Circulating MicroRNAs as Biomarkers of Acute Stroke

Abstract
:1. Introduction
2. Results and Discussion
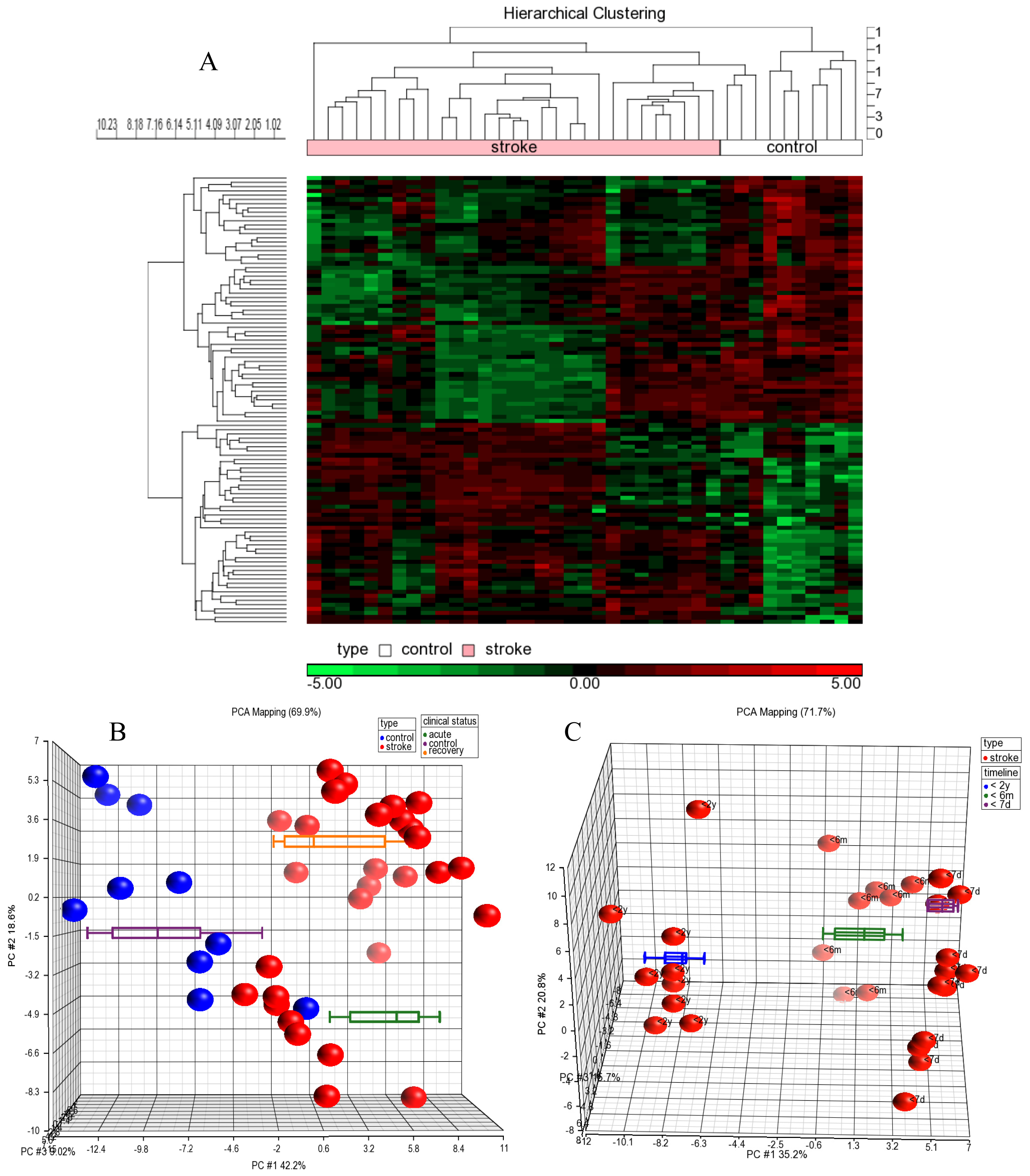
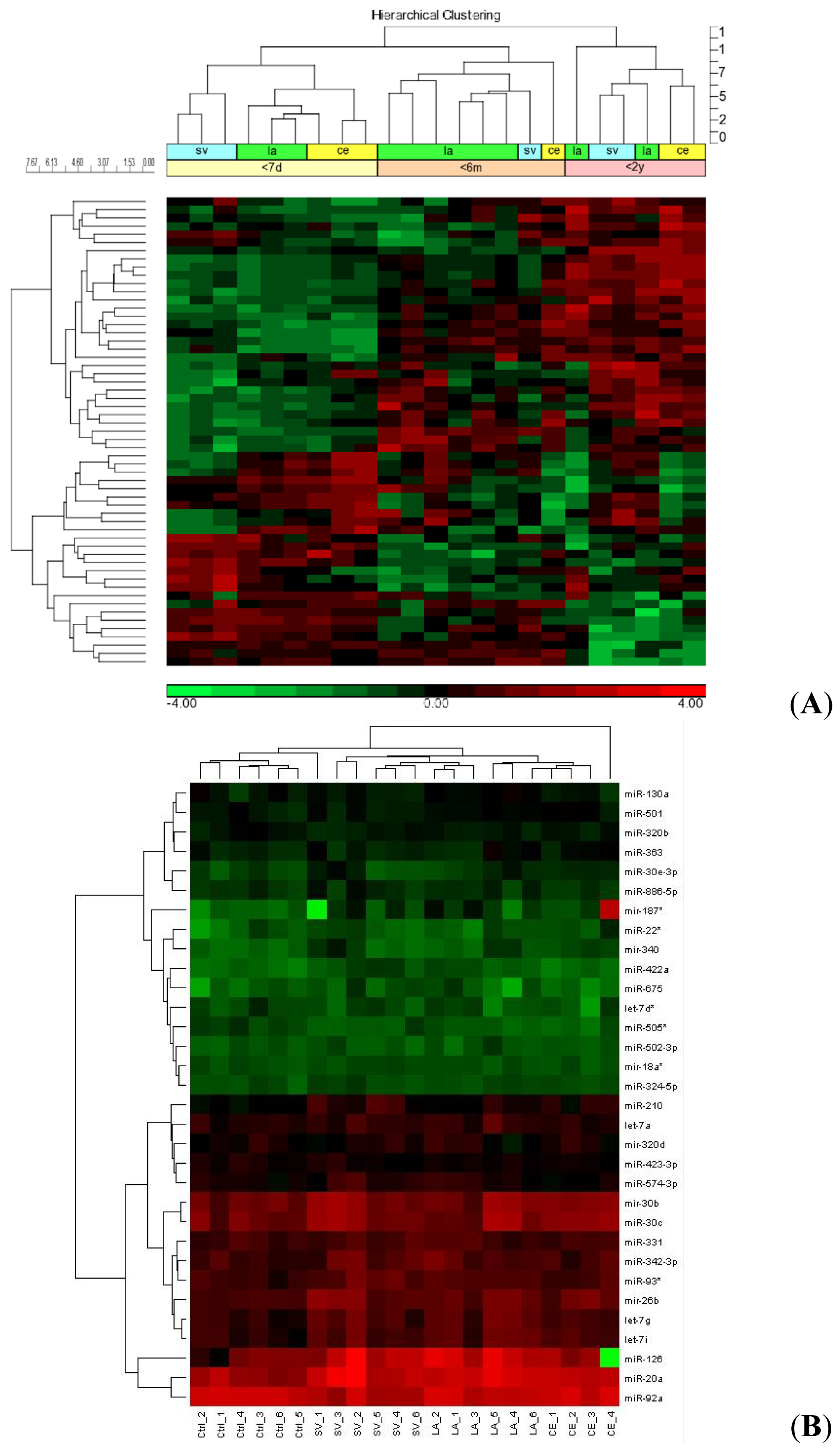
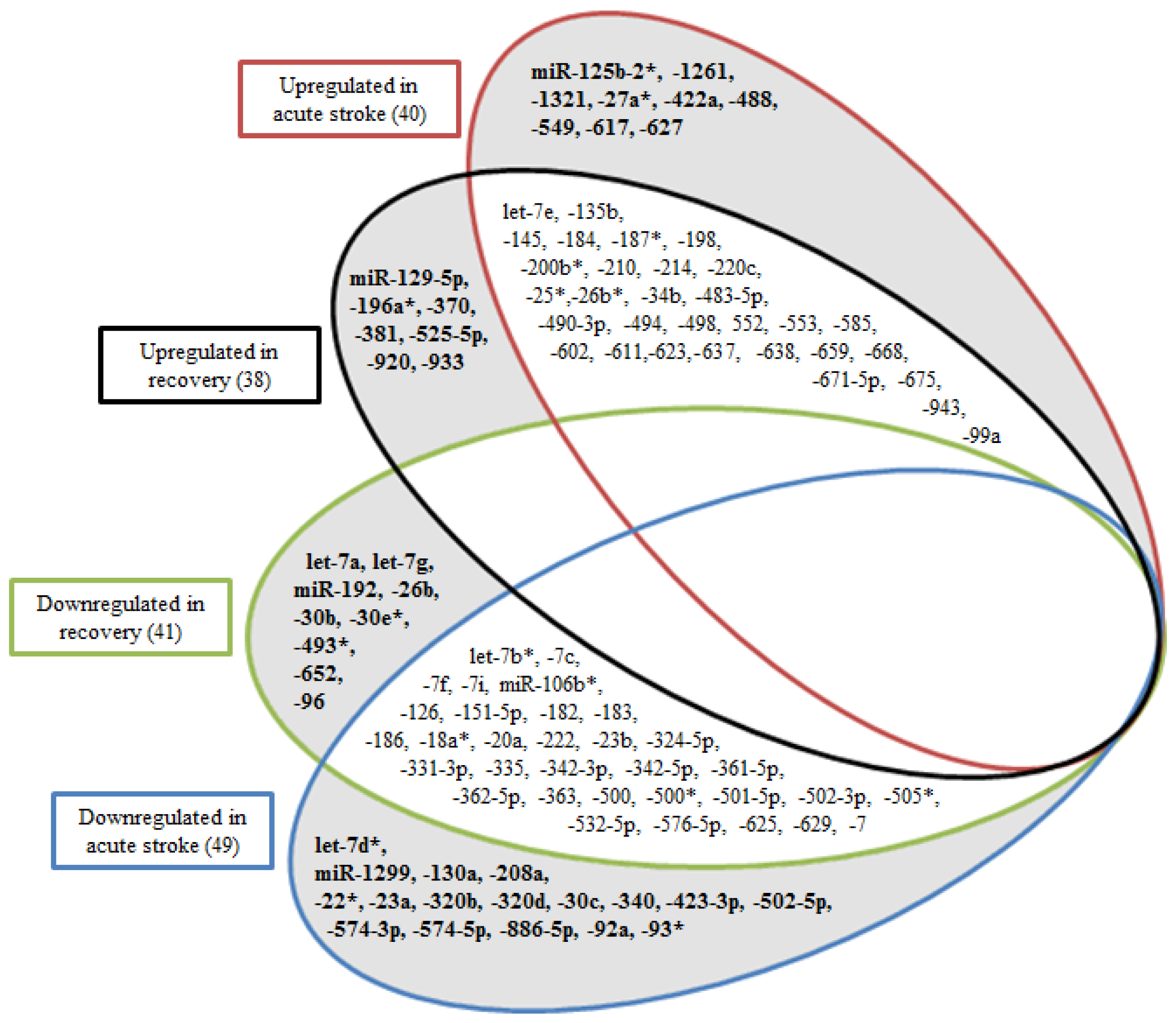
2.1. miRNA Profiles of Stroke Patients Reveal Temporal and Etiology Based Segregation
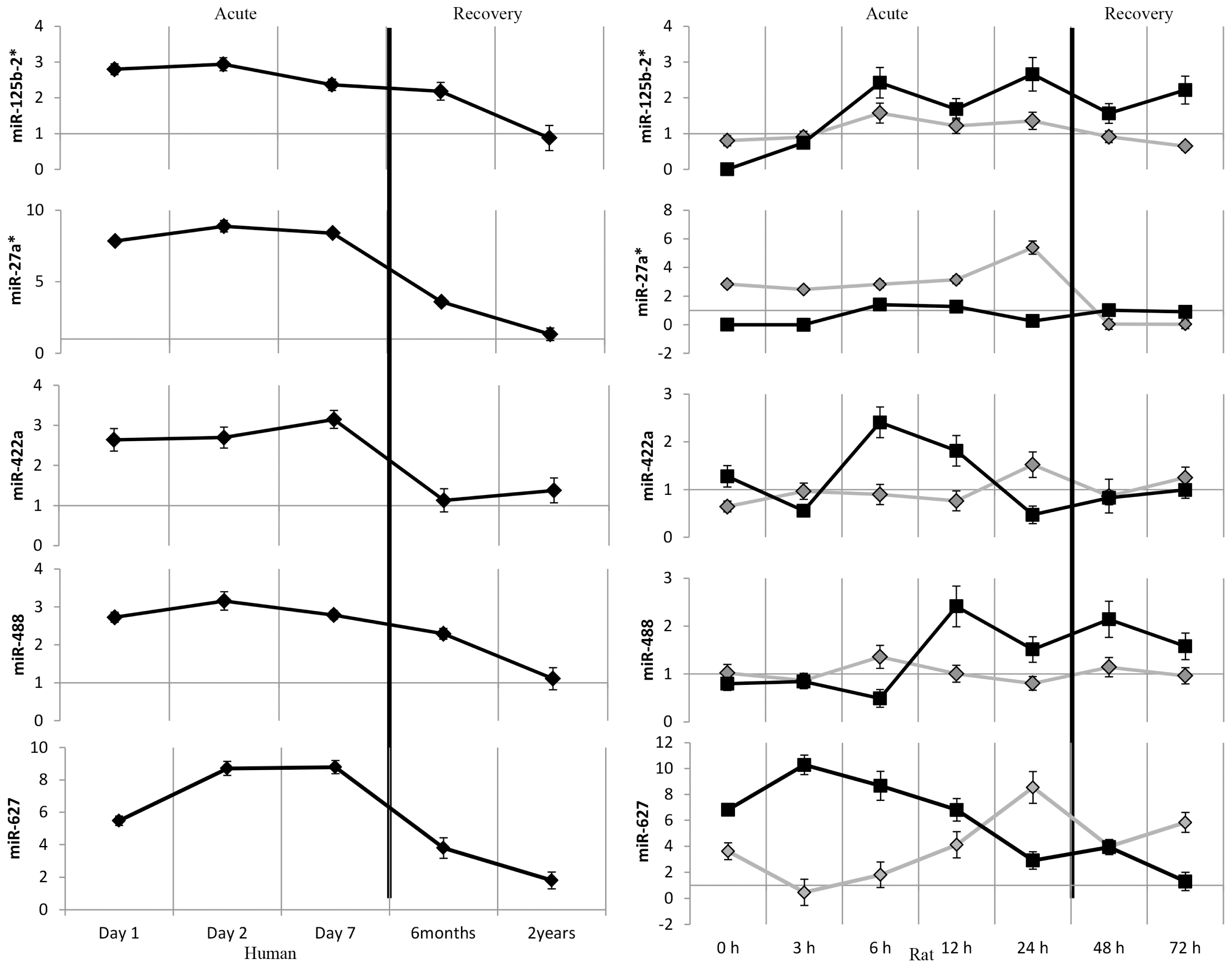
2.2. MicroRNAs as Biomarkers of Acute Stroke
3. Experimental Section
3.1. Patient Enrolment (Standard Protocol Approvals, Registrations, and Patient Consents)
3.2. Middle Cerebral Artery Occlusion (MCAo)
3.3. Total RNA Isolation
3.4. miRNA Microarray and Statistical Analysis
3.5. Quantitative PCR
4. Conclusions
Supplementary Information
ijms-15-01418-s001.pdfAcknowledgments
Conflicts of Interest
- Author ContributionsKJ, SS, DSK, KYL and AA perceived the concept, designed the experiments and wrote the manuscript. All experiments were carried out in KJ’s laboratory. AA performed the microarray. All other experiments including qPCR were carried out by SS, KYL, JRT, PK and FJL. DSK performed the statistical analysis and interpretation of data in relation to metabolic syndrome. JRT, KYL and FJL created the animal models for the study. Blood RNA extraction was carried out by FLY, JRT, DSK and AA. All bioinformatics analyses were performed by AA, SS, DSK and KYL under the guidance of KJ. Patient recruitment and collection of blood samples were carried out by KST, CWW, DAD, FPW and ST.
