The Interstitial Interface within the Renal Stem/Progenitor Cell Niche Exhibits an Unique Microheterogeneous Composition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Vertical Sight of the Renal Stem/Progenitor Cell Niche
2.2. Light Microscopical View to the Renal Stem/Progenitor Cell Niche
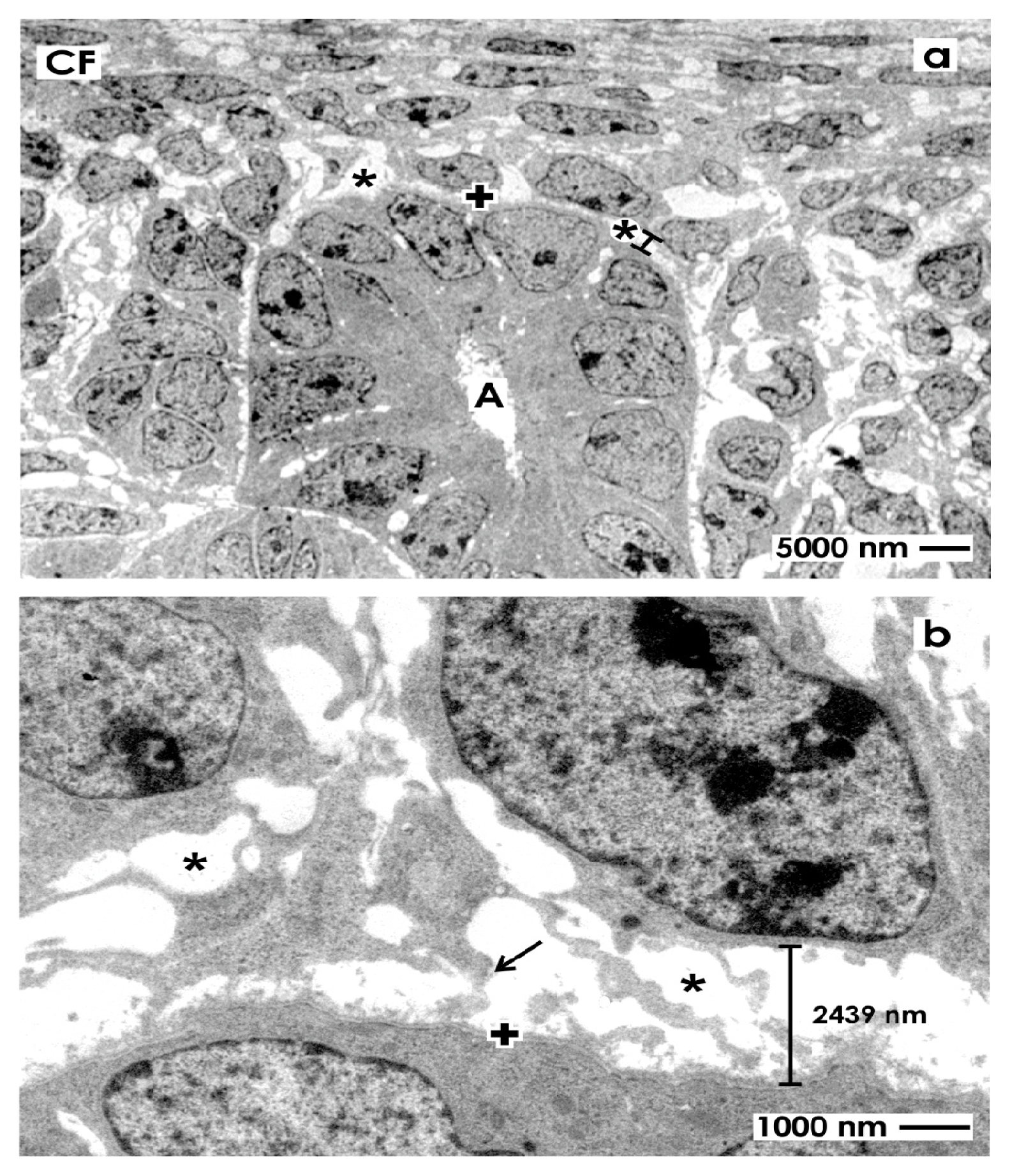
2.3. Electron Microscopical View to the Renal Stem/Progenitor Cell Niche
2.3.1. Traditional Fixation with Glutaraldehyde (GA)
2.3.2. Contrasting with GA Including Cupromeronic Blue
2.3.3. Contrasting with GA Including Ruthenium Red
2.3.4. Contrasting with GA Including Tannic Acid
2.3.5. Site-Specific Reaction of Contrasting
2.4. Offering a Suitable Environment for Initial Repair
3. Experimental Program and Materials
3.1. Tissue Preparation
3.2. Light Microscopy
3.3. Transmission Electron Microscopy
- Specimens for control: 5% GA (Serva, Heidelberg, Germany) buffered with 0.15 M sodium cacodylate, pH 7.4.
