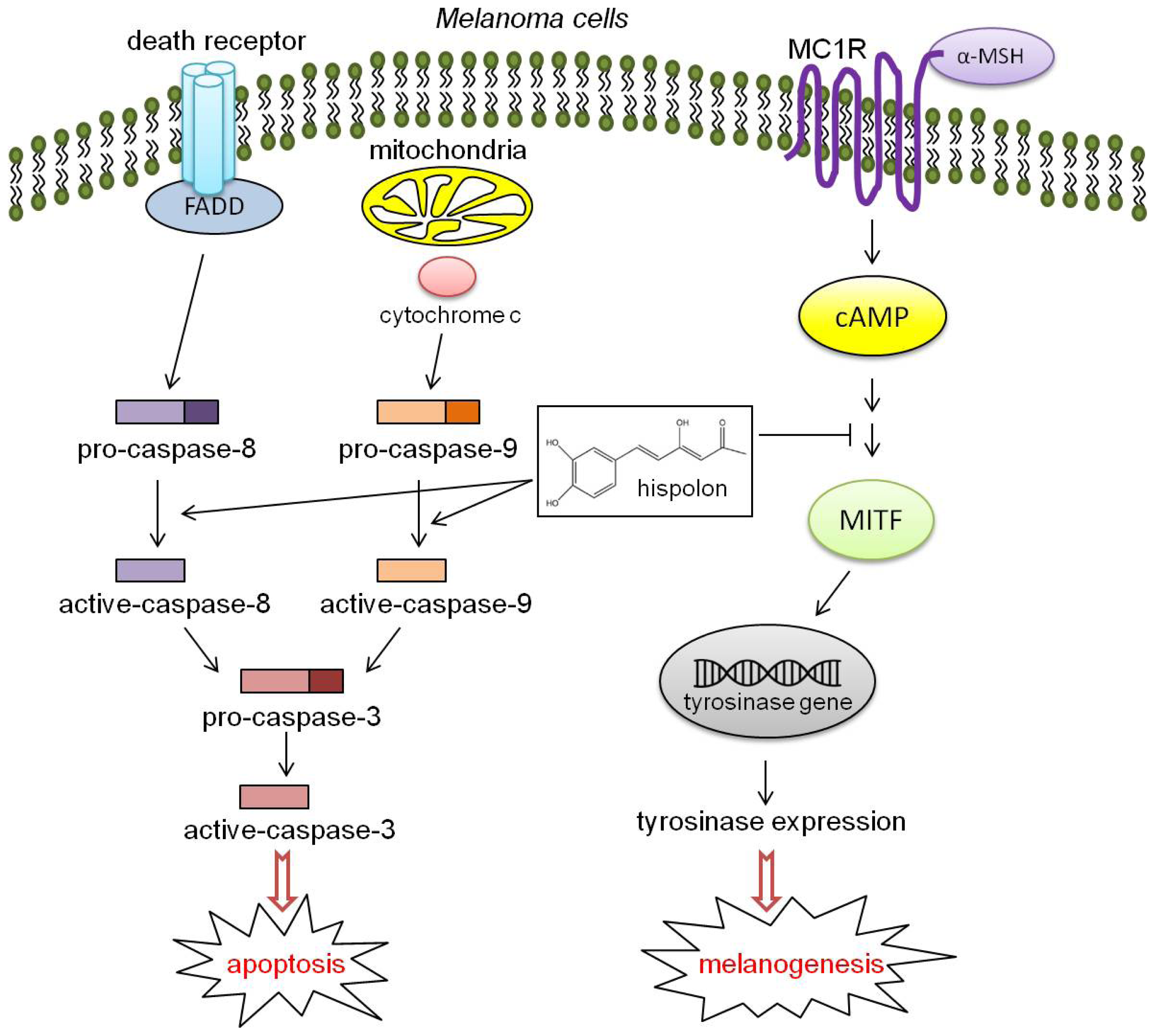
Hispolon Decreases Melanin Production and Induces Apoptosis in Melanoma Cells through the Downregulation of Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Expressions and the Activation of Caspase-3, -8 and -9



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
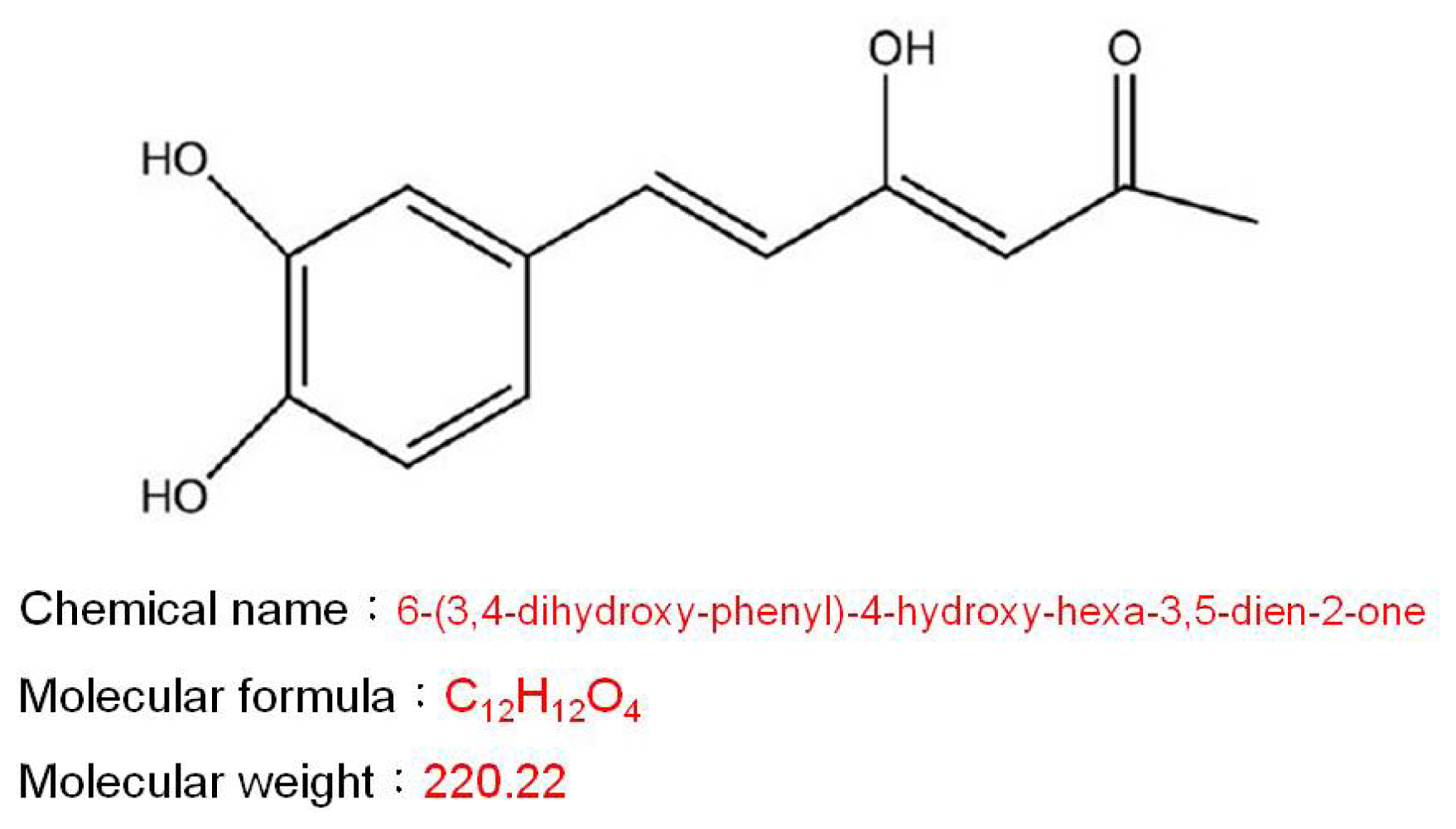
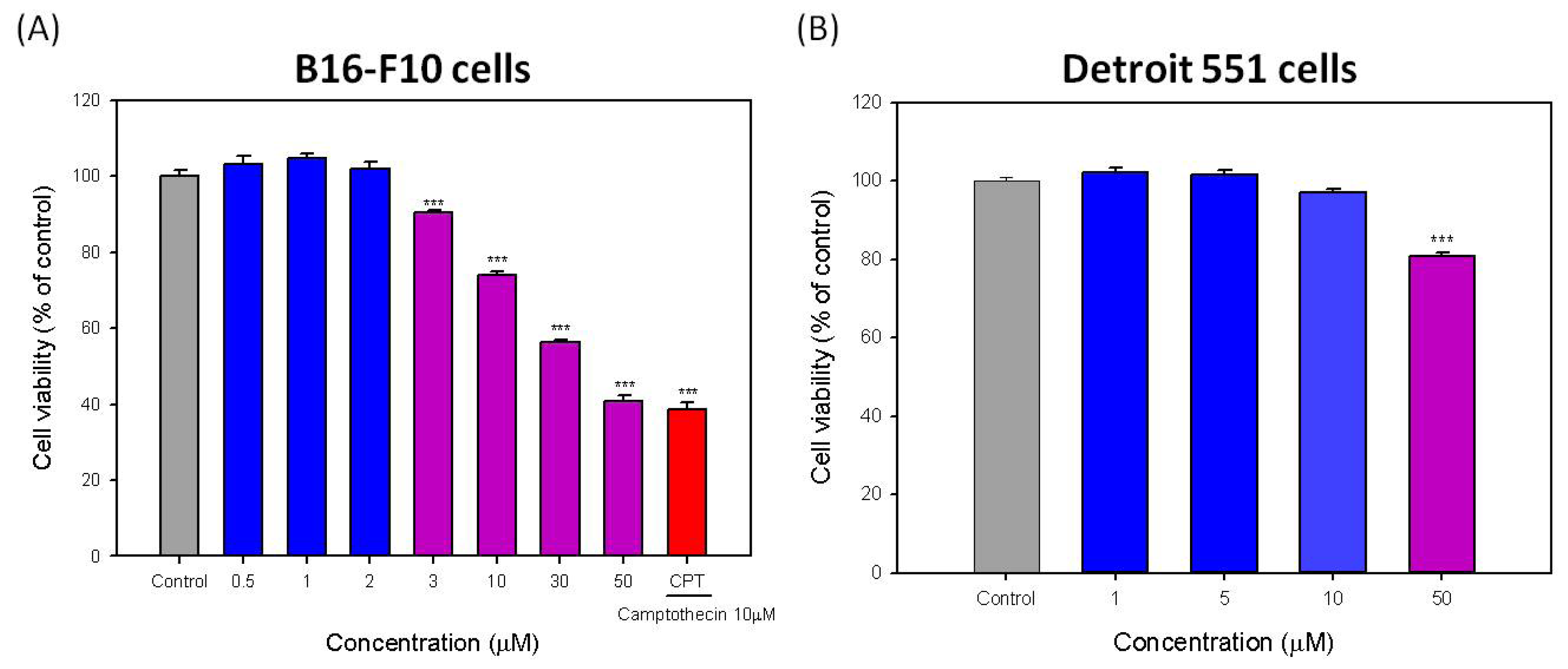
2.1. Effects of Hispolon on Cell Viability
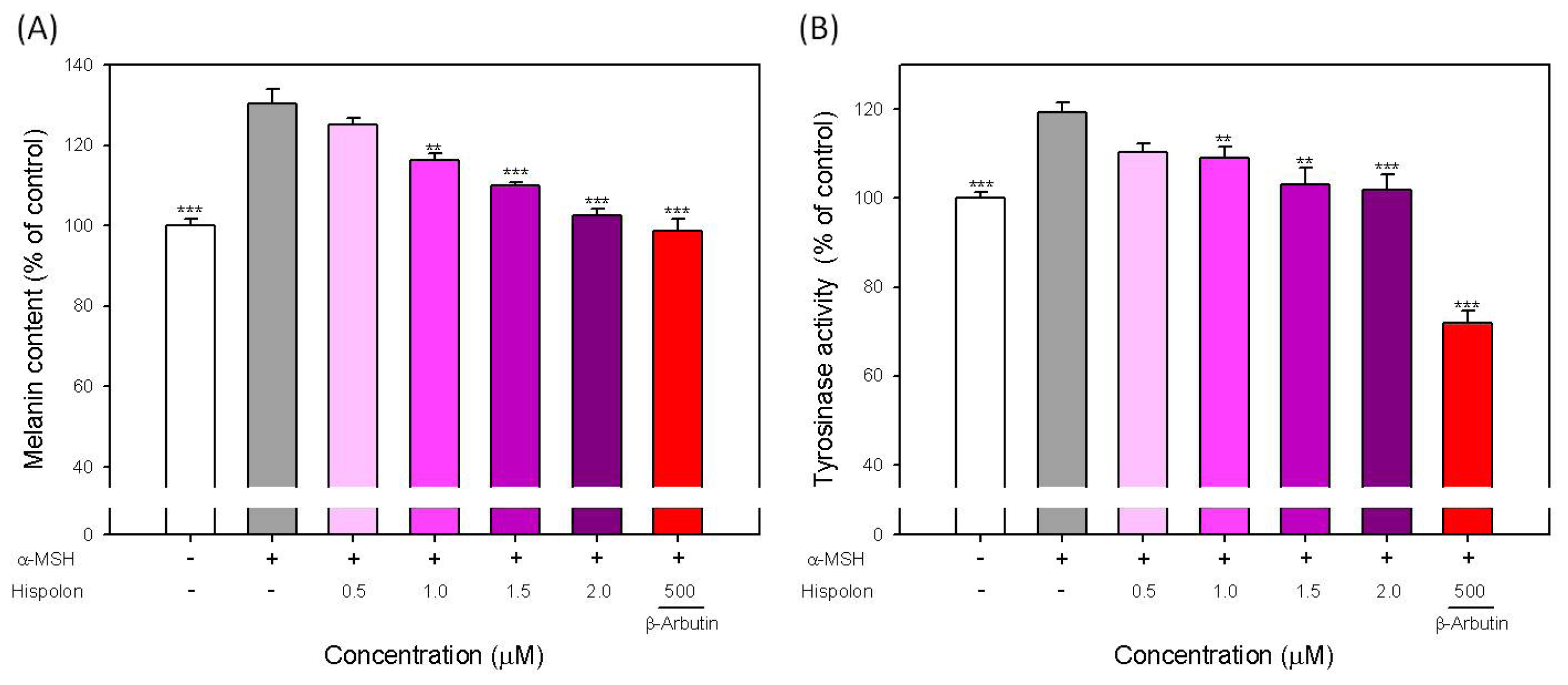
2.2. Effects of Hispolon on the Melanin Content and Cellular Tyrosinase Activity in α-MSH-Stimulated B16-F10 Cells
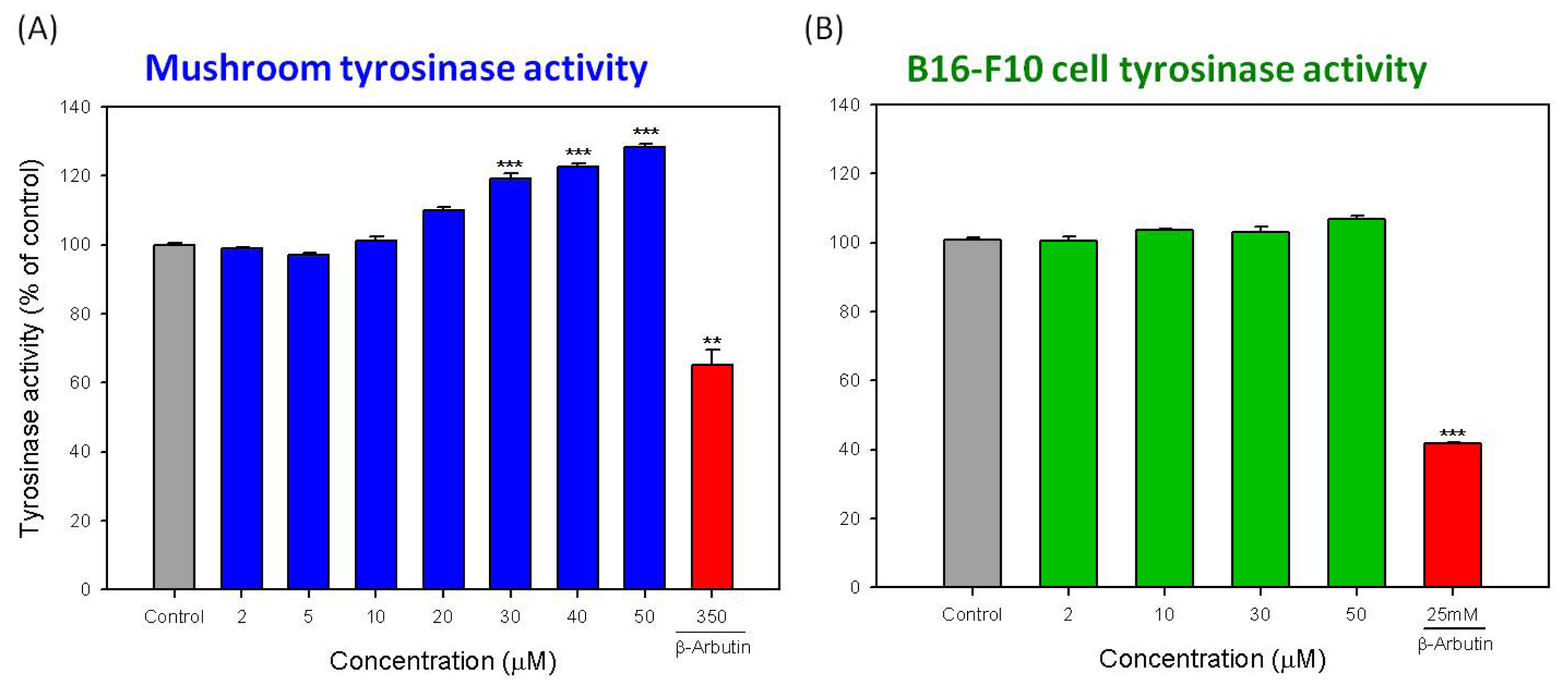
2.3. Direct Effects of Hispolon on the Activity of Mushroom Tyrosinase and Cellular Tyrosinase
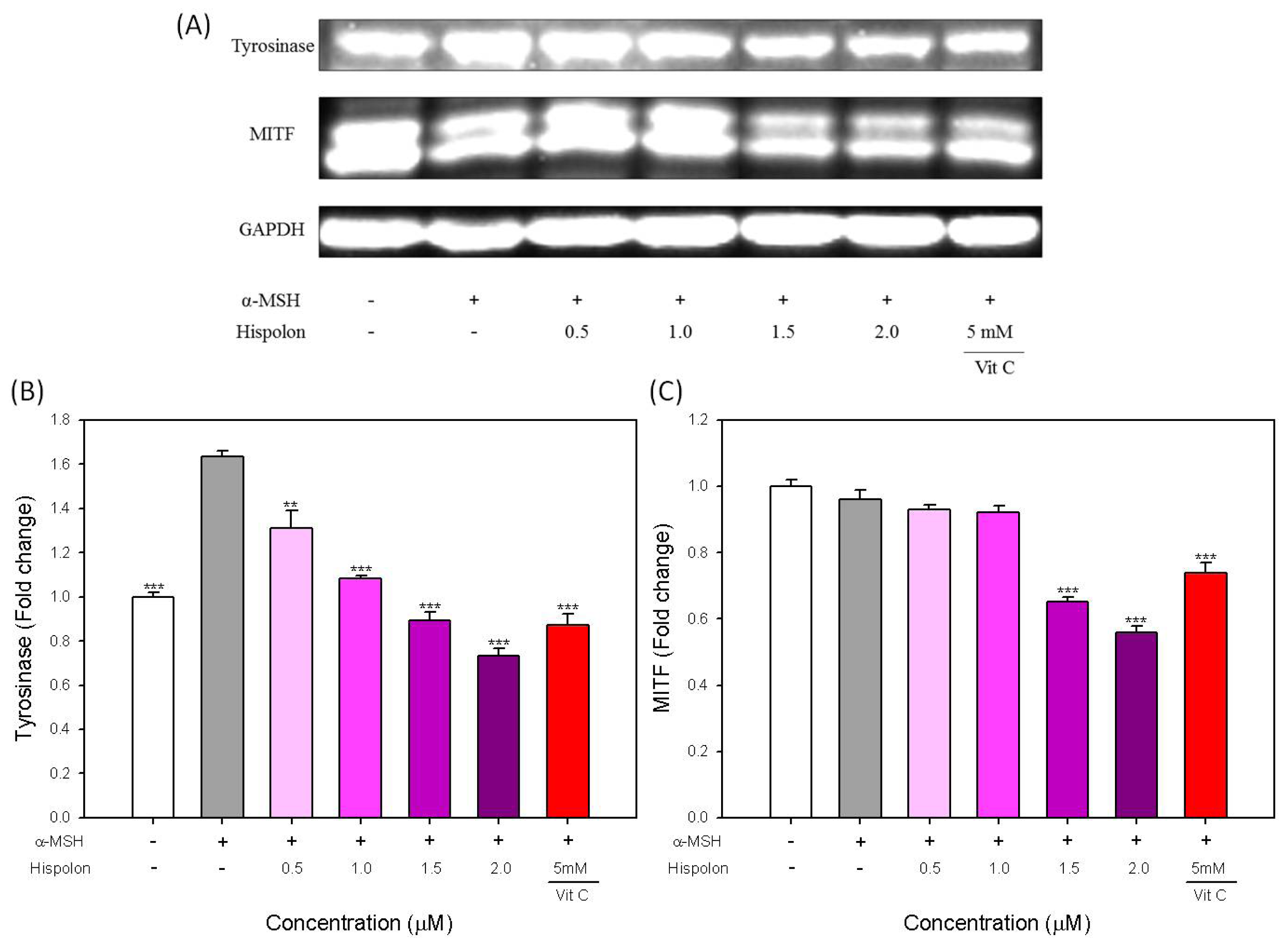
2.4. Effects of Hispolon on the Protein Level of Tyrosinase and MITF in α-MSH-Stimulated B16-F10 Cells
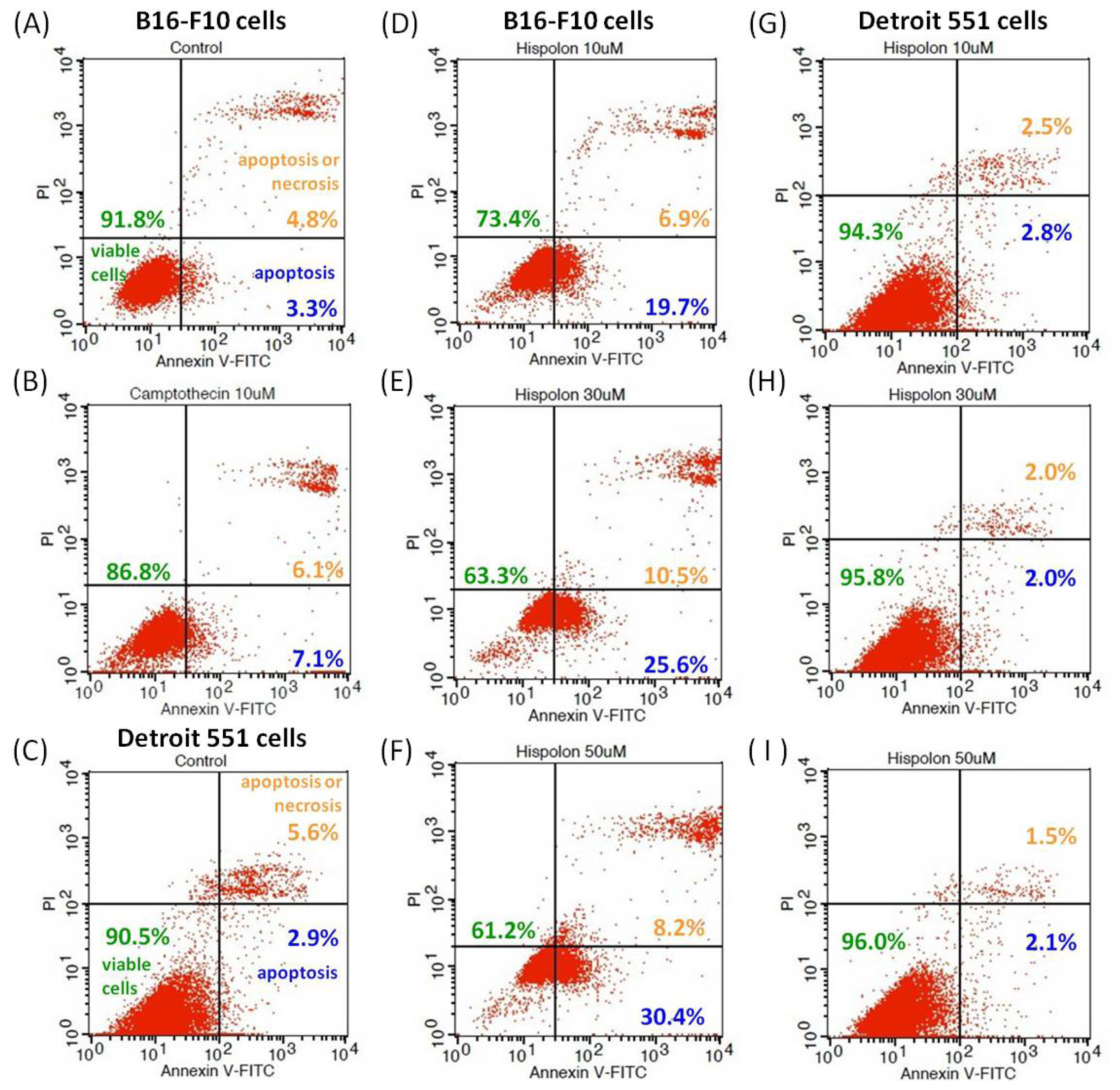
2.5. Effects of Hispolon on Apoptosis in B16-F10 and Detroit 551 Cells
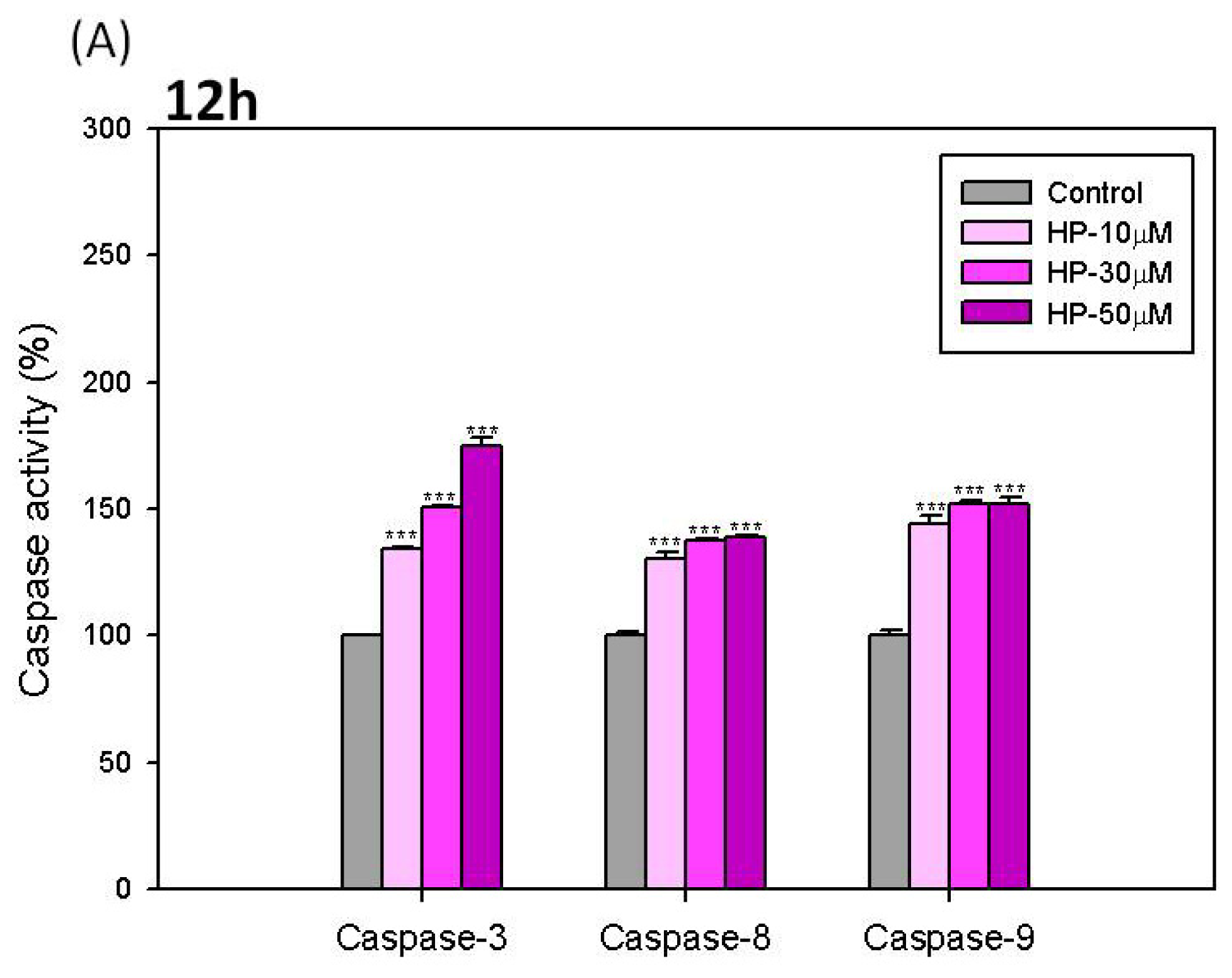
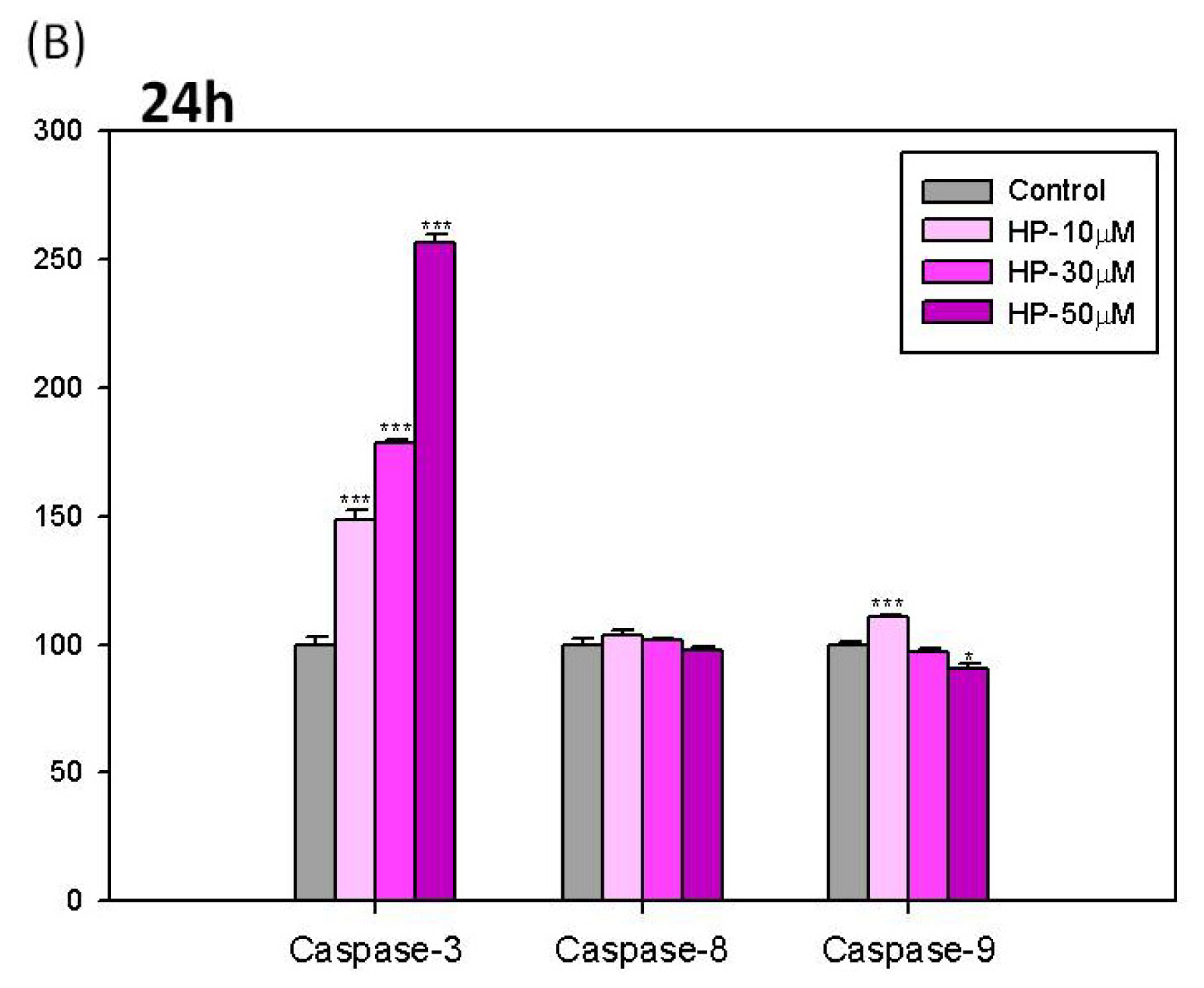
2.6. Effects of Hispolon on the Activation of Caspase-3, -8 and -9 in B16-F10 Cells
3. Experimental Section
3.1. Materials
3.2. Cell Line and Cell Culture
3.3. MTT Assay for Cell Viability
3.4. Melanin Content Analysis
3.5. Tyrosinase Activity Assays
3.6. Western Blot Analysis
3.7. Annexin-V/PI Staining Assay
3.8. Caspases Activity Assays