References
- Roger, V.L.; Go, A.S.; Lloyd-Jones, D.M.; Benjamin, E.J.; Berry, J.D.; Borden, W.B.; Bravata, D.M.; Dai, S.; Ford, E.S.; Fox, C.S.; et al. American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Heart disease and stroke statistics—2012 update: A report from the American Heart Association. Circulation 2012, 125, 188–197. [Google Scholar]
- Cucchiara, B.; Nyquist, P. Blood markers in TIA: Array of hope? Neurology 2011, 77, 1716–1717. [Google Scholar]
- Xu, J.; Zhao, J.; Evan, G.; Xiao, C.; Cheng, Y.; Xiao, J. Circulating microRNAs: Novel biomarkers for cardiovascular diseases. J. Mol. Med. Berl 2011, 90, 865–875. [Google Scholar]
- Jickling, G.C.; Sharp, F.R. Blood biomarkers of ischemic stroke. Neurotherapeutics 2011, 8, 349–360. [Google Scholar]
- Zhan, X.; Jickling, G.C.; Tian, Y.; Stamova, B.; Xu, H.; Ander, B.P.; Turner, R.J.; Mesias, M.; Verro, P.; Bushnell, C.; et al. Transient ischemic attacks characterized by RNA profiles in blood. Neurology 2011, 77, 1718–1724. [Google Scholar]
- Sharp, F.R.; Jickling, G.C.; Stamova, B.; Tian, Y.; Zhan, X.; Ander, B.P.; Cox, C.; Kuczynski, B.; Liu, D. RNA expression profiles from blood for the diagnosis of stroke and its causes. J. Child Neurol 2011, 26, 1131–1136. [Google Scholar]
- Jickling, G.C.; Xu, H.; Stamova, B.; Ander, B.P.; Zhan, X.; Tian, Y.; Liu, D.; Turner, R.J.; Mesias, M.; Verro, P.; et al. Signatures of cardioembolic and large-vessel ischemic stroke. Ann. Neurol 2010, 68, 681–692. [Google Scholar]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar]
- Wightman, B.; Ha, I.; Ruvkin, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. MicroRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Circ. Res 2010, 107, 810–817. [Google Scholar]
- Hauser, S.; Wulfken, L.M.; Holdenrieder, S.; Moritz, R.; Ohlmann, C.H.; Jung, V.; Becker, F.; Herrmann, E.; Walgenbach-Brünagel, G.; von Ruecker, A.; et al. Analysis of serum microRNAs (miR-26a-2*, miR-191, miR-337-3p and miR-378) as potential biomarkers in renal cell carcinoma. Cancer Epidemiol 2012, 36, 391–394. [Google Scholar]
- Gidlöf, O.; Andersson, P.; van der Pals, J.; Götberg, M.; Erlinge, D. Cardiospecific microRNA plasma levels correlate with troponin and cardiac function in patients with ST elevation myocardial infarction, are selectively dependent on renal elimination, and can be detected in urine samples. Cardiology 2011, 118, 217–226. [Google Scholar]
- Jeyaseelan, K.; Lim, K.Y.; Armugam, A. MicroRNA expression in the blood and brain of rats subjected to transient focal ischemia by middle cerebral artery occlusion. Stroke 2008, 39, 959–966. [Google Scholar]
- Tan, K.S.; Armugam, A.; Sepramaniam, S.; Lim, K.Y.; Setyowati, K.D.; Wang, C.W.; Jeyaseelan, K. Expression profile of microRNAs in young stroke patients. PLoS One 2009, 4, e7689. [Google Scholar]
- Dharap, A.; Bowen, K.; Place, R.; Li, L.C.; Vemuganti, R. Transient focal ischemia induces extensive temporal changes in rat cerebral microRNAome. J. Cereb. Blood Flow Metab 2009, 29, 675–687. [Google Scholar]
- Ziats, M.N.; Rennert, O.M. Identification of differentially expressed microRNAs across the developing human brain. Mol. Psychiatry 2013. [Google Scholar] [CrossRef]