- Series with cupromeronic blue: 5% GA buffered with 0.15 M sodium cacodylate, pH 7.4. Then specimens were incubated in 0.1% cupromeronic blue (Santa Cruz, Heidelberg, Germany) and 0.1 M magnesium chloride hexahydrate (Sigma, Taufkirchen, Germany) dissolved in sodium acetate buffer pH 5.6. Counterstaining was performed with 0.5% sodium tungstate dehydrate (Sigma).
- Series with ruthenium red: 5% GA buffered with 0.15 M sodium cacodylate, pH 7.4 + 0.5% ruthenium red (Fluka, Taufkirchen, Germany).
- Series with tannic acid: 5% glutaraldehyde buffered with 0.15 M sodium cacodylate, pH 7.4 + 1% tannic acid (Sigma).
3.4. Amount of Analyzed Specimens
3.5. Definition of Cells within the Renal Stem/Progenitor Cell Niche
4. Conclusions
Conflict of Interest
References
- Thompson, S.; Pannu, N. Renal replacement therapy in end-stage renal disease patient with critical illness. Blood Purif 2012, 34, 132–137. [Google Scholar]
- Fuquay, R.; Teitelbaum, I. Care of the patient after renal allograft failure: Managing the present and planning for the future. Am. J. Nephrol 2012, 36, 348–354. [Google Scholar]
- Issa, N.; Lankireddy, S.; Kukla, A. Should peritoneal dialysis be the preferred therapy pre-kidney transplantation? Adv. Perit. Dial 2012, 28, 89–93. [Google Scholar]
- Caldas, H.C.; Hayashi, A.P.; Abbud-Filho, M. Repairing the chronic damaged kidney: The role of regenerative medicine. Transplant. Proc 2011, 43, 3573–3576. [Google Scholar]
- Burst, V.; Pütsch, F.; Kubacki, T.; Völker, L.A.; Bartram, M.P.; Müller, R.U.; Gillis, M.; Kurschat, C.E.; Grundmann, F.; Müller-Ehmsen, J.; et al. Survival and distribution of injected haematopoietic stem cells in acute kidney injury. Nephrol. Dial. Transplant 2012, 28, 1131–1139. [Google Scholar]
- Faa, G.; Gerosa, C.; Fanni, D.; Monga, G.; Zaffanello, M.; van Eyken, P.; Fanos, V. Morphogenesis and molecular mechanisms involved in human kidney development. J. Cell. Physiol 2012, 227, 1257–1268. [Google Scholar]
- Minuth, W.; Denk, L.; Miess, C.; Glashauser, A. Peculiarities of the extracellular matrix in the interstitium of the renal stem/progenitor cell niche. Histochem. Cell Biol 2011, 136, 321–334. [Google Scholar]
- Pleniceanu, O.; Dekel, B. Stem Cells in Fetal Tissue (The Kidney as a Model). In Human Fetal Tissue Transplantation; Bhattachary, N., Stubblefiled, P., Eds.; Springer Verlag: London, UK, 2013; pp. 121–140. [Google Scholar]
- Little, M.H. Renal organanogenesis: What can it tell us about renal repair and regeneration? Organogenesis 2011, 7, 229–241. [Google Scholar]
- Anglani, F.; Mezzebotta, F.; Ceol, M.; Cristofaro, R.; del Prete, D.; D’Angelo, A. The regenerative potential of the kidney: What can we learn from developmental biology? Stem Cell Rev. Pep 2010, 6, 650–657. [Google Scholar]
- Oxburgh, L.; Brown, A.C.; Fetting, J.; Hill, B. BMP signaling in the nephron progenitor niche. Pediatr. Nephrol 2011, 26, 1491–1497. [Google Scholar]