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Maddodi, N.; Jayanthy, A.; Setaluri, V. Shining light on skin pigmentation: The darker and the brighter side of effects of UV radiation. Photochem. Photobiol 2012, 88, 1075–1082. [Google Scholar]
- Gillbro, J.M.; Olsson, M.J. The melanogenesis and mechanisms of skin-lightening agents-existing and new approaches. Int. J. Cosmet. Sci 2011, 33, 210–221. [Google Scholar]
- Jeong, Y.M.; Oh, W.K.; Tran, T.L.; Kim, W.K.; Sung, S.H.; Bae, K.; Lee, S.; Sung, J.H. Aglycone of Rh4 inhibits melanin synthesis in B16 melanoma cells: Possible involvement of the protein kinase A pathway. Biosci. Biotechnol. Biochem 2013, 77, 119–125. [Google Scholar]
- Su, T.R.; Lin, J.J.; Tsai, C.C.; Huang, T.K.; Yang, Z.Y.; Wu, M.O.; Zheng, Y.Q.; Su, C.C.; Wu, Y.J. Inhibition of melanogenesis by gallic acid: Possible involvement of the PI3K/Akt, MEK/ERK and Wnt/β-catenin signaling pathways in B16F10 cells. Int. J. Mol. Sci 2013, 14, 20443–20458. [Google Scholar]
- Chou, S.T.; Chang, W.L.; Chang, C.T.; Hsu, S.L.; Lin, Y.C.; Shih, Y. Cinnamomum cassia essential oil inhibits α-MSH-induced melanin production and oxidative stress in murine B16 melanoma cells. Int. J. Mol. Sci 2013, 14, 19186–19201. [Google Scholar]
- Yang, C.H.; Chang, N.F.; Chen, Y.S.; Lee, S.M.; Lin, P.J.; Lin, C.C. Comparative study on the photostability of arbutin and deoxy arbutin: Sensitivity to ultraviolet radiation and enhanced photostability by the water-soluble sunscreen, benzophenone-4. Biosci. Biotechnol. Biochem 2013, 77, 1127–1130. [Google Scholar]