- Zeng, L.; Liu, J.; Wang, Y.; Wang, L.; Weng, S.; Tang, Y.; Zheng, C.; Cheng, Q.; Chen, S.; Yang, G.Y. MicroRNA-210 as a novel blood biomarker in acute cerebral ischemia. Front. Biosci. Elite Ed 2011, 3, 1265–1272. [Google Scholar]
- Gan, C.S.; Wang, C.W.; Tan, K.S. Circulatory microRNA-145 expression is increased in cerebral ischemia. Genet. Mol. Res 2012, 11, 147–152. [Google Scholar]
- Ho, A.S.; Huang, X.; Cao, H.; Christman-Skieller, C.; Bennewith, K.; Le, Q.T.; Koong, A.C. Circulating miR-210 as a novel hypoxia marker in pancreatic cancer. Transl. Oncol 2010, 3, 109–113. [Google Scholar]
- Zhao, A.; Li, G.; Péoc’h, M.; Genin, C.; Gigante, M. Serum miR-210 as a novel biomarker for molecular diagnosis of clear cell renal cell carcinoma. Exp. Mol. Pathol 2013, 94, 115–120. [Google Scholar]
- Endo, K.; Naito, Y.; Ji, X.; Nakanishi, M.; Noguchi, T.; Goto, Y.; Nonogi, H.; Ma, X.; Weng, H.; Hirokawa, G.; et al. MicroRNA 210 as a biomarker for congestive heart failure. Biol. Pharm. Bull 2013, 36, 48–54. [Google Scholar]
- Ballabio, E.; Armesto, M.; Breeze, C.E.; Manterola, L.; Arestin, M.; Tramonti, D.; Hatton, C.S.; Lawrie, C.H. Bortezomib action in multiple myeloma: MicroRNA-mediated synergy (and miR-27a/CDK5 driven sensitivity)? Blood Cancer J 2012, 24, e83. [Google Scholar]
- Mitsios, N.; Pennucci, R.; Krupinski, J.; Sanfeliu, C.; Gaffney, J.; Kumar, P.; Kumar, S.; Juan-Babot, O.; Slevin, M. Expression of cyclin-dependent kinase 5 mRNA and protein in the human brain following acute ischemic stroke. Brain Pathol 2007, 17, 11–23. [Google Scholar]
- Muiños-Gimeno, M.; Espinosa-Parrilla, Y.; Guidi, M.; Kagerbauer, B.; Sipilä, T.; Maron, E.; Pettai, K.; Kananen, L.; Navinés, R.; Martín-Santos, R.; et al. Human microRNAs miR-22, miR-138-2, miR-148a, and miR-488 are associated with panic disorder and regulate several anxiety candidate genes and related pathways. Biol. Psychiatry 2011, 69, 526–533. [Google Scholar]
- Hu, H.Y.; Guo, S.; Xi, J.; Yan, Z.; Fu, N.; Zhang, X.; Menzel, C.; Liang, H.; Yang, H.; Zhao, M.; et al. MicroRNA expression and regulation in human, chimpanzee, and macaque brains. PLoS Genet 2011, 7, e1002327. [Google Scholar]
- Kin, K.; Miyagawa, S.; Fukushima, S.; Shirakawa, Y.; Torikai, K.; Shimamura, K.; Daimon, T.; Kawahara, Y.; Kuratani, T.; Sawa, Y. Tissue-and plasma-specific microRNA signatures for atherosclerotic abdominal aortic aneurysm. J. Am. Heart. Assoc 2012, 1, e000745. [Google Scholar]
- Tanaka, M.; Oikawa, K.; Takanashi, M.; Kudo, M.; Ohyashiki, J.; Ohyashiki, K.; Kuroda, M. Down-regulation of miR-92 in human plasma is a novel marker for acute leukemia patients. PLoS One 2009, 4, e5532. [Google Scholar]
- Chen, S.; He, Y.; Ding, J.; Jiang, Y.; Jia, S.; Xia, W.; Zhao, J.; Lu, M.; Gu, Z.; Gao, Y. An insertion/deletion polymorphism in the 3’ untranslated region of beta-transducin repeat-containing protein (β-TrCP) is associated with susceptibility for hepatocellular carcinoma in Chinese. Biochem. Biophys. Res. Commun 2010, 391, 552–556. [Google Scholar]
