1. Introduction
Osteosarcoma is the most common malignant primary bone tumor, particularly among children and adolescent [
1,
2]. Osteosarcoma has high local aggressiveness with a metastasis tendency most commonly to the lung, pleura, bone, and heart [
3]. Consequently, long-term survival is dismal as most osteosarcoma patients often die of systemic metastasis, especially diffuse lung metastases [
4,
5]. Despite recent advances in aggressive treatment modalities, such as adjuvant chemotherapy and wide tumor resection, the five-year survival rate of osteosarcoma patients is only 55%–60%, and lung and skeletal metastases occurs in approximately 30% to 40% of these patients [
6]. Meanwhile, high-dose chemotherapy has a number of adverse effects such as gastrointestinal reactions, bone marrow suppression, cardiac toxicity, and renal toxicity [
7]. Thus, it is urgent to develop new treatments for osteosarcoma.
Growing evidence has shown that voltage-gated potassium channels (Kv channels) play an important role in the development and growth of cancers [
8–
10]. Among Kv channels, potassium voltage-gated channel, shaker-related subfamily, member 3, also known as KCNA3 or Kv1.3, is a protein encoded by the KCNA3 gene, which appears to be intronless and is clustered together with KCNA2 and KCNA10 genes on chromosome 1 [
11]. Kv1.3 regulates membrane potential and Ca
2+ signaling in human T cells and thus plays an important role in T cell proliferation and activation [
12]. In addition, Kv1.3 is known to be expressed in the inner mitochondrial membrane of lymphocytes, and apoptotic protein Bax could insert into the outer mitochondrial membrane and occlude the pore of Kv1.3 via a lysine residue [
13]. Therefore, Kv1.3 modulation appears as one key mechanism that contributes to apoptosis [
14,
15]. Interestingly, recent studies have shown that Kv1.3 is implicated in the growth and proliferation of various cancer cells, such as breast, gastric, gliomas, and prostate cancer [
16–
19]. The tight relationship between Kv1.3 expression and cancer indicates that Kv1.3 is a molecular target for the development of anticancer therapies [
20]. Nevertheless, there is currently debate regarding the relationship between the aberrant Kv1.3 expression and malignancy and invasiveness of cancers. For example, there are apparently controversial conclusions regarding the correlation between Kv1.3 level and tumorigenicity in breast cancer, gliomas, and prostate cancer [
16,
18,
19]. Therefore, further investigations are needed to elucidate the function of Kv1.3 in the tumorigenesis of different cancers.
Up to now, little is known about the relationship between the expression status of Kv1.3 and human osteosarcoma. Moreover, the biological functions of Kv1.3, during the multiple step processes of tumorigenesis of osteosarcoma, are largely unclear. To address these questions, we examined the expression of Kv1.3 in human osteosarcoma by RT-PCR, Western-blot and immunohistochemistry, and determined cell proliferation and apoptosis following knockdown of Kv1.3 by RNA interference (RNAi) in cultured osteosarcoma MG-63 cells. We further monitored tumor growth of osteosarcoma MG-63 mouse xenografts after downregulation of Kv1.3 by adenovirus delivered shRNA.
3. Discussion
Kv channels subtype Kv1.3 has been implicated in the regulation of many cellular functions, including membrane potential, solute and water transport, cell-volume, adhesion, motility, apoptosis and proliferation [
22]. Numerous studies have demonstrated that aberrant expression of Kv1.3 is involved in the progression and survival of cancers [
10]. However, its function during tumorigenesis is debatable [
16,
23,
24]. Up to now, the expression and function of Kv1.3 in human osteosarcoma remain unknown. Therefore, we investigated the expression and function of Kv1.3 in human osteosarcoma in this study.
