sun, Q.S.; Chen, Z.; Liu, F.; Jiang, Y.Y. Homeostatic regulatory role of Pokemon in NF-κB signaling: Stimulating both p65 and IκBα expression in human hepatocellular carcinoma cells. Mol. Cell Biochem 2012, 372, 57–64. [Google Scholar]
- Lee, D.K.; Kang, J.E.; Park, H.J.; Kim, M.H.; Yim, T.H.; Kim, J.M.; Heo, M.K.; Kim, K.Y.; Kwon, H.J.; Hur, M.W. FBI-1 enhances transcription of the nuclear factor-κB (NF-κB)-responsive E-selectin gene by nuclear localization of the p65 subunit of NF-κB. J. Biol. Chem 2005, 280, 27783–27791. [Google Scholar]
- Liu, K.; Liu, F.; Zhang, N.N.; Liu, S.Y.; Jiang, Y.Y. Pokemon Silencing leads to Bim-mediated Anoikis of Human Hepatoma Cell QGY7703. Int. J. Mol. Sci 2012, 13, 5818–5831. [Google Scholar]
- Nebreda, A.R.; Porras, A. p38 MAP kinases: Beyond the stress response. Trends Biochem. Sci 2000, 25, 257–260. [Google Scholar]
- Ono, K.; Han, J. The p38 signal transduction pathway: Activation and function. Cell Signal 2000, 12, 1–13. [Google Scholar]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev 2001, 81, 807–869. [Google Scholar]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochem. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar]
- Lee, J.C.; Laydon, J.T.; McDonnell, P.C.; Gallagher, T.F.; Kumar, S.; Green, D.; McNulty, D.; Blumenthal, M.J.; Heys, J.R.; Landvatter, S.W.; et al. A protein kinase involved in the regulation of inflammatory cytokine biosynthesis. Nature 1994, 372, 739–746. [Google Scholar]
- Wagner, E.F.; Nebreda, A.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar]
- Xu, L.; Chen, S.; Bergan, R.C. MAPKAPK2 and HSP27 are downstream effectors of p38 MAP kinase-mediated matrix metalloproteinase type 2 activation and cell invasion in human prostate cancer. Oncogene 2006, 25, 2987–2998. [Google Scholar]
- Deak, M.; Clifton, A.D.; Lucocq, L.M.; Alessi, D.R. Mitogen- and stress-activated protein kinase-1 (MSK1) is directly activated by MAPK and SAPK2/p38, and may mediate activation of CREB. EMBO J 1998, 17, 4426–4441. [Google Scholar]
- Patel, S.B.; Cameron, P.M.; O’Keefe, S.J.; Frantz-Wattley, B.; Thompson, J.; O’Neill, E.A.; Tennis, T.; Liu, L.; Becker, J.W.; Scapin, G. The three-dimensional structure of MAP kinase p38β: Different features of the ATP-binding site in p38β compared with p38α. Acta Crystallogr. D 2009, 65, 777–785. [Google Scholar]
- Jiang, Y.; Chen, C.; Li, Z.; Guo, W.; Gegner, J.A.; Lin, S.; Han, J. Characterization of the structure and function of a new mitogen-activated protein kinase (p38β). J. Biol. Chem 1996, 271, 17920–17926. [Google Scholar]
- Guo, Y.L.; Kang, B.; Han, J.; Williamson, J.R. p38β MAP kinase protects rat mesangial cells from TNF-α-induced apoptosis. J. Cell Biochem 2001, 82, 556–565. [Google Scholar]
- Kim, H.P.; Wang, X.; Nakao, A.; Kim, S.I.; Murase, N.; Choi, M.E.; Ryter, S.W.; Choi, A.M. Caveolin-1 expression by means of p38β mitogen-activated protein kinase mediates the antiproliferative effect of carbon monoxide. Proc. Natl. Acad. Sci. USA 2005, 102, 11319–11324. [Google Scholar]
- Kalender, A.; Selvaraj, A.; Thomas, G. A matter of energy stress: p38β meets mTORC1. Cell Res 2011, 21, 859–861. [Google Scholar]
- Jiang, L.; Siu, M.K.; Wong, O.G.; Tam, K.F.; Lam, E.W.; Ngan, H.Y.; Le, X.F.; Wong, E.S.; Chan, H.Y.; Cheung, A.N. Overexpression of proto-oncogene FBI-1 activates membrane type 1-matrix metalloproteinase in association with adverse outcome in ovarian cancers. Mol. Cancer 2010, 9. [Google Scholar] [CrossRef] [Green Version]
- Nemoto, S.; Xiang, J.; Huang, S.; Lin, A. Induction of apoptosis by SB202190 through inhibition of p38β mitogen-activated protein kinase. J. Biol. Chem 1998, 273, 16415–16420. [Google Scholar]
- Zu, X.; Yu, L.L.; Sun, Y.; Tian, J.; Liu, F.; Sun, Q.S.; He, S.N.; Sun, G.; Luo, W.S.; Jiang, Y.Y. Global mapping of ZBTB7A transcription factor binding sites in HepG2 cells. Cell Mol. Biol. Lett 2010, 15, 260–271. [Google Scholar]






© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, Z.; Liu, F.; Zhang, N.; Cao, D.; Liu, M.; Tan, Y.; Jiang, Y. p38β, A Novel Regulatory Target of Pokemon in Hepatic Cells. Int. J. Mol. Sci. 2013, 14, 13511-13524. https://doi.org/10.3390/ijms140713511
Chen Z, Liu F, Zhang N, Cao D, Liu M, Tan Y, Jiang Y. p38β, A Novel Regulatory Target of Pokemon in Hepatic Cells. International Journal of Molecular Sciences. 2013; 14(7):13511-13524. https://doi.org/10.3390/ijms140713511
Chicago/Turabian StyleChen, Zhe, Feng Liu, Nannan Zhang, Deliang Cao, Min Liu, Ying Tan, and Yuyang Jiang. 2013. "p38β, A Novel Regulatory Target of Pokemon in Hepatic Cells" International Journal of Molecular Sciences 14, no. 7: 13511-13524. https://doi.org/10.3390/ijms140713511
APA StyleChen, Z., Liu, F., Zhang, N., Cao, D., Liu, M., Tan, Y., & Jiang, Y. (2013). p38β, A Novel Regulatory Target of Pokemon in Hepatic Cells. International Journal of Molecular Sciences, 14(7), 13511-13524. https://doi.org/10.3390/ijms140713511