- Zernecke, A.; Bidzhekov, K.; Noels, H.; Shagdarsuren, E.; Gan, L.; Denecke, B.; Hristov, M.; Köppel, T.; Jahantigh, M.N.; Lutgens, E.; et al. Delivery of microRNA-126 by apoptotic bodies induces CXCL12-dependent vascular protection. Sci. Signal 2009, 2, ra81. [Google Scholar]
- Zhang, R.L.; Chopp, M.; Zhang, Z.G.; Jiang, Q.; Ewing, J.R. A rat model of focal embolic cerebral ischemia. Brain Res 1997, 766, 83–92. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Significantly upregulated miRNAs in all stroke cases | ||||||||
|---|---|---|---|---|---|---|---|---|
| miRNA | p-value | Fold Change | miRNA | p-value | Fold Change | miRNA | p-value | Fold Change |
| hsa-let-7e | 0.000 | 2.228 | hsa-miR-26b* | 0.004 | 3.725 | hsa-miR-602 | 0.000 | 2.546 |
| hsa-miR-125b-2* | 0.007 | 1.795 | hsa-miR-27a* | 0.002 | 3.745 | hsa-miR-611 | 0.002 | 1.815 |
| hsa-miR-1261 | 0.003 | 2.145 | hsa-miR-34b | 0.003 | 1.534 | hsa-miR-617 | 0.011 | 1.513 |
| hsa-miR-129-5p | 0.001 | 1.797 | hsa-miR-370 | 0.002 | 6.215 | hsa-miR-623 | 0.005 | 2.465 |
| hsa-miR-1321 | 0.008 | 1.836 | hsa-miR-381 | 0.014 | 1.719 | hsa-miR-627 | 0.003 | 3.992 |
| hsa-miR-135b | 0.000 | 6.512 | hsa-miR-422a | 0.002 | 1.755 | hsa-miR-637 | 0.002 | 1.737 |
| hsa-miR-145 | 0.000 | 8.353 | hsa-miR-483-5p | 0.000 | 3.482 | hsa-miR-638 | 0.000 | 2.410 |
| hsa-miR-184 | 0.006 | 1.449 | hsa-miR-488 | 0.006 | 2.124 | hsa-miR-659 | 0.000 | 2.464 |
| hsa-miR-187* | 0.000 | 2.404 | hsa-miR-490-3p | 0.001 | 4.836 | hsa-miR-668 | 0.003 | 2.541 |
| hsa-miR-196a* | 0.016 | 1.530 | hsa-miR-494 | 0.000 | 2.916 | hsa-miR-671-5p | 0.000 | 2.907 |
| hsa-miR-198 | 0.001 | 2.069 | hsa-miR-498 | 0.000 | 2.479 | hsa-miR-675 | 0.000 | 4.519 |
| hsa-miR-200b* | 0.000 | 2.573 | hsa-miR-525-5p | 0.002 | 1.999 | hsa-miR-920 | 0.005 | 3.482 |
| hsa-miR-210 | 0.000 | 4.923 | hsa-miR-549 | 0.007 | 1.514 | hsa-miR-933 | 0.015 | 1.437 |
| hsa-miR-214 | 0.005 | 1.666 | hsa-miR-552 | 0.000 | 4.077 | hsa-miR-943 | 0.000 | 2.976 |
| hsa-miR-220c | 0.000 | 11.799 | hsa-miR-553 | 0.000 | 15.392 | hsa-miR-99a | 0.000 | 6.401 |
| hsa-miR-25* | 0.000 | 2.145 | hsa-miR-585 | 0.000 | 7.330 | |||
| Significantly downregulated miRNAs in all stroke cases | ||||||||
|---|---|---|---|---|---|---|---|---|
| miRNA | p-value | Fold Change | miRNA | p-value | Fold Change | miRNA | p-value | Fold Change |
| hsa-let-7a | 0.002 | −1.622 | hsa-miR-222 | 0.003 | −1.569 | hsa-miR-500 | 0.004 | −1.626 |
| hsa-let-7b* | 0.001 | −2.087 | hsa-miR-23a | 0.008 | −1.730 | hsa-miR-500* | 0.000 | −2.309 |
| hsa-let-7c | 0.005 | −1.663 | hsa-miR-23b | 0.003 | −1.380 | hsa-miR-501-5p | 0.000 | −2.450 |
| hsa-let-7d* | 0.002 | −3.050 | hsa-miR-26b | 0.003 | −1.563 | hsa-miR-502-5p | 0.001 | −2.112 |
| hsa-let-7f | 0.001 | −1.790 | hsa-miR-30b | 0.003 | −1.503 | hsa-miR-502-3p | 0.000 | −2.264 |
| hsa-let-7g | 0.007 | −1.578 | hsa-miR-30c | 0.011 | −1.442 | hsa-miR-505* | 0.000 | −2.356 |
| hsa-let-7i | 0.005 | −1.703 | hsa-miR-30e* | 0.001 | −2.534 | hsa-miR-532-5p | 0.000 | −1.775 |
| hsa-miR-106b* | 0.009 | −1.332 | hsa-miR-320b | 0.016 | −2.085 | hsa-miR-574-5p | 0.005 | −1.569 |
| hsa-miR-126 | 0.000 | −2.041 | hsa-miR-320d | 0.016 | −1.766 | hsa-miR-574-3p | 0.014 | −1.528 |