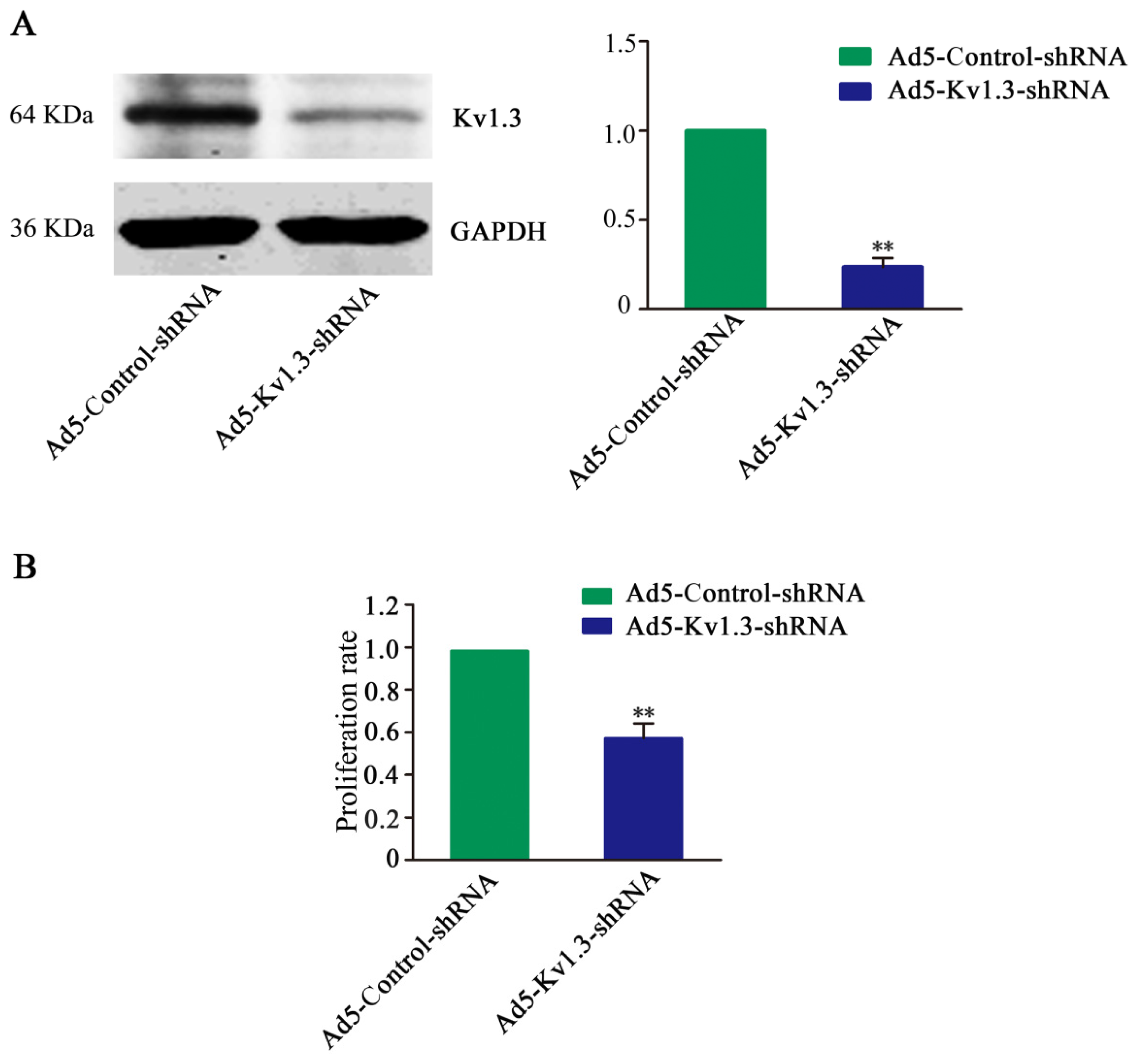
By RT-PCR, Western blot, and immunohistochemistry, we found increased expression of Kv1.3 in human osteosarcoma cell line and tissues. Compared with pharmacologic Kv1.3 inhibitors, such as 4-aminopyridine (4-AP) [
25], tetraethylammonium (TEA) [
25], and margatoxin (MgTX) [
26], small interfering RNA (siRNA) is a more specific tool to