- Dellinger, R.W.; Matundan, H.H.; Ahmed, A.S.; Duong, P.H.; Meyskens, F.L., Jr. Anti-cancer drugs elicit re-expression of UDP-glucuronosyltransferases in melanoma cells. PLoS One 2012, 7, e47696. [Google Scholar]
- Gray-Schopfer, V.; Wellbrock, C.; Marais, R. Melanoma biology and new targeted therapy. Nature 2007, 445, 851–857. [Google Scholar]
- Looi, C.Y.; Moharram, B.; Paydar, M.; Wong, Y.L.; Leong, K.H.; Mohamad, K.; Arya, A.; Wong, W.F.; Mustafa, M.R. Induction of apoptosis in melanoma A375 cells by a chloroform fraction of Centratherum anthelminticum (L.) seeds involves NF-κB, p53 and Bcl-2-controlled mitochondrial signaling pathways. BMC Complement. Altern. Med 2013, 13, 166. [Google Scholar]
- Lim, B.O.; Yamada, K.; Cho, B.G.; Jeon, T.; Hwang, S.G.; Park, T.; Kang, S.A.; Park, D.K. Comparative study on the modulation of IgE and cytokine production by Phellinus linteus grown on germinated brown Rice, Phellinus Linteus and germinated brown rice in murine splenocytes. Biosci. Biotechnol. Biochem 2004, 68, 2391–2394. [Google Scholar]
- Kim, G.Y.; Lee, J.Y.; Lee, J.O.; Ryu, C.H.; Choi, B.T.; Jeong, Y.K.; Lee, K.W.; Jeong, S.C.; Choi, Y.H. Partial characterization and immunostimulatory effect of a novel polysaccharide-protein complex extracted from Phellinus linteus. Biosci. Biotechnol. Biochem 2006, 70, 1218–1226. [Google Scholar]