| hsa-miR-1299 | 0.008 | −1.740 | hsa-miR-324-5p | 0.000 | −2.499 | hsa-miR-576-5p | 0.000 | −2.657 |
| hsa-miR-130a | 0.006 | −1.486 | hsa-miR-331-3p | 0.000 | −1.523 | hsa-miR-625 | 0.000 | −2.289 |
| hsa-miR-151-5p | 0.001 | −1.883 | hsa-miR-335 | 0.002 | −2.142 | hsa-miR-629 | 0.000 | −1.908 |
| hsa-miR-18a* | 0.000 | −1.647 | hsa-miR-340 | 0.007 | −1.498 | hsa-miR-652 | 0.001 | −1.398 |
| hsa-miR-182 | 0.000 | −2.533 | hsa-miR-342-3p | 0.000 | −2.066 | hsa-miR-7 | 0.000 | −2.229 |
| hsa-miR-183 | 0.000 | −2.137 | hsa-miR-342-5p | 0.000 | −2.135 | hsa-miR-886-5p | 0.006 | −2.427 |
| hsa-miR-186 | 0.002 | −1.793 | hsa-miR-361-5p | 0.000 | −2.022 | hsa-miR-92a | 0.003 | −1.420 |
| hsa-miR-192 | 0.002 | −2.183 | hsa-miR-362-5p | 0.002 | −1.944 | hsa-miR-93* | 0.002 | −1.632 |
| hsa-miR-20a | 0.003 | −1.633 | hsa-miR-363 | 0.000 | −1.769 | hsa-miR-96 | 0.005 | −2.101 |
| hsa-miR-208a | 0.005 | −2.085 | hsa-miR-423-3p | 0.016 | −1.614 | |||
| hsa-miR-22* | 0.011 | −1.441 | hsa-miR-493* | 0.011 | −2.058 | |||
| miRNAs | Cohort 1 | Cohort 2 | Cohort 3 | |||
|---|---|---|---|---|---|---|
| Stroke patients | Stroke patients | Metabolic syndrome patients | ||||
| AUC | 95% CI | AUC | 95% CI | AUC | 95% CI | |
| miR-125-2* | 0.95 ± 0.04 | 0.89–1.02 | 0.85 ± 0.05 | 0.77–0.93 | 0.67 ± 0.19 | 0.36–0.98 |
| miR-27a* | 0.89 ± 0.07 | 0.77–1.01 | 0.88 ± 0.05 | 0.81–0.96 | 0.67 ± 0.16 | 0.40–0.93 |
| miR-422a | 0.92 ± 0.06 | 0.82–1.02 | 0.86 ± 0.07 | 0.75–0.97 | 0.30 ± 0.17 | 0.02–0.58 |
| miR-488 | 0.87 ± 0.08 | 0.75–1.00 | 0.86 ± 0.06 | 0.72–0.92 | 0.56 ± 0.21 | 0.20–0.91 |
| miR-627 | 0.84 ± 0.08 | 0.70–0.98 | 0.76 ± 0.06 | 0.66–0.87 | 0.41 ± 0.25 | 0.01–0.82 |
| miR-920 | 0.81 ± 0.05 | 0.68–0.94 | 1.00 ± 0.00 | 1.00–1.00 | 0.33 ± 0.16 | 0.07–0.60 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sepramaniam, S.; Tan, J.-R.; Tan, K.-S.; DeSilva, D.A.; Tavintharan, S.; Woon, F.-P.; Wang, C.-W.; Yong, F.-L.; Karolina, D.-S.; Kaur, P.; et al. Circulating MicroRNAs as Biomarkers of Acute Stroke. Int. J. Mol. Sci. 2014, 15, 1418-1432. https://doi.org/10.3390/ijms15011418
Sepramaniam S, Tan J-R, Tan K-S, DeSilva DA, Tavintharan S, Woon F-P, Wang C-W, Yong F-L, Karolina D-S, Kaur P, et al. Circulating MicroRNAs as Biomarkers of Acute Stroke. International Journal of Molecular Sciences. 2014; 15(1):1418-1432. https://doi.org/10.3390/ijms15011418
Chicago/Turabian StyleSepramaniam, Sugunavathi, Jun-Rong Tan, Kay-Sin Tan, Deidre Ann DeSilva, Subramaniam Tavintharan, Fung-Peng Woon, Chee-Woon Wang, Fung-Lin Yong, Dwi-Setyowati Karolina, Prameet Kaur, and et al. 2014. "Circulating MicroRNAs as Biomarkers of Acute Stroke" International Journal of Molecular Sciences 15, no. 1: 1418-1432. https://doi.org/10.3390/ijms15011418
APA StyleSepramaniam, S., Tan, J. -R., Tan, K. -S., DeSilva, D. A., Tavintharan, S., Woon, F. -P., Wang, C. -W., Yong, F. -L., Karolina, D. -S., Kaur, P., Liu, F. -J., Lim, K. -Y., Armugam, A., & Jeyaseelan, K. (2014). Circulating MicroRNAs as Biomarkers of Acute Stroke. International Journal of Molecular Sciences, 15(1), 1418-1432. https://doi.org/10.3390/ijms15011418