investigate the role of Kv1.3 in cancer progression, as siRNA mediated knockdown of Kv1.3 resulted in reduced proliferation of tumor cell lines with less nonspecific responses [
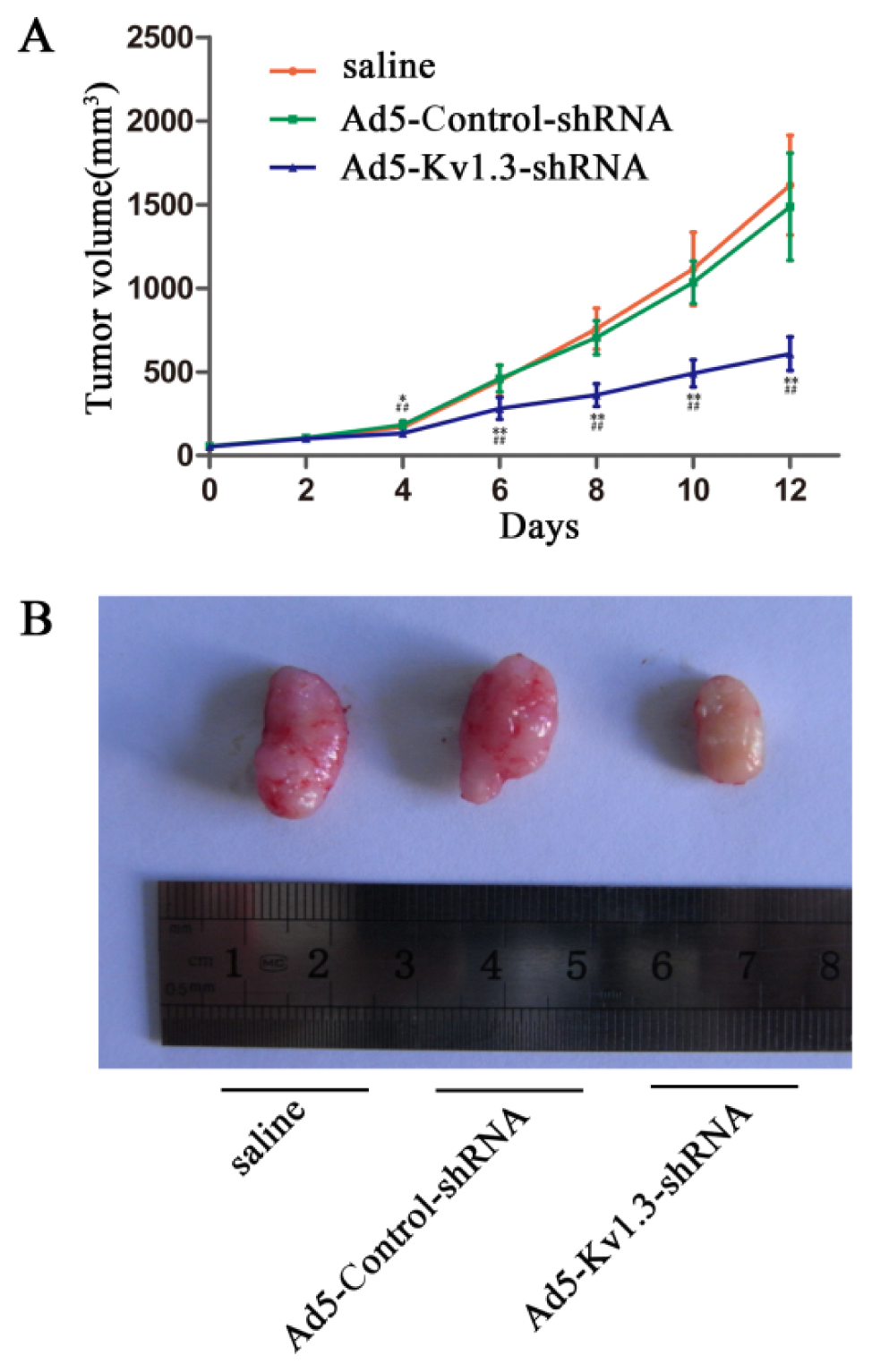
27]. In our study, Kv1.3-shRNA effectively downregulated Kv1.3 expression and significantly inhibited the growth of osterosarcoma cells
in vitro and
in vivo.