- Kim, G.Y.; Oh, W.K.; Shin, B.C.; Shin, Y.I.; Park, Y.C.; Ahn, S.C.; Lee, J.D.; Bae, Y.S.; Kwak, J.Y.; Park, Y.M. Proteoglycan isolated from Phellinus linteus inhibits tumor growth through mechanisms leading to an activation of CD11c+CD8+ DC and type I helper T cell-dominant immune state. FEBS Lett 2004, 576, 391–400. [Google Scholar]
- Zhu, T.; Kim, S.H.; Chen, C.Y. A medicinal mushroom Phellinus linteus. Curr. Med. Chem 2008, 15, 1330–1335. [Google Scholar]
- Ali, N.A.; Ludtke, J.; Pilgrim, H.; Lindequist, U. Inhibition of chemiluminescence response of human mononuclear cells and suppression of mitogen-induced proliferation of spleen lymphocytes of mice by hispolon and hispidin. Pharmazie 1996, 51, 667–670. [Google Scholar]
- Huang, G.J.; Deng, J.S.; Huang, S.S.; Hu, M.L. Hispolon induces apoptosis and cell cycle arrest of human hepatocellular carcinoma Hep3B cells by modulating ERK phosphorylation. J. Agric. Food Chem 2011, 59, 7104–7113. [Google Scholar]
- Huang, G.J.; Deng, J.S.; Chiu, C.S.; Liao, J.C.; Hsieh, W.T.; Sheu, M.J.; Wu, C.H. Hispolon protects against acute liver damage in the rat by inhibiting lipid peroxidation, proinflammatory cytokine, and oxidative stress and downregulating the expressions of iNOS, COX-2, and MMP-9. Evid. Based Complement. Alternat. Med 2011, 2012, 480714. [Google Scholar]