- Piludu, M.; Fanos, V.; Congiu, T.; Piras, M.; Gerosa, C.; Mocci, C.; Fanni, D.; Nemolato, S.; Iacovidou, N.; Faa, G. The pine-cone body: An intermediate structure between the cap mesenchyme and the renal vesicle in the developing nod mouse kidney revealed by an ultrastructural study. J. Matern. Fetal Neonatal. Med 2012, 25, 72–75. [Google Scholar]
- Kloth, S.; Ebenbeck, C.; Monzer, J.; de Vries, U.; Minuth, W.W. Three-dimensional organization of the developing vasculature of the kidney. Cell Tissue Res 1997, 287, 193–201. [Google Scholar]
- Brizzi, M.F.; Tarone, G.; Defilippi, P. Extracellular matrix, integrins, and growth factors as tailors of the stem cell niche. Curr. Opin. Cell Biol 2012, 24, 645–651. [Google Scholar]
- Minuth, W.W.; Denk, L. Illustration of extensive extracellular matrix at the epithelial-mesenchymal interface within the renal stem/progenitor cell niche. BMC Clin. Pathol 2012, 25, 16. [Google Scholar]
- Minuth, W.W.; Denk, L. Cell projections and extracellular matrix cross the interstitial interface within the renal stem/progenitor cell niche. Nephron. Expt. Nephrol 2013, 122, 131–140. [Google Scholar]
- Asko, J.A.; Richardson, G.P. The ultrastructural organization and properties of the mouse tectorial membrane matrix. Hear Res 1988, 35, 21–38. [Google Scholar]
- Rothenburger, M.; Völker, W.; Vischer, P.; Glasmacher, B.; Scheid, H.H.; Deiwick, M. Ultrastructure of proteoglycans in tissue-engineered cardiovascular structures. Tissue Eng 2002, 8, 1049–1056. [Google Scholar]
- Minuth, W.W.; Denk, L.; Meese, C.; Rachel, R.; Roessger, A. Ultrastructural insights in the interface between generated renal tubules and a polyester interstitium. Langmuir 2009, 25, 4621–4627. [Google Scholar]
- Liu, L.; Dunn, S.T.; Christakos, S.; Hanson-Painton, O.; Bourdeau, J.E. Calbindin-D28k gene expression in the developing mouse kidney. Kidney Int 1993, 44, 322–330. [Google Scholar]
- Nishinakamura, R. Stem cells in the embryonic kidney. Kidney Int 2008, 73, 913–917. [Google Scholar]






© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Minuth, W.W.; Denk, L. The Interstitial Interface within the Renal Stem/Progenitor Cell Niche Exhibits an Unique Microheterogeneous Composition. Int. J. Mol. Sci. 2013, 14, 13657-13669. https://doi.org/10.3390/ijms140713657
Minuth WW, Denk L. The Interstitial Interface within the Renal Stem/Progenitor Cell Niche Exhibits an Unique Microheterogeneous Composition. International Journal of Molecular Sciences. 2013; 14(7):13657-13669. https://doi.org/10.3390/ijms140713657
Chicago/Turabian StyleMinuth, Will W., and Lucia Denk. 2013. "The Interstitial Interface within the Renal Stem/Progenitor Cell Niche Exhibits an Unique Microheterogeneous Composition" International Journal of Molecular Sciences 14, no. 7: 13657-13669. https://doi.org/10.3390/ijms140713657
APA StyleMinuth, W. W., & Denk, L. (2013). The Interstitial Interface within the Renal Stem/Progenitor Cell Niche Exhibits an Unique Microheterogeneous Composition. International Journal of Molecular Sciences, 14(7), 13657-13669. https://doi.org/10.3390/ijms140713657