Apoptotic cell death is one of the main mechanisms involved in the decrease of cell growth following the inactivation of oncogene or the activation of tumor suppressor gene. To determine whether the growth inhibitory effect of MG-63 cells mediated by Kv1.3-shRNA was related to the induction of apoptosis, we used flow cytometry to examine apoptotic rate in Ad5-Kv1.3-shRNA infected MG-63 cells and found that Kv1.3-shRNA induced a significant increase in apoptosis. The activation of caspases is one of the major processes in apoptosis [
28]. Thus, we examined the levels of caspase-3/7, the key apoptotic caspases, and the cleavage of PARP, a marker of apoptosis, in Ad5-Kv1.3-shRNA infected cells. Significant increases of activated caspase-3/7, cleaved PARP and cleaved caspase-3 were observed in Ad5-Kv1.3-shRNA infected cells. These data suggest that inhibition of Kv1.3 leads to the activation of caspase-3/7 and the execution of apoptosis.
Notably, recent data suggest that Kv1.3 is active in the inner mitochondrial membrane. On the induction of apoptosis, Bax on the outer mitochondrial membrane binds to and inhibits Kv1.3, resulting in hyperpolarization, an increase in reactive oxygen species production, and cytochrome c release [
13]. Thus, the inhibition of Kv1.3 activity or expression has been proposed as a potential mechanism that could induce apoptosis [
14,
15]. Consistent with these previous data, in this study we found that knockdown of Kv1.3 induced the apoptosis of osteosarcoma cells.
In conclusion, this is the first study to show that Kv1.3 was upregulated in human osteosarcoma and downregulation of Kv1.3 suppressed osteosarcoma growth in vivo and osteosarcoma cell proliferation in vitro, accompanied by increased apoptosis. Our findings suggest that Kv1.3 is a novel target for the development of molecular targeted cancer therapies for osterosarcoma.
4. Experimental Section
4.1. Cell Culture
Human osteosarcoma cell line MG-63, human embryonic kidney cell line 293 (HEK293), human Jurkat cell line TIB-152, and human osteoblastic cell line hFOB 1.19 were obtained from the American type culture of collection (ATCC). MG-63, HEK293, and TIB-152 cells were cultured in RPMI-1640 medium (Gibco, Rockville, MD, USA) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37 °C in a humidified atmosphere with 5% CO2. hFOB 1.19 cells were cultured in ham’sF12/ Dulbecco’s modified Eagle medium (DMEM, Gibco, Rockville, MD, USA) without phenol red supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin at 34 °C in a humidified atmosphere with 5% CO2. Four cell lines were subcultured every three to four days.
4.2. Preparation of Adenoviral shRNA Vectors
The oligonucleotide targeting human Eag was designed and selected as the template: GCA GUG GUA ACC AUG ACA A, which shared no homology with other coding sequences in human genome by BLAST analysis. A ring sequence of nine base pairs (TTC AAG ACG) existed between the sense and antisense strands. The shRNA was synthesized by Sangon Biotech (Shanghai, China). Plasmid pGeneSil-1 containing the human U6 promoter was purchased from GeneSil Biotechnology (Wuhan, China). The shRNA-expressing cassette was subcloned into pAdTrack vector between HindIII and XbaI sites. The recombinant plasmid was linearized by digestion with restriction endonuclease, and subsequently cotransformed into E. coli BJ5183 cells with an adenoviral backbone plasmid, pAdEasy-1. Recombinant plasmids were selected for kanamycin resistance, and transduced into HEK293 cells. A recombinant adenovirus expressing shRNA against Kv1.3 (Ad5-Kv1.3-shRNA) was generated. The recombinant adenovirus (Ad5-Control-shRNA), which contained the CTA CCT GTT CTA GTC TGG ACT sequence and did not target any known human genes, was generated as the control for Ad5-Kv1.3-shRNA. All viruses were propagated and purified on a CsCl gradient using standard methods. The viruses were titered for viral particles using standard methods based on spectrophotometry at 260 nm. Functional titer (plaque forming units) was determined with a plaque assay on HEK293 cells according to the method developed by Quantum Biotechnology.