- Chen, W.; Zhao, Z.; Li, L.; Wu, B.; Chen, S.F.; Zhou, H.; Wang, Y.; Li, Y.Q. Hispolon induces apoptosis in human gastric cancer cells through a ROS-mediated mitochondrial pathway. Free Radic. Biol. Med 2008, 45, 60–72. [Google Scholar]
- Hsiao, P.C.; Hsieh, Y.H.; Chow, J.M.; Yang, S.F.; Hsiao, M.; Hua, K.T.; Lin, C.H.; Chen, H.Y.; Chien, M.H. Hispolon induces apoptosis through JNK1/2-mediated activation of a Caspase-8, -9, and -3-dependent pathway in Acute Myeloid Leukemia (AML) cells and inhibits AML Xenograft tumor growth in vivo. J. Agric. Food Chem. 2013, 61, 10063–10073. [Google Scholar]
- Huang, G.J.; Yang, C.M.; Chang, Y.S.; Amagaya, S.; Wang, H.C.; Hou, W.C.; Huang, S.S.; Hu, M.L. Hispolon suppresses SK-Hep1 human hepatoma cell metastasis by inhibiting matrix metalloproteinase-2/9 and urokinase-plasminogen activator through the PI3K/Akt and ERK signaling pathways. J. Agric. Food Chem 2010, 58, 9468–9475. [Google Scholar]
- Zu, Y.; Wang, D.; Zhao, X.; Jiang, R.; Zhang, Q.; Zhao, D.; Li, Y.; Zu, B.; Sun, Z. A novel preparation method for camptothecin (CPT) loaded folic acid conjugated dextran tumor-targeted nanoparticles. Int. J. Mol. Sci 2011, 12, 4237–4249. [Google Scholar]
- Chen, Y.S.; Lee, S.M.; Lin, C.C.; Liu, C.Y.; Wu, M.C.; Shi, W.L. Kinetic study on the tyrosinase and melanin formation inhibitory activities of carthamus yellow isolated from Carthamus tinctorius L. J. Biosci. Bioeng 2013, 115, 242–245. [Google Scholar]
- Jones, K.; Hughes, J.; Hong, M.; Jia, Q.; Orndorff, S. Modulation of melanogenesis by aloesin: A competitive inhibitor of tyrosinase. Pigment Cell Res 2002, 15, 335–340. [Google Scholar]
- Kim, D.S.; Kim, S.Y.; Park, S.H.; Choi, Y.G.; Kwon, S.B.; Kim, M.K.; Na, J.I.; Youn, S.W.; Park, K.C. Inhibitory effects of 4-N-butylresorcinol on tyrosinase activity and melanin synthesis. Biol. Pharm. Bull 2005, 28, 2216–2219. [Google Scholar]
- Lin, C.C.; Yang, C.H.; Chang, N.F.; Wu, P.S.; Chen, Y.S.; Lee, S.M.; Chen, C.W. Study on the stability of DeoxyArbutin in an anhydrous emulsion system. Int. J. Mol. Sci 2011, 12, 5946–5954. [Google Scholar]
- Chang, T.-S. Natural melanogenesis inhibitors acting through the down-regulation of tyrosinase activity. Materials 2012, 5, 1661–1685. [Google Scholar]
- Chou, T.H.; Ding, H.Y.; Hung, W.J.; Liang, C.H. Antioxidative characteristics and inhibition of alpha-melanocyte-stimulating hormone-stimulated melanogenesis of vanillin and vanillic acid from Origanum vulgare. Exp. Dermatol 2010, 19, 742–750. [Google Scholar]
- Sato, K.; Takahashi, H.; Iraha, R.; Toriyama, M. Down-regulation of tyrosinase expression by acetylsalicylic acid in murine B16 melanoma. Biol. Pharm. Bull 2008, 31, 33–37. [Google Scholar]
- Beretta, G.L.; Gatti, L.; Perego, P.; Zaffaroni, N. Camptothecin resistance in cancer: Insights into the molecular mechanisms of a DNA-damaging drug. Curr. Med. Chem 2013, 20, 1541–1565. [Google Scholar]