4.3. Adenovirus Infection
MG-63 cells (1 × 105) in serum-free RPMI-1640 were infected with Ad5-Kv1.3-shRNA or Ad5-Control-shRNA at 5 MOI (multiplicity of infection, calculated as PFU/cell numbers) in a humidified atmosphere of 5% CO2 at 37 °C. Virus-containing medium was removed eight hours later and replaced with fresh RPMI-1640 medium containing 10% (v/v) FBS. Cells were incubated for additional 48 h.
The total RNA was isolated from the cultured cells by Trizol reagent (Invitrogen, Rockville, MD, USA). RNA purity and integrity was checked by denaturing 1% (w/v) agarose gel. cDNA was synthesized from 1 μg of total RNA using 200 U reverse transcriptase (Takara, Tokyo, Japan) in a 20 μL reaction volume containing 200 μM dNTPs and 2.5 μM oligo-dT primer at 30 °C for 10 min, then at 42 °C for 60 min, and finally at 80 °C for 5 min. One μL of cDNA were amplified by PCR for 35 cycles in 25 μL reaction volume containing 2.5 U DNA polymerase and 200 μM dNTPs. Sequences of forward and reverse primers, amplified fragment sizes, and annealing temperatures were as follows: Kv1.3, 5′-GGT CAT CAA CAT CTCCGG CGT GCG CT-3′, 5′-AGG GCC GCT CCT CCT CCCGC-3′, 67 °C, 314 bp. β-actin, 5′-TCC ACC TTC CAG CAG ATG TG-3′, 5′GCA TTT GCG GTG GAC GAT-3′, 54 °C, 75 bp. PCR products were run on a 2% agarose gel (Sigma, St. Louis, MO, USA) with a 100 bp DNA ladder, and the bands were visualized by ethidium bromide staining on a UV trans illuminator. Each experiment was repeated three times. Some of PCR products were sequenced to check PCR specificity. The relative mRNA level of Kv1.3 was calculated by densitometry analysis of the gels with that of β-actin as the standard.
4.4. Western Blot Analysis
About 5–6 × 107 cells were collected and lysed in ice-cold lysis buffer containing 50 mmol/L Tris-Cl (pH 7.5), 150 mmol/L NaCl, 0.2 mmol/L EDTA, 1 mmol/L PMSF and 1% (v/v) Nonidet-P40 for 30 min. The lysates were centrifuged at 13,200 rpm for 10 min at 4 °C and the supernatants were collected. 25 μg protein were resolved by a 12% SDS-PAGE and blotted onto nitrocellulose membranes (Bio-Rad, Richmond, CA, USA). Membranes were blocked with 10% (w/v) nonfat milk at room temperature for one hour, and then incubated with antibodies against Kv1.3 (Abcam, Cambridge, MA, USA), PARP (Cell Signaling Technology®, Danvers, MA, USA), caspase-3, cleaved caspase-3 (Cell Signaling) and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Abcam) overnight at 4 °C, followed by incubation with horseradish peroxidase-conjugated goat anti-rabbit or anti-mouse secondary antibody (Santa Cruz Biotechnology, CA, USA) at room temperature for one hour. The membranes were, then, developed with chemiluminescent detection kit (Zhongshan Biotechnology, Beijing, China) and exposed to X-ray films. Experiments were performed at least three times with representative data presented.
4.5. Sample Collection and Immunohistochemistry
A total of 41 formalin-fixed, paraffin-embedded osteosarcoma specimens from 22 patients (before the administration of neoadjuvant chemotherapy) were acquired from the Affiliated Southeast Hospital of Xiamen University between January 2009 and June 2012. As a positive control sample [
18], confirmed healthy human brain, obtained by biopsy, was used. All the specimens were collected after obtaining written informed consent according to a protocol approved by Institutional Review Board of the Affiliated Southeast Hospital of Xiamen University.