- Bimonte, S.; Barbieri, A.; Palma, G.; Luciano, A.; Rea, D.; Arra, C. Curcumin inhibits tumor growth and angiogenesis in an orthotopic mouse model of human pancreatic cancer. BioMed Res. Int 2013, 2013, 8. [Google Scholar]
- Ozaki, T.; Ishiguro, S.; Itoh, H.; Furuhama, K.; Nakazawa, M.; Yamashita, T. Cisplatin binding and inactivation of mitochondrial glutamate oxaloacetate transaminase in cisplatin-induced rat nephrotoxicity. Biosci. Biotechnol. Biochem 2013, 77, 1645–1649. [Google Scholar]
- Lee, H.J.; Lee, H.J.; Lim, E.S.; Ahn, K.S.; Shim, B.S.; Kim, H.M.; Gong, S.J.; Kim, D.K.; Kim, S.H. Cambodian Phellinus linteus inhibits experimental metastasis of melanoma cells in mice via regulation of urokinase type plasminogen activator. Biol. Pharm. Bull 2005, 28, 27–31. [Google Scholar]
- Park, H.H. Structural features of caspase-activating complexes. Int. J. Mol. Sci 2012, 13, 4807–4818. [Google Scholar]
- Fiandalo, M.V.; Kyprianou, N. Caspase control: Protagonists of cancer cell apoptosis. Exp. Oncol 2012, 34, 165–175. [Google Scholar]
- Lai, Y.K.; Hsu, J.T.; Chu, C.C.; Chang, T.Y.; Pan, K.L.; Lin, C.C. Enhanced recombinant protein production and differential expression of molecular chaperones in sf-caspase-1-repressed stable cells after baculovirus infection. BMC Biotechnol 2012, 12, 83. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248. [Google Scholar]









© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, Y.-S.; Lee, S.-M.; Lin, C.-C.; Liu, C.-Y. Hispolon Decreases Melanin Production and Induces Apoptosis in Melanoma Cells through the Downregulation of Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Expressions and the Activation of Caspase-3, -8 and -9. Int. J. Mol. Sci. 2014, 15, 1201-1215. https://doi.org/10.3390/ijms15011201
Chen Y-S, Lee S-M, Lin C-C, Liu C-Y. Hispolon Decreases Melanin Production and Induces Apoptosis in Melanoma Cells through the Downregulation of Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Expressions and the Activation of Caspase-3, -8 and -9. International Journal of Molecular Sciences. 2014; 15(1):1201-1215. https://doi.org/10.3390/ijms15011201
Chicago/Turabian StyleChen, Yi-Shyan, Shu-Mei Lee, Chih-Chien Lin, and Chia-Yi Liu. 2014. "Hispolon Decreases Melanin Production and Induces Apoptosis in Melanoma Cells through the Downregulation of Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Expressions and the Activation of Caspase-3, -8 and -9" International Journal of Molecular Sciences 15, no. 1: 1201-1215. https://doi.org/10.3390/ijms15011201
APA StyleChen, Y. -S., Lee, S. -M., Lin, C. -C., & Liu, C. -Y. (2014). Hispolon Decreases Melanin Production and Induces Apoptosis in Melanoma Cells through the Downregulation of Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Expressions and the Activation of Caspase-3, -8 and -9. International Journal of Molecular Sciences, 15(1), 1201-1215. https://doi.org/10.3390/ijms15011201