For immunohistochemistry, the tissue slide was baked on a rack in a dry oven at 60 °C for two hours to remove the coated paraffin. The slide was immersed twice in xylene (Zhongshan Biotechnology) for three minutes, hydrated consectively with 100%, 95%, 70%, and 50% ethanol (Zhongshan Biotechnology) and rinsed with cold top water for five minutes. After dewaxing and blocking endogenous peroxidases, the sections were treated at 100 °C in EDTA (1 mM, pH 8.0) for antigen retrieval, and then incubated with anti-Kv1.3 antibody (1:500, Abcam) overnight at 4 °C. The slide was washed with PBS the following day and incubated with biotinylated goat anti-rabbit IgG (Abcam) at room temperature for one hour. Avidin biotin complex (ABC) (Zhongshan Biotechnology) and diaminobenzamidine (DAB) (Zhongshan Biotechnology) were used to visualize the tissue slide according to manufacturer’s instructions, and then the sections were counterstained with haematoxylin.
4.6. Cell Proliferation Assay
The cell proliferation was analyzed by using Cell Counting Assay Kit-8 (CCK-8) (Dojindo Molecular Technologies, Gaithersburg, MD, USA) according to the manufacturer’s protocol. In brief, 1 × 105 cells were starved in serum-free medium for 12 h and then the cells were transduced. After 48 h, cells were harvested. Ten microliters of Cell Counting Assay Kit-8 solution was added to each well, the cells were incubated for another hour, and the absorbance (A) at 450 nm was measured by using spectrophotometer (Bio-Rad). Experiments were performed at least three times with representative data presented.
4.7. Tumor Xenograft Model
Thymys-null BALB/c nude mice (female, age six to eight weeks) were obtained from the Animal Center of Chinese Academy of Medical Sciences. All animal procedures were performed according to approved protocols and in accordance with recommendations for the proper use and care of laboratory animals. Osteosarcoma xenografts were established in nude mice as previously described [
24]. A total of 1.5 × 10
6 MG-63 cells in 150 μL phosphate-buffered aline (PBS) were subcutaneously injected in the hind right leg. One week later, the tumors grew to visible size. The osteosarcoma-bearing mice were randomly divided into three groups (six in each group). Group 1 received intra-tumor injections with Ad5- Kv1.3-shRNA (10 MOI) every two days (six injections total). Group 2 received intra-tumor injections of Ad5-Control-shRNA (10 MOI) every two days (six injections total). Group 3 received normal saline injection as controls. All treatments were given every other day for a total of seven doses. Tumor volume (cm
3) was determined based on the following formula: ab
2/2 where a was the length and b was the width of the tumor [
29].
4.8. Analysis of Cell Apoptosis
Apoptosis was determined using flpo cytometry and the Apo-ONE homogeneous caspase-3/7 assay (Promega, Madison, WI, USA) according to the manufacturer’s instructions.
For flor cytometry, cells were prepared and analyzed using a FACScan apparatus (Becton-Dickinson, San Jose, CA, USA) following the manufacturer’s protocol. Early apoptotic cells were defined as Annexin V positive and propidium iodide (PI) negative cells.
For caspase-3/7 assay, 20 μL of cell suspension (1 × 105 cells/mL) was seeded into 96-well plates (Sigma, St. Louis, MO, USA) and incubated at 37 °C overnight. After the cells were treated with various adenovirus constructs, an equal volume of Apo-ONE Caspase-3/7-reagent was added to a 96-well plate and incubated at room temperature for one hour. The luminescence of each sample was measured in a plate-reading luminometer (Tecan, Switzerland). Experiments were performed at least three times with representative data presented.
4.9. Statistical Analysis
All data were presented as mean ± standard deviation (SD). Statistical significance was determined using t-test or analysis of variance (ANOVA) using the SPSS18.0 program. p < 0.05 was considered as statistically significant.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




